
Minibiotus diversus, Roszkowska, Daniel Adrian Ciobanu Milena & Kaczmarek, Łukasz, 2015
|
publication ID |
https://doi.org/10.11646/zootaxa.3941.4.4 |
|
publication LSID |
lsid:zoobank.org:pub:66179A2D-14A6-4C5F-91F7-331E7922D1B0 |
|
DOI |
https://doi.org/10.5281/zenodo.6106651 |
|
persistent identifier |
https://treatment.plazi.org/id/A12287B0-FFC0-FFB2-E6B3-FBD026BA0898 |
|
treatment provided by |
Plazi |
|
scientific name |
Minibiotus diversus |
| status |
sp. nov. |
Minibiotus diversus View in CoL sp. nov.
http://www.tardigrada.net/register/0019.htm ( Figs 15 – 31 View FIGURES 15 – 16 View FIGURES 17 – 18 View FIGURES 19 – 24 View FIGURES 25 – 29 View FIGURES 30 – 31 , Tables 3 – 4 View TABLE 3 View TABLE 4 )
Material examined: Holotype, 100 paratypes, three exuvia and five eggs.
Description of the adults ( Figs 15 – 16 View FIGURES 15 – 16 ) (measurements in µm, pt ratios and statistics in Table 3 View TABLE 3 ): Body white/yellowish (in live specimens) or transparent (after fixation in Hoyer’s medium). Eyes present. Entire cuticle covered by numerous round or oval pores (0.6 – 4.8 Μm in diameter). Pores are distributed randomly on dorsal side of the body or arranged in more or less regular rows (lines) on ventral side ( Figs 15 – 16 View FIGURES 15 – 16 and 19 – 20 View FIGURES 19 – 24 ). Large, circular dorsal pores include: 3 – 6 pores (2.2 – 3.2 Μm in diameter) on head region ( Fig. 22 View FIGURES 19 – 24 ) and 3 – 5 pores (2.2 – 3.2 Μm in diameter) caudally, just above IV pair of legs ( Figs 23 – 24 View FIGURES 19 – 24 ). Large pores (3.1 – 4.8 Μm in diameter) also present on external side of legs I–III (one on each leg) ( Fig. 29 View FIGURES 25 – 29 ). Pores on ventral side less densely distributed and arranged in loose bands of 4 – 5 larger pores with smaller pores between ( Fig. 19 View FIGURES 19 – 24 ). On ventral side of head a triangular shape is formed by two rows of pores (three pores in first row and two or single pore in second) ( Fig. 21 View FIGURES 19 – 24 ). Overall, pores are larger on caudal end, head segment and on external side of legs I – III. A ring of pores around the mouth opening absent. Cribriform areas not visible. A fine granulation on all legs present ( Figs 25 – 28 View FIGURES 25 – 29 ).
Mouth antero-ventral. Bucco-pharyngeal apparatus of the Minibiotus type ( Fig. 17 View FIGURES 17 – 18 ). Oral cavity armature absent or not visible under PCM. Buccal tube with poorly visible ventral lamina and with an anterior and a posterior bend (both visible in lateral view only). Buccal tube walls just below stylet attachments thickened ( Fig. 17 View FIGURES 17 – 18 ). Pharyngeal apophyses triangular or round; close to the first macroplacoid. Three granular macroplacoids and a minute microplacoid present in the round pharyngeal bulb. Septulum absent. All macroplacoids similar, almost identical in size and the macroplacoid length sequence: 1=2 ≅ 3 ( Fig. 17 View FIGURES 17 – 18 ).
Claws of the Macrobiotus type ( Figs 18 View FIGURES 17 – 18 and 25 – 28 View FIGURES 25 – 29 ). Primary branches of claws with thin accessory points. Smooth lunules present on all legs. Bars and other cuticular thickenings on legs absent.
Eggs (all measurements in Table 4 View TABLE 4 ): White, laid free, spherical and without areolation ( Figs 30 – 31 View FIGURES 30 – 31 ). Processes screw-like, each surrounded by a separate membrane ( Fig. 31 View FIGURES 30 – 31 ). On the egg circumference 21–22 processes present. Surface between processes smooth under PCM ( Fig. 30 View FIGURES 30 – 31 ).
Locus typicus: 47°41'19.6''N, 26°57'36.6''E; 147 m asl: Romania, Botoşani County, Suliţa village, Cozancea Monastery, deciduous forest ( Fagus sylvatica L., Quercus robur L. and Q. petraea L.), moss from tree.
Etymology: In Latin ‘ diversus’ means ‘diverse’ referring to the fact that this species has several different types of pores in the cuticle.
Type depositories: Holotype (slide: MC1-1), 54 paratypes (slides MC1-2, MC1-3, MC1-5, MC1-6, MC1-7, MC1-8, MC1-13) and three eggs (slide MC1-4, MC1-13) are preserved at the Department of Animal Taxonomy and Ecology, Adam Mickiewicz University in Poznań, Umultowska 89, 61–614 Poznań, Poland. Additionally, 17 paratypes (slides: MC1-11, MC1-14) and one egg (slide MC1-16) are deposited at Natural History Museum of “Alexandru Ioan Cuza” University from Iaşi, Romania (Bd. Independenţei No.16, 700101), 29 paratypes (slides: MC1-9, MC1-10, MC1-12, MC1-17) and one egg (slide MC1-15) are deposited at collection of Peter Degma, the Department of Zoology, Comenius University, Bratislava, Slovakia.
Differential diagnosis. The new species, in the arrangements of cuticle pores, are most similar to M. gumersindoi Guil & Guidetti, 2005 and in adults and eggs morphology to M. weglarskae Michalczyk et al., 2005 , but it differs from:
M. gumersindoi by: different arrangement of pores on the dorsal side of the body (randomly arranged pores in M. diversus sp. nov. vs pores arranged in an undefined number of transverse bands in M. gumersindoi ), the presence of triangular formation of pores on the ventral side of the head, the absence of triangle/ pentagonal-like formation of pores on legs I – III, presence of granulation on legs, presence of eyes and by slightly longer macroplacoids (mI/mII: 1.1–1.7 Μm; mIII: 1.2–1.7 Μm in M. diversus sp. nov. vs all 1.0 Μm in M. gumersindoi ).
