
Pilophorus Hahn, 1826
|
publication ID |
https://doi.org/10.11646/zootaxa.4942.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:CDF398FE-B0F6-40E6-967D-FB857C1565BD |
|
DOI |
https://doi.org/10.5281/zenodo.4618372 |
|
persistent identifier |
https://treatment.plazi.org/id/A16ADB37-EB00-0C78-FF47-C3A47FB9FA82 |
|
treatment provided by |
Plazi |
|
scientific name |
Pilophorus Hahn |
| status |
|
Genus Pilophorus Hahn View in CoL
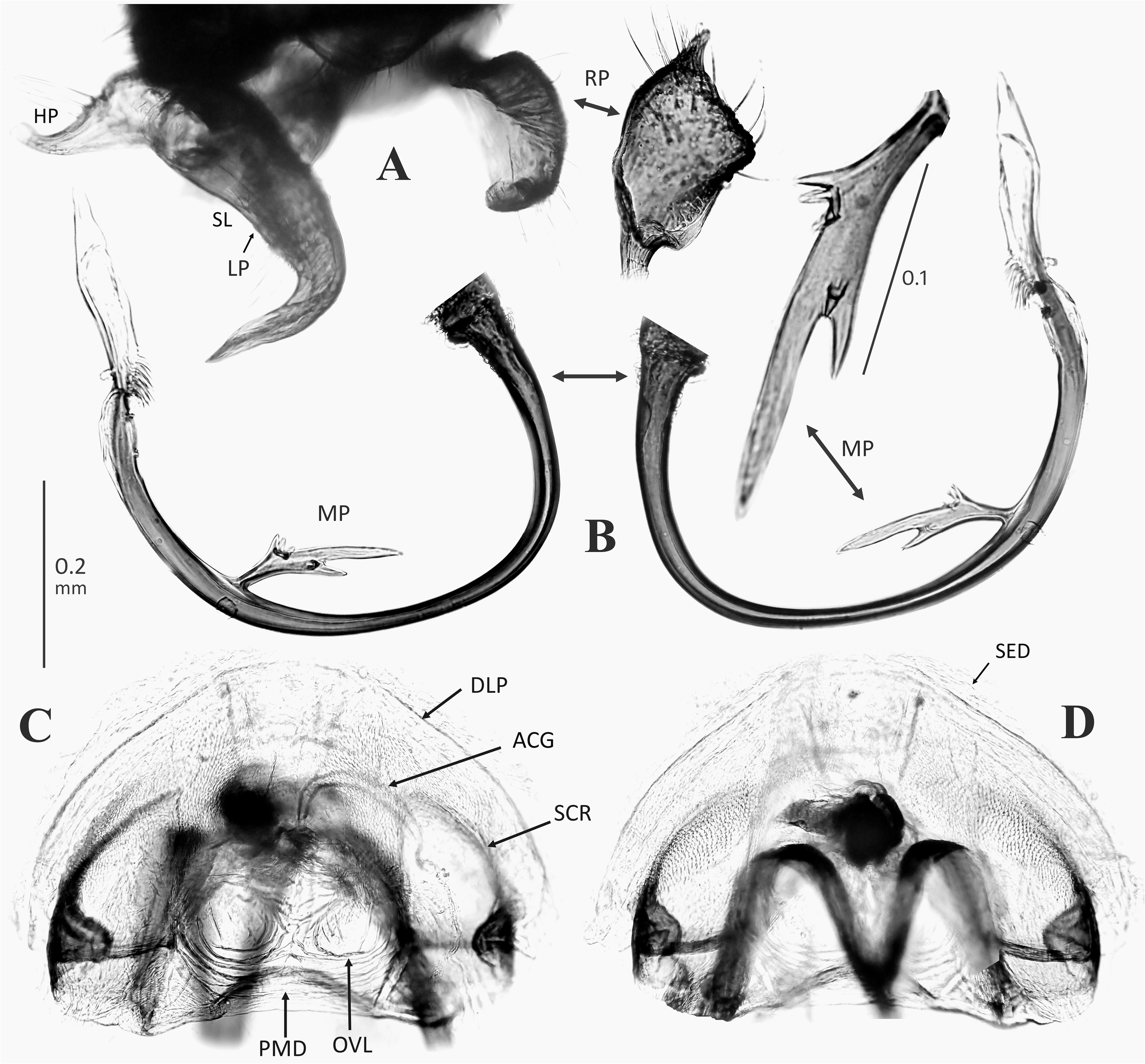
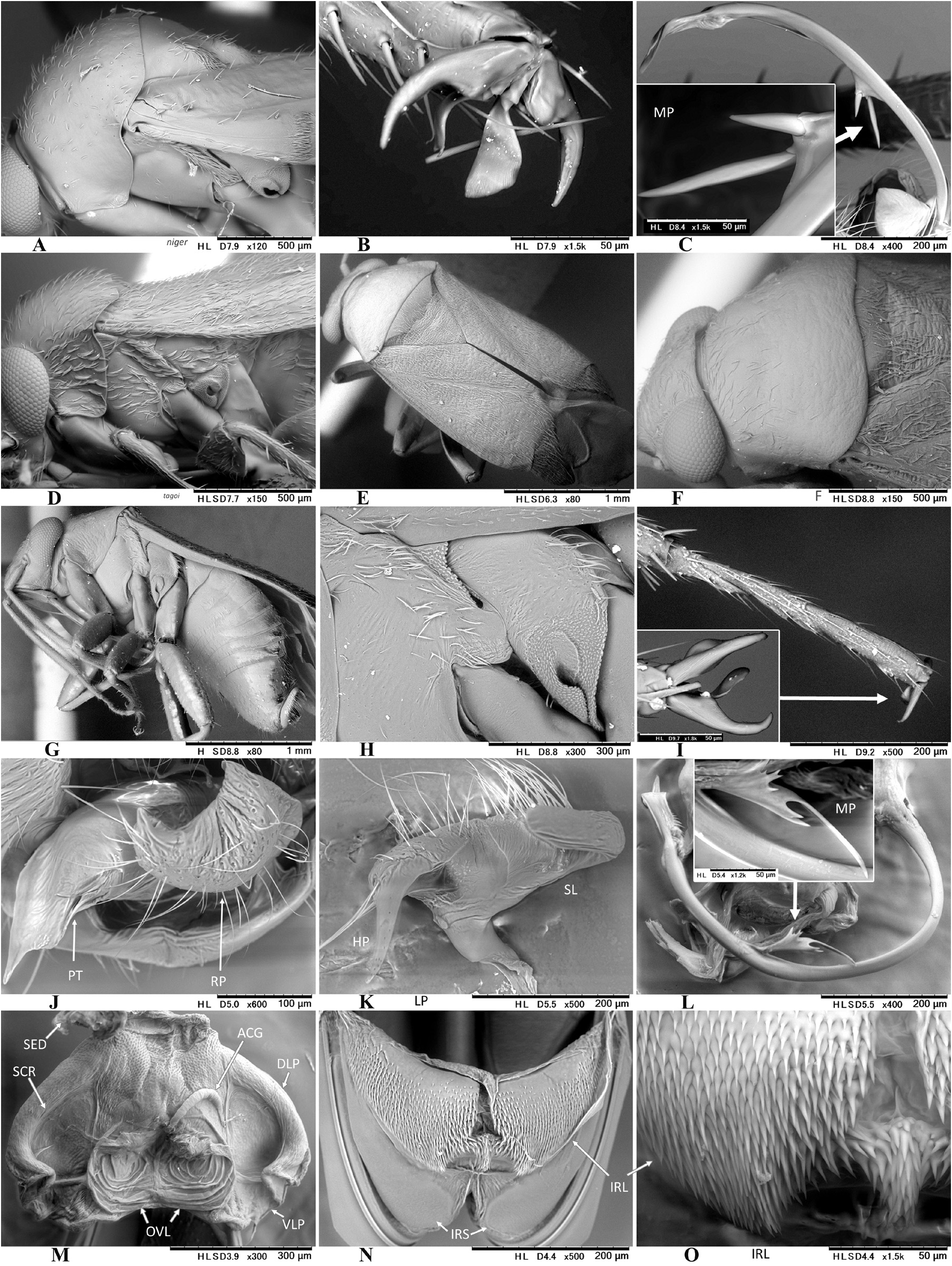
Diagnosis: Asian Pilophorus species are recognized by the following characters: Small to moderate size (2.2̅ 4.8 mm in total length); usually antlike in overall appearance, elongate (rarely body ovoid or elongate-oval as in P. miyamotoi , P. tagoi and P. validicornis , and P. yunganensis Schuh, 1984 from continental China) and always macropterous body form; partly clustered scale-like setae on scutellum and mesepimeron (cf. Schuh, 1984); presence of median band of scale-like setae on corium (posterior band variable, sometimes scattered, or partly zigzag or interrupted); often weakly curved metafemur; fleshy, apically convergent parempodia; rounded (cf. Fig. 3D View FIGURE 3 ) or linear ( Fig. 5C View FIGURE 5 ) clustered scale-like setae on anterolateral part of abdomen (sterna