
Paragymnomerus signaticollis tauricus ( Kostylev, 1940 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4378.3.10 |
|
publication LSID |
lsid:zoobank.org:pub:B7F0A2A3-01C4-4B93-86CB-1B97FCBF2F6F |
|
DOI |
https://doi.org/10.5281/zenodo.5998038 |
|
persistent identifier |
https://treatment.plazi.org/id/A11DD26C-FF9E-BF20-2697-A86902EC4D9D |
|
treatment provided by |
Plazi |
|
scientific name |
Paragymnomerus signaticollis tauricus ( Kostylev, 1940 ) |
| status |
|
Paragymnomerus signaticollis tauricus ( Kostylev, 1940) View in CoL
Odynerus signaticollis tauricus Kostylev 1940: 39 –40, ♀ (type locality: “ Crimée, Karadag ” [ Russia]), holotype , ♀, “ Tauria , Karadag ” ( ZMMU); van der Vecht & Fischer 1972: 17.
Paragymnomerus signaticollis tauricus: Kurzenko 1977: 681 View in CoL , Southern Crimea; Amolin 2009: 25–26, Ukraine (Luhansk Prov.); Fateryga & Ivanov 2009: 67, Crimea (Karadag); Fateryga 2010: 77, Crimea; 2018: 218, Crimea (several localities); Fateryga & Zhidkov 2012: 82, Crimea (Tarkhankut); Gusenleitner 2013a: 66, Turkey, Ukraine.
Paragymnomerus signaticollis: Kurzenko 1981: 95 View in CoL (part.), South Coast of the Crimea.
Both females and males of Paragymnomerus signaticollis tauricus View in CoL differ from that of the nominotypical subspecies, P. signaticollis signaticollis ( Morawitz, 1888) View in CoL , in having paler (whitish yellow not lemon yellow) and less extensive pale pattern of the body. Males of P. signaticollis tauricus View in CoL were not previously described, but all their structures, including the genitalia, do not differ from those of the nominotypical subspecies. Detailed description of the coloration of P. signaticollis tauricus View in CoL male is provided by Fateryga (2018).
Material examined. Holotype, ♀: Crimea: Karadag, 13.VII.1928, leg. G. Kostylev. Paratypes : Crimea: Karadag , 29.VI.1928, 4.VII.1928, 2 ♀, leg. G. Kostylev ( ZMMU). Additional material: Russia: Crimea: Tarkhankut Peninsula, Kipchak, 20.VI.2008, 1 ♀, leg. A. Fateryga , 17.VI.2015, 1 ♂, leg. V. Zhidkov; Tarkhankut Peninsula, Kipchak–Bolshoy Kastel, 16.VI.2015, 1 ♂, leg. V. Zhidkov; Tarkhankut Peninsula, Bolshoy Kastel, 4.VII.2014, 11.VI.2015, 2 ♀, leg. V. Zhidkov, 12.VII.2014, 13.VII.2014, 14.VII.2014, 22.VII.2015, 4.VI.2016, 29.VI.2016, 1–12.VI.2017, 22.VI.2017, 5 ♀, 4 ♂, leg. A. Fateryga (CFUS), 12–15. VII.2014, 4 ♀, leg. D. Puzanov (DPYE); Tarkhankut Peninsula, Dzhangul, 29.VI.2017, 5 ♂, leg. A. Fateryga; Sevastopol, Cape Lukull , 7.VII.2015, 2 ♀, leg. A. Fateryga ( CFUS).
Distribution. Russia (Crimea), Ukraine, Turkey. The nominotypical subspecies, Paragymnomerus signaticollis signaticollis , is distributed in Azerbaijan, Turkmenistan, Tajikistan, and Uzbekistan ( Kurzenko 1977). The species was omitted from the “Fauna Europaea” database ( Gusenleitner 2013b), though P. signaticollis tauricus was described from Europe (the Crimea).