M. weglarskae by: arrangement of pores on the dorsal side of the body (randomly arranged pores in M. diversus sp. nov. vs pores arranged in 10 transversal bands in M. weglarskae ), absence of bi-, trilobed and starshaped pores, the presence of triangular formation of pores on the ventral side of the head, the presence of large pores on the frontal and caudal end, the presence of large pores on the external side of legs I – III, stylet supports inserted in a more anterior position ([ 50.5–52.9] in M. diversus sp. nov. vs [ 54.5–59.6] in M. weglarskae ), a smaller pt of buccal tube internal width ([ 2.1–3.0] in M. diversus sp. nov. vs [ 4.0–4.7] in M. weglarskae ), slightly larger diameter of eggs without processes (44.9 – 47.8 µm in M. diversus sp. nov. vs ca. 40 µm in M. weglarskae ), slightly larger diameter of eggs with processes (51.5 – 53.9 µm in M. diversus sp. nov. vs ca. 50 µm in M. weglarskae ), slightly smaller number of processes on the egg circumference ( 21 – 22 in M. diversus sp. nov. vs ca. 24 in M. weglarskae ), shorter egg processes (3.1 – 3.6 µm in M. diversus sp. nov. v.s 5.0 – 5.5 µm in M. weglarskae ), smaller diameter of the distal disc of the processes (3.0 – 3.8 µm in M. diversus sp. nov. vs 5.0 – 5.5 µm in M. weglarskae ) and slightly larger distances between processes (3.6–4.8 µm in M. diversus sp. nov. vs ca. 3.0 µm in M. weglarskae ).
M. diversus View in CoL sp. nov. is also very similar in some aspects of the adult and/ or egg morphology to: M. bisoctus ( Horning et al. 1978) View in CoL , M. eichhorni Michalczyk & Kaczmarek, 2004 View in CoL , M. formosus Zawierucha et al. 2014 View in CoL , M. furcatus ( Ehrenberg, 1859) View in CoL , M. harrylewisi Meyer & Hinton, 2009 View in CoL , M. jonesorum Meyer et al., 2011 View in CoL , M. keppelensis Claxton, 1998 View in CoL , M. orthofasciatus Fontoura et al., 2009a View in CoL , M. poricinctus Claxton, 1998 View in CoL , M. pustulatus ( Ramazzotti, 1959) View in CoL , M. ramazzottii Binda & Pilato, 1992 View in CoL , M. vinciguerrae Binda & Pilato, 1992 View in CoL and M. xavieri Fontoura et al., 2009b View in CoL , but it differs from:
M. bisoctus View in CoL by: arrangement of pores on dorsal side of the body (randomly arranged pores in M. diversus View in CoL sp. nov. vs pores arranged in undefined number of transverse bands in M. bisoctus View in CoL ), absence of bi-, trilobed and starshaped pores, the presence of triangular formation of pores on the ventral side of the head, the presence of large pores on the frontal and caudal end, the presence of large pores on the external side of legs I – III, stylet supports inserted in a more anterior position ([ 50.5–52.9] in the new species vs ca. [ 60.3] in M. bisoctus View in CoL ) smaller pt of macroplacoid row ([ 22.3–25.8] in M. diversus View in CoL sp. nov. vs ca. [ 31.0] in M. bisoctus View in CoL (according to Claxton 1998)).
M. eichhorni View in CoL by: arrangement of pores on dorsal side of the body (randomly arranged pores in M. diversus View in CoL sp. nov. vs pores arranged in six transverse bands in M. eichhorni View in CoL ), absence of star-shaped pores and four pores around the mouth opening, the presence of triangular formation of pores on the ventral side of the head, the presence of large pores on the external side of legs I – III, a different macroplacoid sequence (1=2 ≅ 3 in the new species vs 2<3<1 Μm in M. eichhorni View in CoL ), slightly smaller body size (147 – 200 µm in M. diversus View in CoL sp. nov. vs 204.3–389.5 µm in M. eichhorni View in CoL ), shorter buccal tube (19.8 – 23.9 µm in M. diversus View in CoL sp. nov. vs 24.7 – 34.2 µm in M. eichhorni View in CoL ), stylet supports inserted in a more anterior position ([ 50.5–52.9] in M. diversus View in CoL sp. nov. vs [ 65.4–70.6] in M. eichhorni View in CoL ), smaller external width of buccal tube (1.3 – 1.7 µm in M. diversus View in CoL sp. nov. vs 1.9 – 3.3 µm in M. eichhorni View in CoL ), shorter placoids (mI/mII: 1.1–1.7 Μm [ 5.5–7.2]; mIII: 1.2–1.7 Μm [ 6.0–7.1]; mi: 0.4 – 0.7µm [ 2.0–3.0] in M. diversus View in CoL sp. nov. vs mI: 2.9–4.3µm [ 10.3–12.5]; mII: 1.9–3.1 µm [ 6.9–9.2]; mIII: 2.4–3.4 µm [ 9.3–10.6]; mi: 1.1–1.9 µm [ 4.6–6.1] in M. eichhorni View in CoL ) and shorter external claws I–IV (I: 4.6–6.0µm; II: 4.7–6.6 µm; III: 4.5–6.2µm; IV: 4.8–7.0 µm in M. diversus View in CoL sp. nov. vs I: 6.7 – 10.5 µm; II: 6.7 – 11.4 µm; III: 7.6 – 11.4 µm; IV: 8.6 – 14.3 µm in M. eichhorni View in CoL ).