III and/or IV); usually widened, more or less splayedout left paramere; ovoid to elongate-oval right paramere; C- to J-shaped (rarely nearly straight) endosoma often bearing a simple, flagellate or branched median process (MP); thick-rimmed sclerotized ring (SCR); and minutely spinulate interramal lobe (IRL) [some characters in the male and female genitalia are shared by other genera (e.g., Hypseloecus , Pherolepis ) (cf. Figs 13 View FIGURE 13 , 20 View FIGURE 20 J–O)]. Schuh (1984, 1991), Schuh & Schwartz (1988), Yasunaga & Schuh (2013) and Yasunaga et al. (2014) provided additional diagnostic characters. Fukuda et al. (2020) also proposed the nymphal ‘metanotal ridge’ (in 2nd–5th instars) as a synapomorphy shared by Pilophorus members.
Distribution. The majority of species are known from the Holarctic and Oriental Regions, with three Ethiopian spp. and one Australian species, P. perplexus Douglas & Scott , possibly introduced from Europe ( Malipatil et al., 2020). The generic placement of the Ethiopian species, P. linnavuorii , needs verification (see discussion below).
Discussion. Pilophorus is composed of morphologically very diverse species. In SE Asia, some members are ‘strikingly’ antlike because of conspicuous modification of the pronotum (cf., P. barbiger Yasunaga & Schuh, 2013 , Indochina) or the scutellum (cf., P. aurifasciatus Nakatani & Komatsu, 2013 , Malaysia). Most members of Pilophorus co-occur with certain ant species ( Yasunaga 2001; Nakatani et al. 2013; Yasunaga & Schuh 2013) and are assumed to be correlated by virtue of their myrmecomorphy. In temperate and subboreal climate zones of Asia, ‘strikingly ant-mimetic’ members are not present, and most congeners have the fiddle-shaped or panduriform bodies which are more or less constricted in the middle of the hemelytra. On the other hand, P. tagoi ( Fig. 6C View FIGURE 6 ) uniquely has an ovoid body shape and is not antlike (HCR 0.81̅0.89). This Japanese Cryptomeria specialist is externally similar to certain member of the eastern Palearctic genus Pherolepis rather than Pilophorus (as in couplet 4 of above key and Fig. 6D, F, G View FIGURE 6 ). Pherolepis species usually have the larger body, uniformly brownish coloration, almost linear antennal segment II, less scale-like setae on the thoracic pleura and abdomen, and shiny hemelytron ( Fig. 6 View FIGURE 6 D–F, H). Known host associations of Pherolepis spp. are restricted to deciduous broadleaf trees so far ( Kerzhner, 1988; Yasunaga, 2001; Yasunaga & Duwal 2016), except for a Chinese pine-inhabitant, Ph. robustus Zhang & Liu ( Zhang & Liu, 2009) .