Natural history. Nesting substrate. All studied nests of Paragymnomerus signaticollis tauricus were recorded on a slightly inclined (nearly horizontal) surface. The nesting site observed in 2015 (with 22 discovered nests) was an old dirt road overgrown with steppe vegetation dominated by Bothriochloa ischaemum (L.) Keng and Festuca valesiaca Schleich. ex Gaudin (Poaceae) and total coverage about 50% ( Fig. 1 View FIGURES 1–6 ). It was located in the valley “Bolshoy Kastel”. No nests were recorded there in 2016 and 2017, when the grass vegetation became denser. New nesting sites, however, were recorded in 2017. The first one, with 17 discovered nests, was located near the nesting site studied in 2015, but on the functioning dirt road. The second nesting site observed in 2017 (with 46 nests) was discovered in the natural boundary “Dzhangul” on a similar functioning dirt road ( Fig. 2 View FIGURES 1–6 ). The nests were located in each nesting site within an area of about 20–50 m 2. The soil at the nesting sites was brown carbonate clay loam of different densities. The dense clay loam was formed at the nesting site in 2015, the middle one at the first nesting site in 2017, and the light clay loam was formed at the second nesting site in 2017 ( Table 1). The greater content of physical clay parts at the nesting site in 2015 can be explained by mixing of clay with dust at two other sites on the functioning dirt roads.
Nesting activity. The females of Paragymnomerus signaticollis tauricus became active at 7.50 in case of sleeping outside of the nests or at 8.25 in case of sleeping in the nests and finished activity usually at 18.10 (solar time). Females started their nesting by searching for a place for the nest, which was a long process. In one case an observed female had finished her nest at 11.00 and began a new nest only on the next day at 11.20. After the place for the nest had been chosen, females went for water. The water collecting site ( Fig. 3 View FIGURES 1–6 ) was located 110 m directly west of the nesting site observed in 2015. There was a well at the water collecting site, which was probably made by Ancient Greeks and then modified in the modern time. This well was used by people resting there for taking water with a bucket and water was usually flowing out from it into a watering trough. Female wasps usually took water from this watering trough ( Fig. 4 View FIGURES 1–6 ). When there were no people there the females were observed taking water from the sea 30 m north of the main water collecting site. A flight for water usually lasted for less than a minute if females had not been disturbed by humans during the process. Otherwise they could be absent for several minutes.
A female returned to the nesting site with the water in her crop and regurgitated it onto the ground surface to moisten it. After that, she began to excavate a vertical nest burrow and construct the turret at the same time. She mixed mortar with water and soil, formed mud pellets, retrieved them from the nest burrow, and laid them around its entrance. After each mud pellet was laid, she smoothed its inner surface with her mouthparts. She held on to the inner surface of the turret with her fore legs and the outer one with the middle and hind legs ( Fig. 5 View FIGURES 1–6 ). In this way, the turret grew upwards and the burrow grew downwards at the same time. The turrets of P. signaticollis tauricus were not solid but had a laced structure with numerous minute perforations ( Fig. 6 View FIGURES 1–6 ).