M. formosus View in CoL by: arrangement of pores on dorsal side of the body (randomly arranged pores in M. diversus View in CoL sp. nov. vs pores arranged in 9 – 10 transverse bands in M. formosus View in CoL ), the presence of large pores on the frontal and caudal end, the presence of triangular formation of pores on ventral side of the head, the presence rows of pores between the legs, a different macroplacoid length sequence (1=2 ≅ 3 in M. diversus View in CoL vs 2<1< 3 in M. formosus View in CoL ), larger single pores on legs I–III (3.1 – 4.8 µm in M. diversus View in CoL sp. nov. vs 2.1 – 2.9 µm in M. formosus View in CoL ) and different shape of egg processes (screw-like processes within a membrane in M. diversus View in CoL sp. nov. vs single tip cones without a membrane in M. formosus View in CoL ).
M. furcatus View in CoL (according to the re–description, Binda & Pilato 1992) by: arrangement of pores on dorsal side of the body (randomly arranged pores in M. diversus View in CoL sp. nov. vs pores arranged in undefined number of transverse bands in M. furcatus View in CoL ), absence of tri- and quadrilobed cuticular pores, the presence of large pores on the frontal and caudal end, the presence of triangular formation of pores on the ventral side of the head, present of large pores on the external side of the legs I–III, presence of granulation on legs, absence of the oral cavity armature, a different macroplacoid length sequence (1=2 ≅ 3 in M. diversus View in CoL vs 2<3< 1 in M. furcatus View in CoL ), stylet supports inserted in a more anterior position ([ 50.5–52.9] in M. diversus View in CoL vs ca. [ 68.4] in M. furcatus View in CoL ) and a different shape of egg processes (screw-like processes within a membrane in M. diversus View in CoL sp. nov. vs single tip cones without a membrane in M. furcatus View in CoL ).
M. harrylewisi View in CoL by: arrangement of pores on dorsal side of the body (randomly arranged pores in M. diversus View in CoL sp. nov. vs pores arranged in 10 transverse bands in M. harrylewisi View in CoL ), absence of tri- and quadrilobed cuticular pores, the presence of triangular formation of pores on the ventral side of the head, a different macroplacoid length sequence (1=2 ≅ 3 in M. diversus View in CoL sp. nov. vs 2≤3< 1 in M. harrylewisi View in CoL ), slightly smaller body size (147 – 200 µm in M. diversus View in CoL sp. nov. vs 200.3 – 386.6 µm in M. harrylewisi View in CoL ), narrower external buccal tube (1.3 – 1.7 µm in M. diversus View in CoL sp. nov. vs 1.9 – 3.4 µm in M. harrylewisi View in CoL ), stylet supports inserted in a more anterior position ([ 50.5–52.9] in M. diversus View in CoL sp. nov. vs [ 61.4–67.6] in M. harrylewisi View in CoL ), shorter placoid row (5.3 – 6.9 µm [ 25.3–29.3] in M. diversus View in CoL sp. nov. vs 10.4 – 34.7 µm [ 34.6–46.0]) in M. harrylewisi View in CoL ) and a different shape of egg processes (screwlike processes within a membrane in M. diversus View in CoL sp. nov. vs single tip cones without a membrane in M. harrylewisi View in CoL ).
M. jonesorum View in CoL by: arrangement of pores on dorsal side of the body (randomly arranged pores in M. diversus View in CoL sp. nov. vs pores arranged in 10 transverse bands in M. jonesorum View in CoL ), absence of tri- and quadrilobed cuticular pores, the presence of triangular formation of pores on the ventral side of the head, a different macroplacoid length sequence (1=2 ≅ 3 in the new species vs 1<2<3 Μm in M. jonesorum View in CoL ), the presence of eyes, smaller body size (147–200 µm in M. diversus View in CoL sp. nov. vs 205.9–276.7 µm in M. jonesorum View in CoL ), slightly shorter buccal tube (19.8 – 23.9 µm in M. diversus View in CoL sp. nov. vs 24.4 – 29.6 µm in M. jonesorum View in CoL ), smaller buccal tube external width (1.3 – 1.7 µm in M. diversus View in CoL sp. nov. vs 2.1 – 2.6 µm in M. jonesorum View in CoL ), stylet supports inserted in a more anterior position ([ 50.5–52.9] in M. diversus View in CoL sp. nov. vs [ 63.0–65.6] in M. jonesorum View in CoL ), shorter macroplacoids II and III (mII: 1.1 – 1.7 µm [ 5.5–7.2]; mIII: 1.2 – 1.7 µm [ 6.0–7.1] in M. diversus View in CoL sp. nov. vs mII: 1.9 – 2.3 µm [ 7.1–8.8]; mIII: 2.4 – 2.6 µm [ 8.4–9.9] in M. jonesorum View in CoL ), shorter macroplacoid row (4.5 – 6.0 µm [ 22.3–25.8] in M. diversus View in CoL sp. nov. vs 7.0 – 8.4 µm [ 27.0–34.4] in M. jonesorum View in CoL ), the presence of a microplacoid and slightly shorter primary and secondary branches of external claws I–IV (see Table 2 View TABLE 2 below and Table 2 View TABLE 2 in Meyer et al. 2011 for the exact differences in dimensions of claws).
M. keppelensis View in CoL by: lack of red pigmented granules, arrangement of pores on dorsal side of the body (randomly arranged pores in M. diversus View in CoL sp. nov. vs pores arranged in 10 transverse bands in M. keppelensis View in CoL ), presence of a few larger pores on dorsal side of the head segment and caudal end, presence of triangular formation of pores on ventral side of the head segment, the presence of rows of pores between the legs, a different macroplacoid length sequence (1=2 ≅ 3 in M. diversus View in CoL sp. nov. vs 2=3< 1 in M. keppelensis View in CoL ), slightly shorter buccal tube (19.8 – 23.9 µm in M. diversus View in CoL sp. nov. vs 24.9 – 28.4 µm in M. keppelensis View in CoL ) slightly larger pt of buccal tube external width ([ 6.3–7.6] in M. diversus View in CoL sp. nov. vs ca. [ 5.8] in M. keppelensis View in CoL ), stylet supports inserted in a more anterior position ([ 50.5–52.9] in M. diversus View in CoL sp. nov. vs ca. [ 60.6] in M. keppelensis View in CoL ), shorter macroplacoid row (4.5 – 6.0 µm in M. diversus View in CoL sp. nov. vs 7.0 – 7.6 µm in M. keppelensis View in CoL ), and different shape of egg processes (screw-like processes within a membrane in M. diversus View in CoL sp. nov. vs short cones with pointed apices in M. keppelensis View in CoL ).