Judging from the similar shape of the male and female genitalia, Pilophorus , Pherolepis and Hypseloecus evidently constitute a monophyletic group (Schuh & Menard, 2013; Yasunaga & Duwal, 2016). Hypseloecus can be distinguished readily from other two genera by the unique host association with Loranthaceae and Santalaceae mistletoes (Schuh & Menard, 2011) and possession of a pair of pleural glands ( sensu Yasunaga & Duwal, 2019). However, almost all the other diagnostic characters of Hypseloecus are shared by Pilophorus and Pherolepis . Additionally, both latter genera are equally composed of conifer- and broadleaf-inhabiting species. Some Pilophorus species, e.g. P. tagoi Yasunaga & Duwal and P. yunganensis Schuh , have the ovoid or elongate-oval body form of Pherolepis members. We cannot suggest any morphological character unequivocally demarcating each genus. Pherolepis species are currently supposed to be principally phytophagous, and their nymphal forms lack the metanotal ridge ( sensu Fukuda et al. 2020); however, nymphal forms were confirmed only for Ph. fasciatus ( Yasunaga, 2001) and Ph. hizenicus n. sp. (cf. Fig. 6H View FIGURE 6 ). Although Pherolepis is considered to be synonymous with Pilophorus based on available evidence, the definitive treatment is beyond a scope of this work and will depend upon the acquisition of DNA sequence data for many relevant pilophorine taxa.
Pilophorus members are currently known from the Palearctic (except for the Australian Region) and Nearctic Regions. Three species were described from the Ethiopian Region, P. lestoni Schuh, 1989 , P. linnavuorii Schuh, 1989 and P. pilosus Odhiambo, 1959 (cf. Schuh, 2002 –2013); however, judging from its ovoid, non-antlike body form (habitus image available on website: https://www.discoverlife.org/mp/20q?search= Pilophorus +linnavuori), P. linnavuorii may eventually require placement in Hypseloecus .