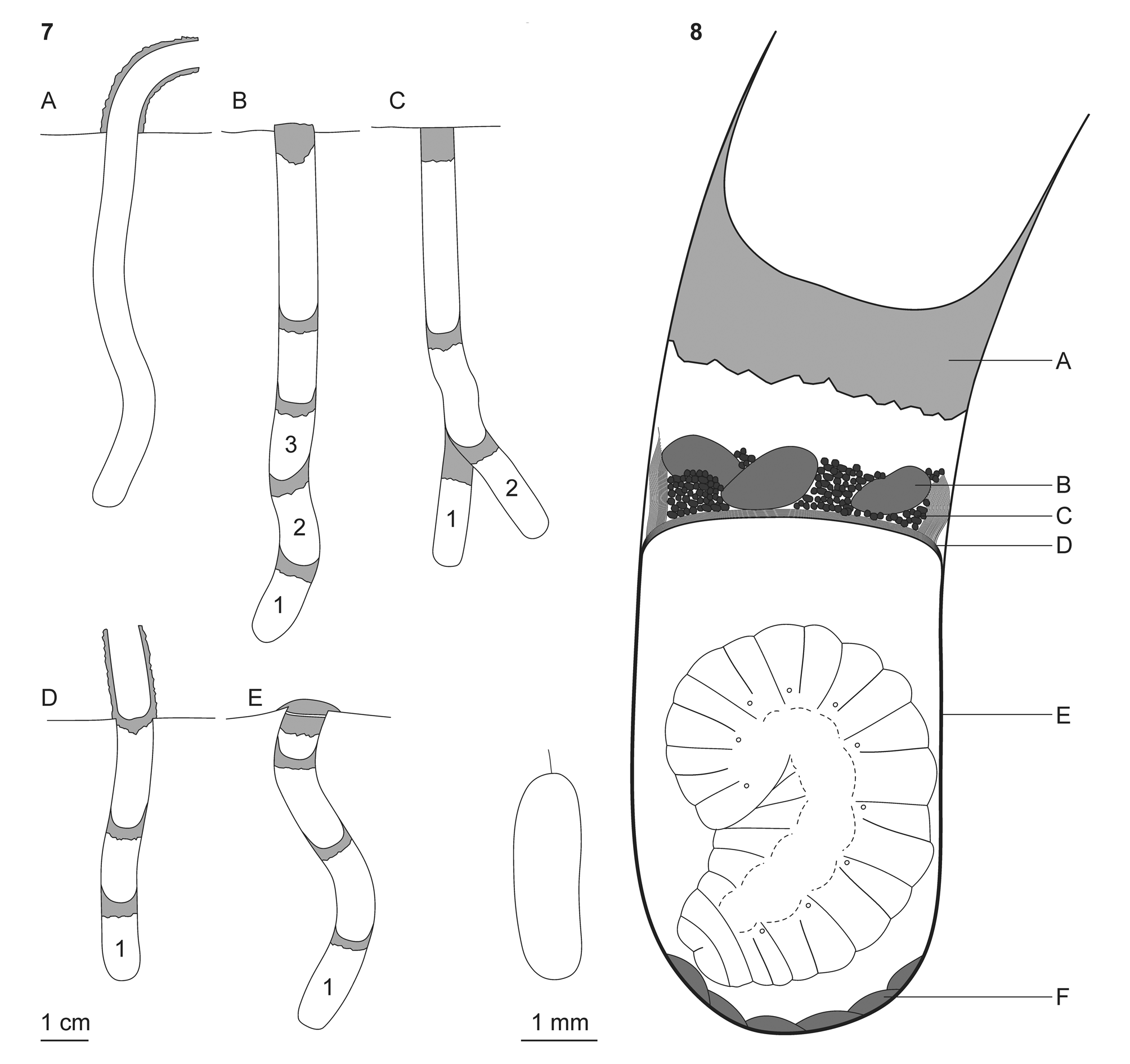
The complete turrets of the nests of P. signaticollis tauricus were 11–28 mm high and had a more or less curved shape ( Table 2) with the entrance opening sideways ( Fig. 7A View FIGURES 7–8 ). The construction of the turret lasted for 97 min. in one observed case (the constructed turret was 19 mm high and its deflection from the vertical position was 7 mm; the female wasp flew for water eight times during the building process).
After the turret was completed, the female had been continuing the excavation of the nest burrow for several hours (depending on its depth). At this time she also retrieved mud pellets from the nest, but these pellets were noticeably bigger than those used for the turret. The female retrieved them from the nest entrance and tried to fly up ( Fig. 9 View FIGURES 9–14 ). Usually she accomplished this only after several attempts and then dropped the pellet away from the nest entrance before returning to the nest. The excavation of the nest burrow was periodically interrupted by flights for water.