M. orthofasciatus View in CoL by: arrangement of pores on dorsal side of the body (randomly arranged pores in M. diversus View in CoL sp. nov. vs pores arranged in 11 transverse bands in M. orthofasciatus View in CoL ), absence of tri- and quadrilobed cuticular pores, the presence of triangular formation of pores on the ventral side of the head, the presence of large pores on the external side of legs I – III, the presence of rows of pores between the legs, the presence of granulation on all the legs, stylet supports inserted in a more anterior position ([ 50.5–52.9] in M. diversus View in CoL sp. nov. vs [ 66.5–67.8] in M. orthofasciatus View in CoL ), shorter macroplacoids I and III (mI: 1.1 – 1.7 µm [ 5.5–7.1]; mIII: 1.2 – 1.7 µm [ 6.0–7.1] in M. diversus View in CoL sp. nov. vs mI: 1.9 – 3.1 µm [ 9.5–13.7]; mIII: 1.9 – 2.4 µm [ 9.5–10.7] in M. orthofasciatus View in CoL ), slightly shorter macroplacoid row (4.5 – 6.0µm [ 22.3–25.8] in M. diversus View in CoL sp. nov. vs 6.1 – 7.4 µm [ 30.7–34.1] in M. orthofasciatus View in CoL ), slightly lower number of processes on egg circumference ( 21–22 in M. diversus View in CoL sp. nov. vs ca. 24 in M. orthofasciatus View in CoL ), shorter egg processes (3.1 – 3.6 µm in M. diversus View in CoL sp. nov. vs 4.2 – 4.5 µm in M. orthofasciatus View in CoL ) narrower process bases (1.2 – 1.8 µm in M. diversus View in CoL sp. nov. vs 2.3 – 2.7 µm in M. orthofasciatus View in CoL ).
M. poricinctus View in CoL by: arrangement of pores on dorsal side of the body (randomly arranged pores in M. diversus View in CoL sp. nov. vs pores arranged in eight transverse bands in M. poricinctus View in CoL ), the presence of triangular formation of pores on the ventral side of the head, the presence of large pores on the frontal and caudal end, the presence of rows of pores between the legs, a different macroplacoid length sequence (1=2 ≅ 3 in M. diversus View in CoL sp. nov. vs 2=3< 1 in M. poricinctus View in CoL ), stylet supports inserted in a more anterior position ([ 50.5–52.9] in M. diversus View in CoL sp. nov. vs ca. [ 59.5] in M. poricinctus View in CoL ), slightly shorter macroplacoid row (4.5 – 6.0 µm [ 22.3–25.8] in M. diversus View in CoL sp. nov. vs 6.0 – 8.9 µm [ ca. 30.9] in M. poricinctus View in CoL ), smaller diameter of eggs with processes (51.5 – 53.9 µm in M. diversus View in CoL sp. nov. ca. 60.0 µm in M. poricinctus View in CoL ), shorter egg processes (3.1 – 3.6 µm in M. diversus View in CoL sp. nov. vs 6.5 – 7.0 µm in M. poricinctus View in CoL ) and smaller diameter of the distal discs of the processes (3.0–3.8 µm in M. diversus View in CoL sp. nov. vs ca. 5.5 µm in M. poricinctus View in CoL ).
M. pustulatus View in CoL by: absence of triangular and polygonal pores, smaller pores on dorsal side (2.2 – 3.2 Μm in diameter on frontal and caudal end in M. diversus View in CoL vs 4 – 5 Μm in diameter on frontal side and 6 – 7 Μm in diameter on caudal end in M. pustulatus View in CoL ), the presence of triangular formation of pores on ventral side of the head, the presence of rows of pores between the legs and a different shape of the egg processes (screw-like processes within a membrane in M. diversus View in CoL sp. nov. vs single tip cones without a membrane in M. pustulatus View in CoL ).
M. ramazzottii View in CoL by: arrangement of pores on dorsal side of the body (randomly arranged pores in M. diversus View in CoL sp. nov. vs pores arranged in undefined number of transverse bands in M. ramazzottii View in CoL ), the presence of large pores on the frontal and caudal end, the presence of triangular formation of pores on the ventral side of the head, the presence of large pores on legs I – III, the presence of rows of pores between the legs, absence of the oral cavity armature, a different macroplacoid length sequence (1=2 ≅ 3 in the new species vs 3<2<1 Μm in M. ramazzottii View in CoL ), shorter buccal tube (19.8 – 23.9 µm in M. diversus View in CoL sp. nov. vs ca. 35.5 µm in M. ramazzottii View in CoL ), smaller total buccal tube width (0.5 – 1.7 µm in M. diversus View in CoL sp. nov. vs ca. 4.1 µm in M. ramazzottii View in CoL ), stylet supports inserted in a more anterior position ([ 50.5–52.9] in M. diversus View in CoL sp. nov. vs [ 68.2–68.3] in M. ramazzottii View in CoL ), shorter placoids (mI/mII: 1.1–1.7 µm [ 5.5–7.1]; mIII: 1.2–1.7 µm [ 6.0–7.1] in M. diversus View in CoL sp. nov. vs mI: ca. 5.0 µm [ 14.0]; mII: ca. 4.7 µm [ 13.1]; mIII: 4.6 µm [ 12.8] in M. ramazzottii View in CoL ), smaller microplacoid (0.4–0.7 µm [ 2.0–3.0] in M. diversus View in CoL sp. nov. vs ca. 2.3 µm [ ca. 6.4] in M ramazzottii View in CoL ) and shorter external primary branches of claws I, II, (I: 4.6–6.0 µm [ 20.3–26.6]; II: 4.7–6.6 µm [ 23.5–28.4] in M. diversus View in CoL sp. nov. vs I: 10.7 µm [ 30.0]; II: 12.5 µm [ 35.2], in M. ramazzottii View in CoL ).