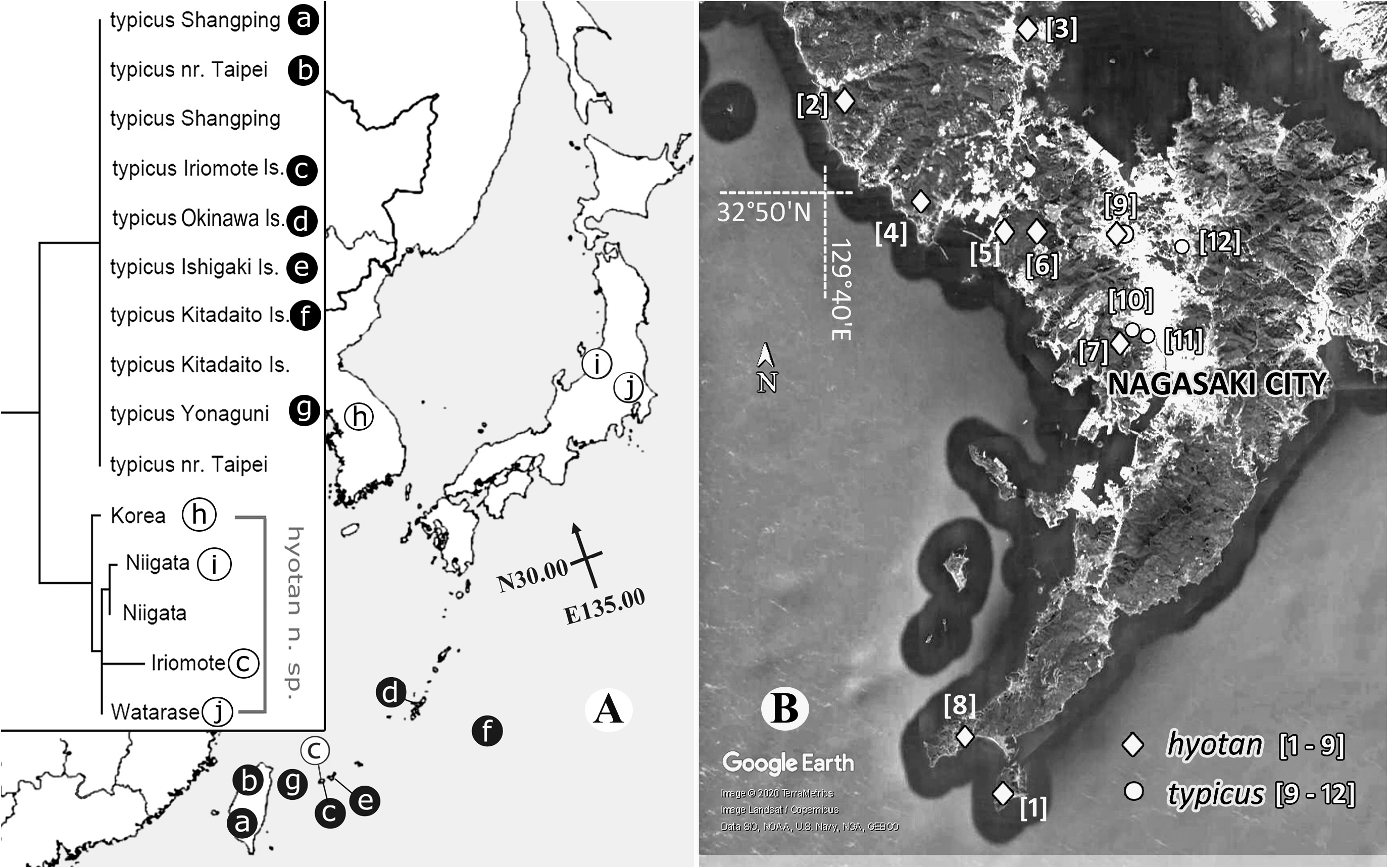
The Japanese fauna of Pilophorus now comprises 17 species. The majority are known to inhabit various plants (of more than two plant families) and do not appear to be host plant specific, except for P. choii , P. lucidus and P. pseudoperplexus restricted to fagaceous oaks (mostly Quercus spp.), conifer specialists— P. miyamotoi , P. nakatanii , P. tagoi and P. varidicornis , and P. okamotoi currently found on a herbaceous Artemisia species. Nonetheless, almost of all Pilophorus species are posited to be predaceous (cf. Yasunaga & Duwal, 2016), and the pilophorine host plant associations are presumed to be influenced by the presence of acceptable prey organisms ( Schuh & Schwartz, 1988). Immature forms of Pilophorus hyotan n. sp. ( Figs 4 View FIGURE 4 C–H), P. setulosus and P. typicus inhabit a variety of plant taxa (see above checklist and biology section of each species). When laboratory reared, the mature females of both P. hyotan n. sp. and P. typicus easily oviposit into the leaves and stems of Kalanchoe daigremontiana (native to Madagascar and now widespread as a foliage houseplant) ( Fukuda et al., 2020 and present observation, cf. Fig. 4B View FIGURE 4 ). Pilophorus hyotan n. sp. and P. typicus are considered to be potential candidates for IPM programs of economically important pests in agro-ecosystems (e.g., Ito et al., 2009, 2011; Yasunaga, 2001; Fukuda et al., 2020). However, many previous reports concerning ‘ P. typicus ’ from Japan and Korea (cf. Fig. 8A View FIGURE 8 ) were obviously confused with P. hyotan n. sp. (cf. Yasunaga, 2001; Komi, 2016).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.