When the nest burrow was completed the female laid an egg and flew for hunting. After hunting and provisioning, the female sealed the cell with a mud partition. Then she did one of the following three actions: laid the second egg within the same nest burrow, excavated the second cell as its lateral branch, or sealed the whole nest. Only the third case was directly observed. The female flew for water and then regurgitated it onto the top of the turret. Then, she started forming a mud pellet with her mouthparts while holding on to the turret and/or the ground surface with all of her legs ( Fig. 10 View FIGURES 9–14 ). Then, the female took the pellet with her fore legs and came down into the nest where she used it to construct a transverse mud partition directly above the last nest cell. Her work was periodically interrupted by flights for water. In this way, she also built one or two additional partitions above the last cell. Only after that the female made the final nest plug at the ground level. This plug was much thicker ( Fig. 7B–C View FIGURES 7–8 ) than the cell partitions and the “vestibular” partitions; in one case it consisted of two parts ( Fig. 7E View FIGURES 7–8 ). The nest turret was generally completely demolished during the sealing of the nest ( Fig. 11 View FIGURES 9–14 ) and the final plug usually became indistinguishable from the surrounding ground after drying. One studied nest had a partially undemolished turret ( Fig. 7D View FIGURES 7–8 ) that had been probably caused by the death of the female wasp. Generally females used only the material of the demolished turret to seal the nest. One of the observed nests, however, had a partially broken turret; the female demolished it and then took two additional mud portions from the ground surface near the nest. The sealing of the nest took 60 and 80 min. in two observed cases.
Hunting. The hunting site ( Fig. 12 View FIGURES 9–14 ) was located 50 m directly south of the nesting site studied in 2015. The prey of Paragymnomerus signaticollis tauricus were sawfly larvae of the genus Megalodontes Latreille ( Hymenoptera : Megalodontesidae ). At least some of them belonged to M. plagiocephalus (Fabricius) forma “a” sensu Taeger (2002). They were identified as described in the “Material and methods” section. Adult sawflies of another species, M. medius Konow , however, were found at the same place in 2017. Thus, the larvae used by P.
signaticollis tauricus as the prey might belong also to this species. All observed larvae were feeding on a single host plant species, Seseli tortuosum L. ( Apiaceae ). Each larva inhabited a personal silk web ( Fig. 13 View FIGURES 9–14 ).
Hunting was observed 17 times. Each time females made patrolling flights around the plants of S. tortuosum . Once a female noticed a web with Megalodontes larva, it landed and went deep into its proximal end ( Fig. 14 View FIGURES 9–14 ). This action made the larva come closer to the distal end of the web and then female came there and tried to take it. The larva retreated to the proximal end of the web. These actions repeated several times: the female went from proximal to distal end of the web and the larva retreated from her in the opposite direction. Three times females left their attempts to take larvae and started searching other webs. After 6–8 attempts, however, a female usually successfully took the larva from the distal end of the web. She grasped it by the distal end of the body with her mandibles and fore legs. Then, the female took also the proximal end of the larva with her middle legs and stung it several times in the thoracic segments ( Fig. 15 View FIGURES 15–20 ). The female’s hind legs were used during these actions to hold on to the web. Next, the female turned the larva and seized its prothorax with mandibles. After that she flew away. The whole hunting usually took 0.5–1.5 min. and the stinging lasted for 6–8 sec.
Feeding of adult females on Megalodontes larva was observed two times. Each time the female took the larva in the way described above, but did not fly away with it. Instead of that she flew onto a nearest grass stem (in the observed cases it was Marrubium peregrinum L.). Then, she gnawed the prothorax of the victim and had been sucking the hemolymph from it during 1–2 min. ( Fig. 16 View FIGURES 15–20 ). Later, she dropped the larva and flew away. Examination of the dropped larvae revealed small wounds on either the ventral or lateral surfaces of the prothorax.