M. vinciguerrae View in CoL by: the presence of large pores on the frontal and caudal end, the presence of triangular formation of pores on ventral side of the head, the presence of large pores on the external side of legs I – III, the presence of rows of pores between the legs, absence of the oral cavity armature, a different macroplacoid length sequence (1=2 ≅ 3 in the new species vs 2<3<1 Μm in M. vinciguerrae View in CoL ), smaller body size (147 – 200 µm in M. diversus View in CoL sp. nov. vs 380 – 520 µm in M. vinciguerrae View in CoL ), stylet supports inserted in a more anterior position ([ 50.5–52.9] in M. diversus View in CoL sp. nov. vs [ 66.1–68.18] in M. vinciguerrae View in CoL ), shorter macroplacoids (mI/mII: 1.1–1.7 µm [ 5.5–7.1]; mIII: 1.2–1.7 µm [ 6.0–7.1] in M. diversus View in CoL sp. nov. vs mI: ca. 4.4 µm [ ca. 12.4]; mII: ca. 3.6 µm [ ca. 10.13]; mIII: ca. 3.6 µm [ ca. 10.33] in M. vinciguerrae View in CoL ), shorter microplacoid (0.4–0.7 µm [ 2.0–3.0] in M. diversus View in CoL sp. nov. vs mi: ca. 1.9 µm [ ca. 5.4] in M. vinciguerrae View in CoL ), shorter placoid row (5.3 – 6.9 µm [ 25.3–29.3] in M. diversus View in CoL sp. nov. vs ca. 15.4 µm [ ca. 43.8] in M. vinciguerrae View in CoL ), shorter primary branches of external/anterior claws II, III, IV (II: 4.7 – 6.6 µm [ 23.5–28.4]; III: 4.5 – 6.2 µm [ 22.5–27.8]; IV: 4.8 – 7.0 µm [ 24.0–30.2] in M. diversus View in CoL sp. nov. vs II: ca. 12.3 µm [ ca. 35.1]; III: ca. 12.6 µm [ ca. 36.0]; IV: 14.6 µm [ ca. 41.6] in M. vinciguerrae View in CoL ) and a different shape of egg processes (screw-like processes within a membrane in M. diversus View in CoL sp. nov. vs cones without a membrane in M. vinciguerrae View in CoL ).
M. xavieri View in CoL by: arrangement of pores on the dorsal side of the body (randomly arranged pores in M. diversus View in CoL sp. nov. vs pores arranged in nine wide transverse bands in M. xavieri View in CoL ), the absence of trilobed pores, the presence of differently sized pores (all pores of similar size in M. xavieri View in CoL ), the presence of large pores on the frontal and caudal end, the presence of triangular formation of pores on the ventral side of the head, the presence of rows of pores between the legs, the presence of granulation on legs, a different macroplacoid sequence (1=2⊋ ≅ 3 in M. diversus View in CoL sp. nov. vs 2<3<1 Μm in M. xavieri View in CoL ), smaller body size (147–200 µm in M. diversus View in CoL sp. nov. vs 275–410 µm in M. xavieri View in CoL ), a shorter buccal tube (19.8–23.9 µm in M. diversus View in CoL sp. nov. vs 27.5–32.7 µm in M. xavieri View in CoL ), a narrower buccal tube (1.3 – 1.7 µm [ 6.3–7.6] in M. diversus View in CoL sp. nov. vs 2.4 – 3.1 µm [ 8.3–9.8] in M. xavieri View in CoL ), stylet supports inserted in a more anterior position ([ 50.5–52.9] in M. diversus View in CoL sp. nov. vs [ 66.1–67.9] in M. xavieri View in CoL ), shorter macroplacoids (mI/mII: 1.1–1.7 µm [ 5.5–7.1]; mIII: 1.2–1.7 µm [ 6.0–7.1] in M. diversus View in CoL sp. nov. vs mI: 3.6 – 4.5 µm [ 12.7–13.8]; mII: 2.9 – 3.6 µm[ 10.3–11.1]; mIII: 3.0 – 3.9 µm [ 10.9–11.9] in M. xavieri View in CoL ), a shorter macroplacoid row (4.5 – 6.0 µm [ 22.3–25.8] in M. diversus View in CoL sp. nov. vs 9.8 – 12.6 µm [ 35.6–38.5] in M. xavieri View in CoL ), smaller microplacoid (0.4 – 0.7 µm [ 2.0–3.0] in M. diversus View in CoL sp. nov. vs 1.5 – 2.0 µm [ 5.0–6.2] in M. xavieri View in CoL ), shorter placoid row (5.3 – 6.9 µm [ 25.3–29.3] in M. diversus View in CoL sp. nov. vs 10.9 – 13.9 µm [ 39.6–43.3] in M. xavieri View in CoL ), different lengths of external/anterior and internal/posterior claws II – IV (see Table 3 View TABLE 3 below and Table 2 View TABLE 2 in Fontoura et al. (2009b) for the exact differences in dimensions of claws) and different shape of egg processes (screw-like processes within a membrane in M. diversus View in CoL sp. nov. vs long cones with bi- or multi-tipped tips and without a membrane in M. xavieri View in CoL ).
