
Nipponoluciola, Ballantyne & Kawashima & Jusoh & Suzuki, 2022
|
publication ID |
https://doi.org/ 10.5852/ejt.2022.855.2023 |
|
publication LSID |
lsid:zoobank.org:pub:A31C64CB-6C6D-424A-A54C-F86FDD77ABAB |
|
DOI |
https://doi.org/10.5281/zenodo.7528340 |
|
persistent identifier |
https://treatment.plazi.org/id/A2478793-7742-FFB6-FD34-F964833FF832 |
|
treatment provided by |
Felipe |
|
scientific name |
Nipponoluciola |
| status |
gen. nov. |
Nipponoluciola gen. nov.
urn:lsid:zoobank.org:act:BD75E8CA-BC67-4597-A0A5-6BCEF5BA7B4F
Figs 1‒3 View Fig View Fig View Fig , 5‒12 View Fig View Fig View Fig View Fig View Fig View Fig View Fig View Fig
Type species
Luciola cruciata Motschulsky, 1854 View in CoL , designated here.
Diagnosis
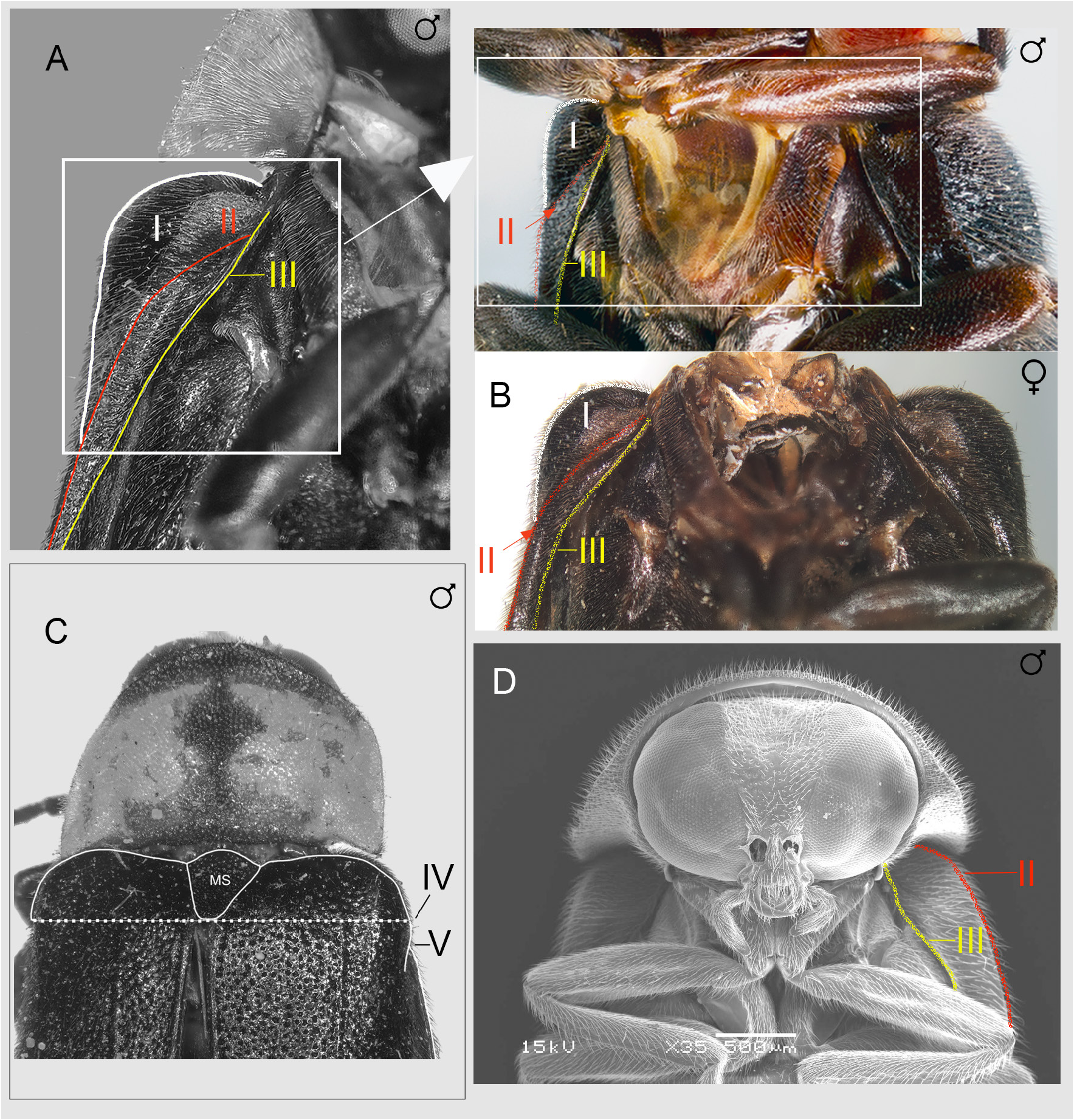
Nipponoluciola gen. nov. is a genus of Japanese Luciolinae fireflies and consists of two species both of which have aquatic larvae with abdominal gills. While Nipponoluciola belongs to that group of 18 genera of Luciolinae where the aedeagus in the male has the LL widely visible at the side of the ML ( Ballantyne et al. 2019), it is one of only two genera in this group of 18 (the other is Missimia ) where the elytral epipleuron at its base is very narrow and when viewed from beneath in a horizontal specimen the humerus is widely visible in both males and females ( Fig. 3A–C View Fig ). In the other genera of Luciolinae with LL visible at the sides of the ML the base of the elytral epipleuron is expanded, and the humerus is not visible from below in a horizontal specimen (example in Fig. 3D View Fig ). It is distinguished from Missimia by distribution ( Missimia is a rare genus known only from four specimens taken in the New Guinean highlands), and the unique feature of Missimia , where the labrum and clypeus are immovably joined ( Fig. 4E View Fig ; Table 1 View Table 1 ; Ballantyne & Lambkin 2009: figs 27, 29–30, 32). The clypeolabral suture in Nipponoluciola is flexible. It is distinguished from the genus Aquatica , with which it shares similar aquatic larvae bearing abdominal