Each female observed in 2015 brought 1– 2 larvae per day to her nest. This low rate of provisioning was probably connected with the possible end of the nesting period of P. signaticollis tauricus observed on July 22–26, 2015. The provisioning observed on June 29–July 1, 2017 was more rapid (up to five larvae per day). There were 3 to 5 sawfly larvae stored in one cell ( Table 2; Fig. 17 View FIGURES 15–20 ).
Structure of complete nests. The main parameters of the nests of Paragymnomerus signaticollis tauricus are summarized in Table 2. The complete nests contained 1–4 cells; the nest burrow was 55–120 mm deep and 7–9 mm wide. The nests with several cells had either linear ( Fig. 7B View FIGURES 7–8 ) or branched structure ( Fig. 7C View FIGURES 7–8 ). The nest burrow was vertical, somewhat curved (more or less S-shaped). The inner walls of the nest burrow were smooth, but minutely transversely wrinkled. The nest burrow was not filled with mud because only turret material had been usually used by females to seal the nest (see above). The cells of P. signaticollis tauricus and other parts of the nest burrow were of the same diameter; the inner length of each cell varied from 13 to 23 mm ( Table 2). Each cell in the nests was sealed by a mud partition; similar partitions were found in “vestibular” space. The nest plug was thicker than the other partitions and usually towered 1–2 mm above the ground surface.
Immature instars and development. Five nests dissected during the provisioning period and directly after they had been sealed by females contained eggs in their single cells. Each egg lay loose on the bottom of the cell. Nevertheless, the eggs had distinct filaments ( Fig. 8 View FIGURES 7–8 ). The eggs had probably been initially suspended by the females on the cell walls, but then were torn off and fell. The egg size was 2.3–2.8×0.8–0.9 mm (mean— 2.66±0.09×0.88±0.02 mm; N =5); the length of the filament was 0.2–0.3 mm.
When the larva hatched from the egg, it immediately began feeding on the stored sawfly larvae. After consuming all provisions, it spun a cocoon. The cocooning began with storing prey remains and their feces at the top of the cell. These waste products were separated from the remaining part of the cell with multilayered part of the cocoon, with thin layers. Then, the larva constructed the thick part of the cocoon that covered the bottom of the cell and its lateral walls. This part was made with the help of fluid secretion penetrating into the cell walls and making them durable and water-proof. After the cocoon construction, the meconium was discharged onto the bottom of the cell. Then, the larva passed to the prepupal stage and coiled its body. The inner volume of the cocoon exceeded the size of the prepupa ( Fig. 8 View FIGURES 7–8 ). Hibernation occurred at the prepupal stage ( Fig. 18 View FIGURES 15–20 ). After the winter diapause the prepupa unrolled and straightened its body. Pupation occurred approximately 10 days after unrolling; the pupa developed for 20–25 days. Four prepupae from two nests were obtained in 2015. One male from the first nest emerged on June 4, 2016, and two males from the second nest emerged in June, 2017. The third prepupa from the second nest was stored in ethanol and its development was not observed. These observations showed that Paragymnomerus signaticollis tauricus is an univoltine species. Wasps of this species were recorded from the beginning of June till the end of July (see “Material examined”). The period of their activity, however, is somewhat shorter (for about a month) in a separate year.
Parasites, associates, and reproductive success. Cuckoo wasps ( Hymenoptera : Chrysididae ) were found in two of the 15 observed complete cells. They were, with high probability, Chrysis valesiana Frey-Gessner. Females of this species were observed and caught near the nests; this chrysidid species is known as the parasite of Paragymnomerus spiricornis and two other ground nesting eumenines of the closely related genera Paravespa and Tropidodynerus ( Martynova & Fateryga 2015) . The first cell with the cuckoo wasp was found in the largest nest with four cells which was dissected after its sealing by the female of Paragymnomerus signaticollis tauricus . This cell contained a young chrysidid larva. It consumed three of the five Megalodontes larvae stored there and then spun a cocoon. Thus, this cuckoo wasp acted as a cleptoparasite. Unfortunately, it died inside its cocoon. The second cell with the cuckoo wasp was the second cell in the branched two-celled nest ( Fig. 7C View FIGURES 7–8 ); this nest was dissected in 2017, a month after they had been sealed by the female wasp. The cell contained the cocoon of the cuckoo wasp along with two Megalodontes larvae which were not consumed by the cleptoparasite. The chrysidid prepupa in the cocoon had not pupated before the present paper was submitted. Cocoons of the observed cuckoo wasp were remarkable. They consisted of two layers and were not different from the cocoons of Ch. valesiana observed in the nests of Paravespa rex (von Schulthess) ( Fateryga & Ivanov 2013).