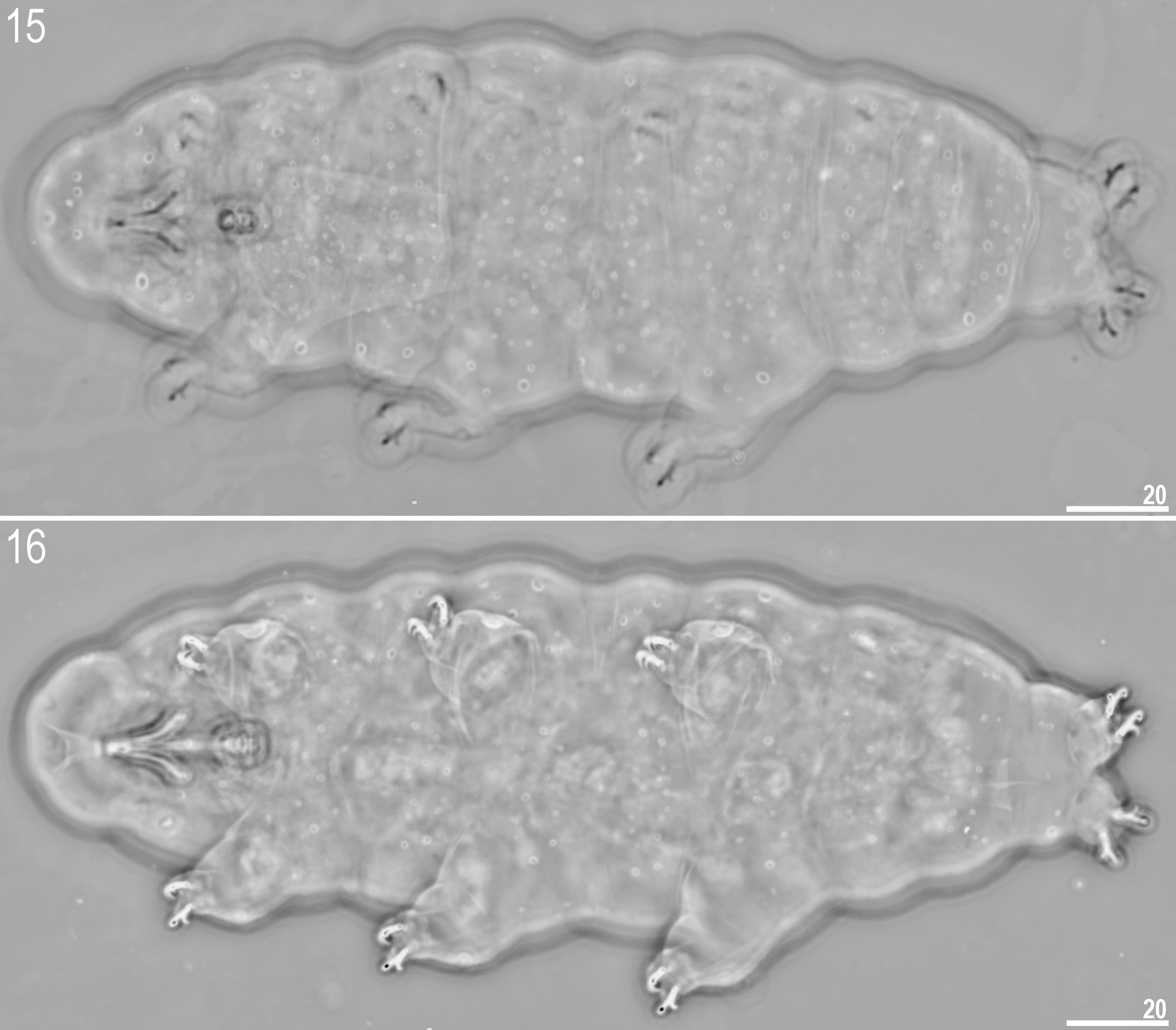

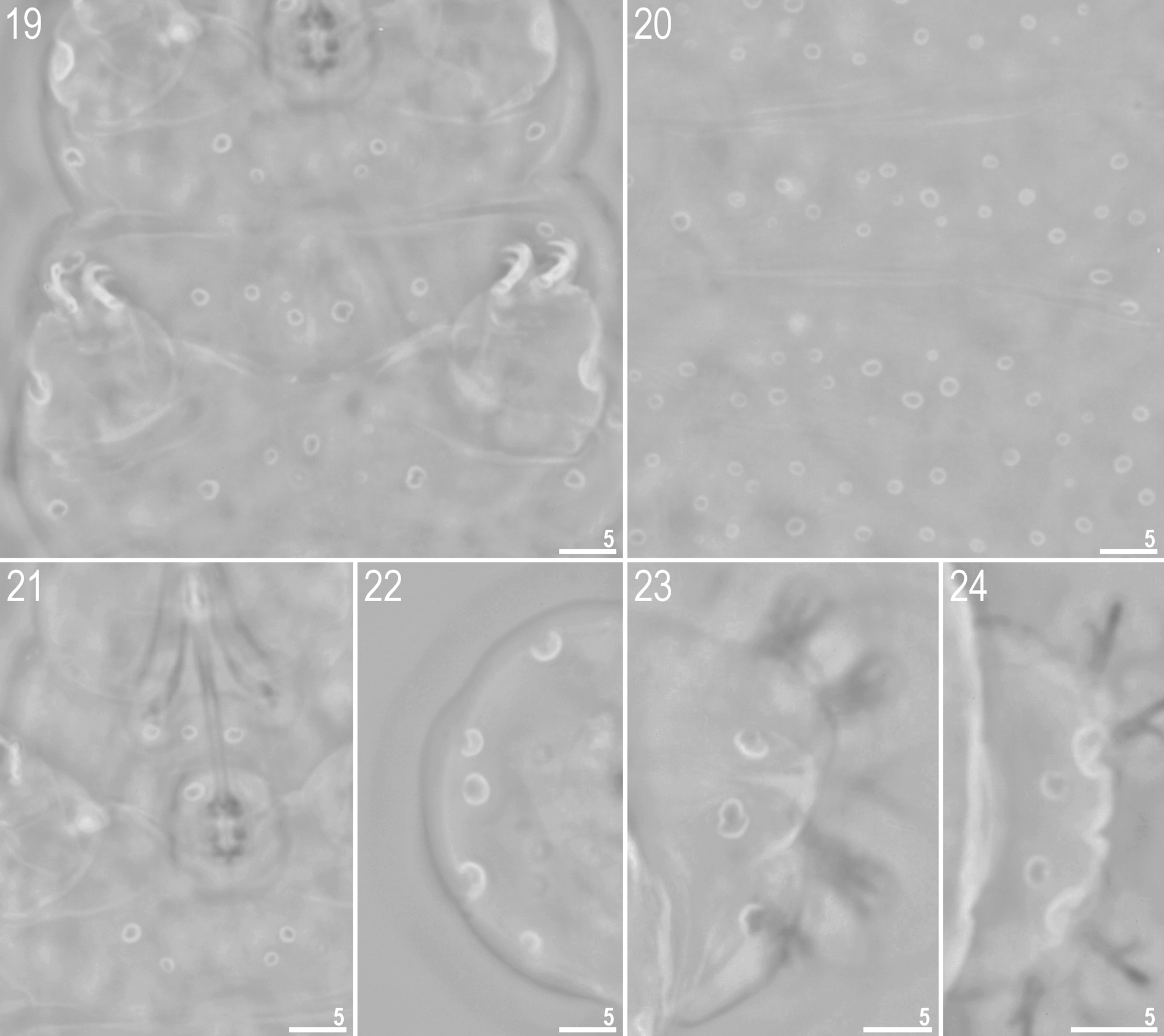

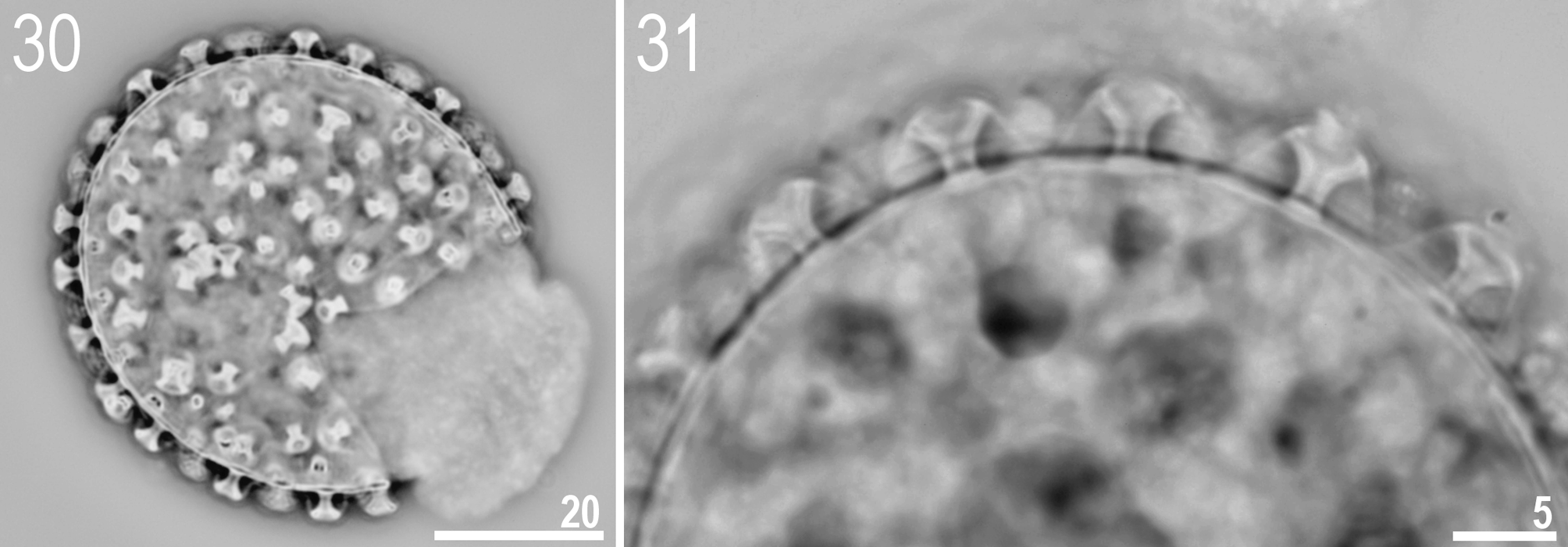
TABLE 3. Measurements and pt values of selected morphological structures of fifteen specimens from the type population of Minibiotus diversus sp. nov. mounted in Hoyer’s medium (N—number of specimens / structures measured, RANGE refers to the smallest and the largest structure among all measured specimens; SD—standard deviation).
