
Pseudolimea arctica ( Zakharov, 1966 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3859.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:24FCAAE1-AB7C-4FAD-8698-D0C9F12400EC |
|
DOI |
https://doi.org/10.5281/zenodo.5228257 |
|
persistent identifier |
https://treatment.plazi.org/id/A2311D4D-9F02-E339-04E6-F884FC952EDB |
|
treatment provided by |
Felipe |
|
scientific name |
Pseudolimea arctica ( Zakharov, 1966 ) |
| status |
|
Pseudolimea arctica ( Zakharov, 1966)
( Figure 10 P–R View FIGURE 10 )
1966 Lima (Pseudolimea) arctica sp.—Zakharov, p. 62, pl. 16, figs. 6–11.
1966 Pseudolimea aff. parallela Orbigny [sic]—Zakharov, p. 64, pl. 16, fig. 12.
1982 Pseudolimea cf. arctica (Zakharov) —Fürsich, p. 61, fig. 25 A.
2011 Pseudolimea sp. —Hammer et al., fig. 7k, tab. 2.
Material examined. 18 specimens, all single valves; shells and internal moulds. See Appendix 1 for the list of specimens.
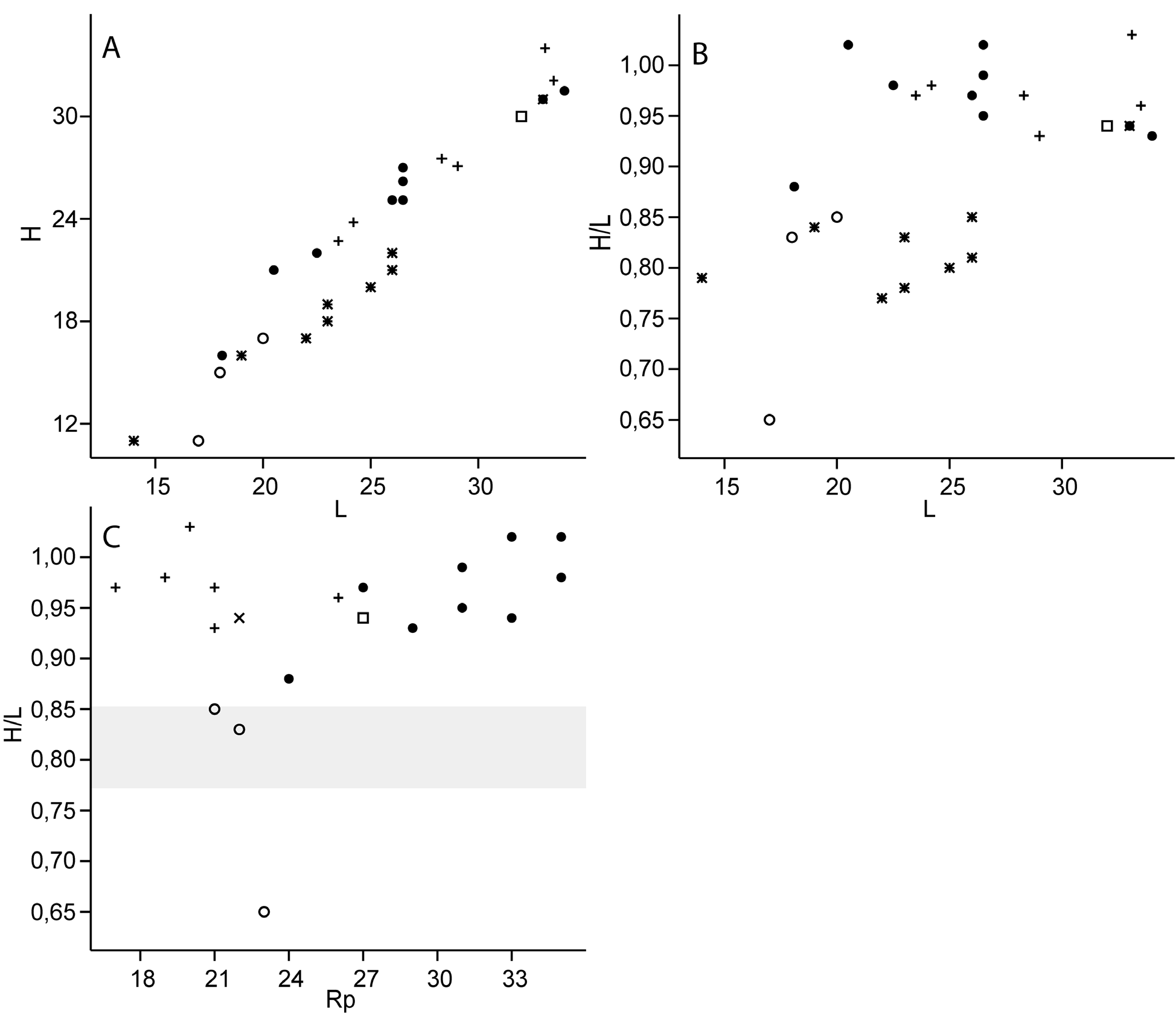
Dimensions. 18.1–34 mm in length, 16–31.5 mm in height. See Figure 12 A–C View FIGURE 12 and Appendix 2I for details.