Other observed cells of P. signaticollis tauricus were free of parasites. However, all larvae from the nests dissected in July, 2015 and in June, 2017 (during the provisioning period and directly after they had been sealed by the females) died during their transportation to the laboratory.
Two ant species were observed disturbing the nesting females of P. signaticollis tauricus : Cataglyphis aenescens (Nylander) and rarely Messor structor (Latreille) ( Hymenoptera : Formicidae ). They attacked the females to steal portions of water which had been regurgitated by the wasps to make a mortar. This behavior of the ants was observed only at the beginning of the excavation of the nest burrow and at the ending of the sealing of the nest by the females. The ants, however, did not cause a significant inconvenience to the wasps—the females immediately drove them away and returned to their work.
Nearly 85% of the nests had been abandoned by females before the excavation of the nest burrow was finished. Most of them were left for no visible reason; one nest contained a dead female wasp which had also died for unknown reason. One nest was left because the female had been disturbed by humans. The burrows of four abandoned nests were occupied by nests of two bee species: Hoplitis papaveris (Latreille) (three nests) ( Fig. 19 View FIGURES 15–20 ) and Megachile pilidens Alfken (one nest) ( Hymenoptera : Megachilidae ).
One observed nest with the turret broken by humans was not abandoned by the female. Instead, she threw two stored sawfly larvae out of the nest burrow and then she started to deepen the nest burrow and to construct a new turret. The egg was probably also discarded in that case. Another observed nest with the turret accidentally broken during the observations was also not abandoned. This time the female, however, did not throw any contents of the cell. Instead, she took the mud for the new turret from lateral walls of the nest burrow and the latter became more curved ( Fig. 7E View FIGURES 7–8 ). Then, she demolished the newly constructed turret and sealed the nest.
Male behavior. Males were recorded at the hunting site in 2015–2017 and at the second nesting site in 2017. They made patrolling flights around the plants of Seseli tortuosum infested by the Megalodontes larvae at the hunting site and along the dirt road at the second nesting site in 2017. In the latter case they sometimes set on small stones near nests and waited for females; these males attacked females arriving to nests but the females generally rejected them immediately. A courtship was observed only once on a branch of Centaurea lavrenkoana Klokov in the steppe near the second nesting site in 2017 ( Fig. 20 View FIGURES 15–20 ). Copulation was not observed. Adult feeding of the males was recorded on flowers of Teucrium chamaedrys L. and T. capitatum L. ( Lamiaceae ). No males were recorded at the water collecting site.



| ZMMU |
Zoological Museum, Moscow Lomonosov State University |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Paragymnomerus signaticollis tauricus ( Kostylev, 1940 )
| Fateryga, Alexander V. 2018 |
Odynerus signaticollis tauricus
| Kostylev 1940 : 39 |
| Vecht & Fischer 1972 : 17 |
Paragymnomerus signaticollis tauricus: Kurzenko 1977 : 681
| Kurzenko 1977 : 681 |
| Fateryga & Ivanov 2009 : 67 |
| Fateryga 2010 : 77 |
| Fateryga & Zhidkov 2012 : 82 |
| Gusenleitner 2013a : 66 |
Paragymnomerus signaticollis:
| Kurzenko 1981 : 95 |