| CHARACTER | N | RANGE | MEAN | SD | Holotype |
|---|---|---|---|---|---|
| µm pt | µm pt | µm pt | µm pt | ||
| Body length | 15 | 147–200 739–862 | 173 784 | 15 35 | 200 862 |
| Buccal tube | |||||
| Length | 15 | 19.8–23.9 - | 22.1 – | 1.3 – | 23.2 – |
| Stylet support insertion point | 15 | 10.0–12.4 50.5–52.9 | 11.5 51.8 | 0.8 0.8 | 12.0 51.7 |
| External width | 15 | 1.3–1.7 6.3–7.6 | 1.5 6.8 | 0.1 0.4 | 1.5 6.5 |
| Internal width | 15 | 0.5–0.7 2.1–3.0 | 0.6 2.6 | 0.1 0.3 | 0.6 2.6 |
| Ventral lamina length | 0 | ?? | ?? | ?? | ?? |
| Placoid lengths | |||||
| Macroplacoid 1 | 15 | 1.1–1.7 5.5–7.1 | 1.4 6.3 | 0.2 0.5 | 1.6 6.9 |
| Macroplacoid 2 | 15 | 1.1–1.7 5.5–7.2 | 1.4 6.5 | 0.2 0.4 | 1.6 6.9 |
| Macroplacoid 3 | 15 | 1.2–1.7 6.0–7.1 | 1.4 6.5 | 0.1 0.4 | 1.6 6.9 |
| Microplacoid | 15 | 0.4–0.7 2.0–3.0 | 0.6 2.6 | 0.1 0.4 | 0.7 3.0 |
| Macroplacoid row | 15 | 4.5–6.0 22.3–25.8 | 5.3 24.0 | 0.4 1.2 | 5.9 25.4 |
| Placoid row | 15 | 5.3–6.9 25.3–29.3 | 6.1 27.7 | 0.5 1.3 | 6.8 29.3 |
| Claw 1 lengths | |||||
| External base | 0 | ?? | ?? | ?? | ?? |
| External primary branch | 14 | 4.6–6.0 20.3–26.6 | 5.3 24.2 | 0.5 1.8 | 5.8 25.0 |
| External secondary branch | 12 | 3.0–4.2 14.9–19.3 | 3.6 16.3 | 0.4 1.2 | 3.9 16.8 |
| Internal base | 0 | ?? | ?? | ?? | ?? |
| Internal primary branch | 13 | 4.2–5.9 19.7–26.1 | 5.2 23.5 | 0.5 1.9 | 5.5 23.7 |
| Internal secondary branch | 11 | 3.0–3.9 12.9–16.8 | 3.3 15.2 | 0.3 1.3 | ?? |
| Claw 2 lengths | |||||
| External base | 0 | ?? | ?? | ?? | ?? |
| External primary branch | 14 | 4.7–6.6 23.5–28.4 | 5.7 25.7 | 0.5 1.6 | 6.6 28.4 |
| External secondary branch | 14 | 3.3–4.6 15.0–19.8 | 3.9 17.8 | 0.4 1.3 | 4.6 19.8 |
| Internal base | 0 | ?? | ?? | ?? | ?? |
| Internal primary branch | 14 | 4.8–6.0 21.6–26.4 | 5.4 24.6 | 0.4 1.5 | 5.7 24.6 |
| Internal secondary branch | 13 | 2.9–4.5 13.7–20.2 | 3.7 16.8 | 0.5 1.9 | ?? |
| Claw 3 lengths | |||||
| External base | 0 | ?? | ?? | ?? | ?? |
| External primary branch | 14 | 4.5–6.2 22.5–27.8 | 5.7 25.6 | 0.4 1.7 | 6.2 26.7 |
| External secondary branch | 11 | 2.9–4.6 14.5–20.6 | 3.9 17.5 | 0.5 2.0 | 4.4 19.0 |
| Internal base | 0 | ?? | ?? | ?? | ?? |
| Internal primary branch | 14 | 4.8–5.8 21.1–26.6 | 5.4 24.1 | 0.3 1.4 | 5.6 24.1 |
| Internal secondary branch | 13 | 2.9–4.1 12.4–19.2 | 3.6 16.3 | 0.4 1.9 | ?? |
| Claw 4 lengths | |||||
| Anterior base | 0 | ?? | ?? | ?? | ?? |
| Anterior primary branch | 12 | 4.8–7.0 24.0–30.2 | 6.0 27.4 | 0.6 2.1 | 7.0 30.2 |
| Anterior secondary branch | 12 | 3.6–5.4 17.1–23.3 | 4.3 19.4 | 0.5 1.8 | 5.4 23.3 |
| Posterior base | 0 | ?? | ?? | ?? | ?? |
| Posterior primary branch | 12 | 5.1–7.1 24.7–31.8 | 6.1 27.7 | 0.6 1.9 | 6.5 28.0 |
| Posterior secondary branch | 12 | 3.2–5.0 16.0–22.4 | 4.2 19.3 | 0.5 1.8 | 4.9 21.1 |
TABLE 4. Measurements of Minibiotus diversus sp. nov. eggs mounted in Hoyer’s medium (N—number of specimens / structures measured, RANGE refers to the smallest and the largest structure among all measured specimens; SD—standard deviation).
| CHARACTER | N | RANGE | MEAN | SD |
|---|---|---|---|---|
| Diameter of egg without processes | 3 | 44.9–47.8 | 46.1 | 1.5 |
| Diameter of egg with processes | 3 | 51.5–53.9 | 52.8 | 1.2 |
| Process height | 9 | 3.1–3.6 | 3.3 | 0.2 |
| Process base width | 9 | 1.2–1.8 | 1.5 | 0.2 |
| Process base/height ratio | 9 | 39%–51% | 44% | 5% |
| Terminal disc width | 9 | 3.0–3.8 | 3.5 | 0.3 |
| Distance between processes | 9 | 3.6–4.8 | 4.3 | 0.4 |
| Number of processes on the egg circumference | 3 | 21–22 | 21.3 | 0.6 |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Minibiotus diversus
| Roszkowska, Daniel Adrian Ciobanu Milena & Kaczmarek, Łukasz 2015 |
M. formosus
| Zawierucha et al. 2014 |
M. jonesorum Meyer et al ., 2011
| Meyer et al. 2011 |
M. harrylewisi
| Meyer & Hinton 2009 |
M. orthofasciatus Fontoura et al ., 2009a
| Fontoura et al. 2009 |
M. xavieri Fontoura et al ., 2009b
| Fontoura et al. 2009 |
M. eichhorni
| Michalczyk & Kaczmarek 2004 |
M. keppelensis
| Claxton 1998 |
M. poricinctus
| Claxton 1998 |
M. ramazzottii
| Binda & Pilato 1992 |
M. vinciguerrae
| Binda & Pilato 1992 |
M. bisoctus (
| Horning et al. 1978 |
M. pustulatus (
| Ramazzotti 1959 |
M. furcatus (
| Ehrenberg 1859 |