Diagnosis. A Pseudolimea with up to 35 radial, v-shaped primary ribs with weaker ribs on anterior and posterior parts of the shell. Inter-rib spaces with single smaller riblet.
Description. Shell small to medium, up to 34 mm long and 31.5 mm high. Shell oval, inequilateral, posteroventrally elongated, triangular, with umbonal angle around 90°. Anterodorsal margin weakly convex, passing abruptly into broadly arcuate ventral margin. Posteroventral margin tightly arcuate, passing gently into straight and long posterodorsal margin. Posterodorsal margin about twice length of anterodorsal margin. Hinge line straight, around quarter shell length, with smaller anterior and larger posterior auricles not projecting beyond shell outline. Umbo small, weakly prosogyrate beak projecting beyond hinge line. Shell ornament of 24–35 v-shaped primary ribs with intercalated weaker secondary riblets in interspaces. Radial ornament dissected by weak, commarginal growth lines visible on outer shell surface only. Internal moulds show only rounded primary ribs. Ribs start on umbo, getting progressively stronger towards ventral margin; strongest on shell mid-flank, becoming weaker close to anterior and posterior shell margins. Ribs fade and then disappear on shell anterior where ventral curvature bends up towards anterodorsal margin; on posterior margin ribs disappear where posteroventral curvature passes into straight posterodorsal shell margin. Dorsal margins and auricles ornamented by concentric growth lines only. Dentition not observed.
Remarks. We include this species in the Siberian Kimmeridgian–Volgian taxon Pseudolimea arctica ( Zakharov, 1966) , based on its external shell proportions and the number of ribs. Comparison of shell measurments ( Fig. 12 A–C View FIGURE 12 ) shows that the Svalbard Pseudolimea specimens and P. arctica have an almost identical shell shape. The Svalbard specimens are more densely ribbed than P. arctica ( Zakharov, 1966) . However, more sparsely ribbed Svalbard specimens form a morphological continuity with the densely ribbed specimens of P. arctica , so there is no reason to separate them on rib number. Coarsely ribbed P. arctica specimens from Siberia overlap in rib number with P. multicostata Fürsich, 1982 , from England, Greenland and Siberia (Tab. 10, Zakharov 1966; Fürsich 1982; Kelly 1984) and with Lima (Mantellum) parallela (J. Sowerby, 1812) from the Aptian–Albian of England ( Woods 1904), which has no more than 23 ribs. However, both P. multicostata and P. parallela are more elongated than P. arctica from Siberia and Svalbard ( Fig. 12 B, C View FIGURE 12 ). Interestingly, Lima (Pseudolimea) aff. parallela (J. Sowerby, 1812) non (d’Orbigny), from the upper Ryazanian of Siberia ( Zakharov 1966) has 27 ribs and similar shell proportions to P. arctica , but not to P. parallela as shown by the data of Woods (1904), and should therefore also be included into P. arctica .
Occurrence. Seep 9 (uppermost Ryazanian), Slottsmøya Member, Svalbard ( Tab. 1 View TABLE 1 ). It also occurs in the Kimmeridgian, Volgian and possibly also the Upper Ryazanian of Siberia ( Zakharov 1966) and the Volgian of Greenland ( Fürsich 1982).
Palaeoecology. Pseudolimea arctica probably took advantage of hard substrates present in the seeps, such as exposed carbonate and other shells, for byssal attachment. Modern limids can be found both in shallow and deep marine environments ( Allen 2004). They are suspension feeders, byssally attached to rocks, shells and marine plants. They can also form ‘nests’ composed of mucus, byssal threads and various incorporated hard elements if the substrate proves unsuitable for byssal attachment ( Merrill & Turner 1963). Some species possess the ability to swim by clapping their valves, and others are able to relocate using their foot and byssus ( Mikkelsen & Bieler 2008). Unlike other monomyarian bivalve families, limids are not pleurothetic, i.e. they do not rest on one of the valves, but orient their commissure perpendicular to the substrate ( Mikkelsen & Bieler 2008). The extinct genus Pseudolimea probably behaved in the same way. No association between limids and symbiotic bacteria has been recorded to date, but Acesta oophaga Järnegren, Schander & Young, 2007 in Järnegren et al. 2007, from hydrocarbon seep environments in the Gulf of Mexico has morphological adaptations for feeding on the eggs of the tubeworm Lamellibrachia luymesi van der Land & Nørrevang, 1975 . It has a folded posterodorsal shell margin that surrounds the tubeworm plume, where eggs are being released from the tube ( Järnegren et al. 2005). No such adaptation has been recorded in any species of Pseudolimea .

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.