gills, by the following features: Aquatica species adult males may have pale margined elytra ( Fig. 4A View Fig ), the elytral epipleuron at its base is wide ( Fig. 3D View Fig ), the LO in V7 is usually retracted from the posterior margin ( Fig. 4B View Fig ), the lateral margins of V7 are not angulated, both left and right margins of the posterior area of the sheath sternite have pointed projections, the aedeagal basal piece is not notched ( Fig. 4C–D View Fig ). Females have thin paired bursa plates ( Fu et al. 2010). See Table 2 View Table 2 .
Males have black elytra and the pronotum may have median dark brownish to blackish markings and an underlying pink fat body (in N. cruciata ) ( Figs 1 View Fig , 2G View Fig , 6 View Fig ), or have no dark markings and underlying yellowish orange fat body (in N. owadai ) ( Figs 1 View Fig , 2H View Fig ). Females are coloured like males, and are larger, macropterous, with spermatophore digesting gland, no obvious bursa plates and a small transverse median oviduct plate in N. cruciata ( Fig. 7 View Fig ). Larvae ( Figs 8–9 View Fig View Fig ) are aquatic, have forked gills along the sides of the abdominal segments 1–8 and apical sense organs on maxillary and labial palpomeres. Distinguished most obviously from Aquatica larvae by the differences in the markings of protergum (outlined subsequently); distinguished from A. lateralis by the unbranched setae on the dorsal surface of the mandibles, and the absence of microspines on pale areas of paired tergites of meso- and metathoraces, and abdominal segments 1–8 ( Okada 1928: figs 2‒3, 2‒4 depicted but did not name these microspines; IK confirmed their presence in A. lateralis ).
Etymology
The generic name seeks to honour the country of origin of these two species and their status as celebrated icons. ‘Nippon’ is the Japanese name for Japan, and Luciola represents the genus to which these species were formerly assigned. Gender: feminine.
Description
Male ( Figs 1–3 View Fig View Fig View Fig , 5–6 View Fig View Fig , 10‒12 View Fig View Fig View Fig )
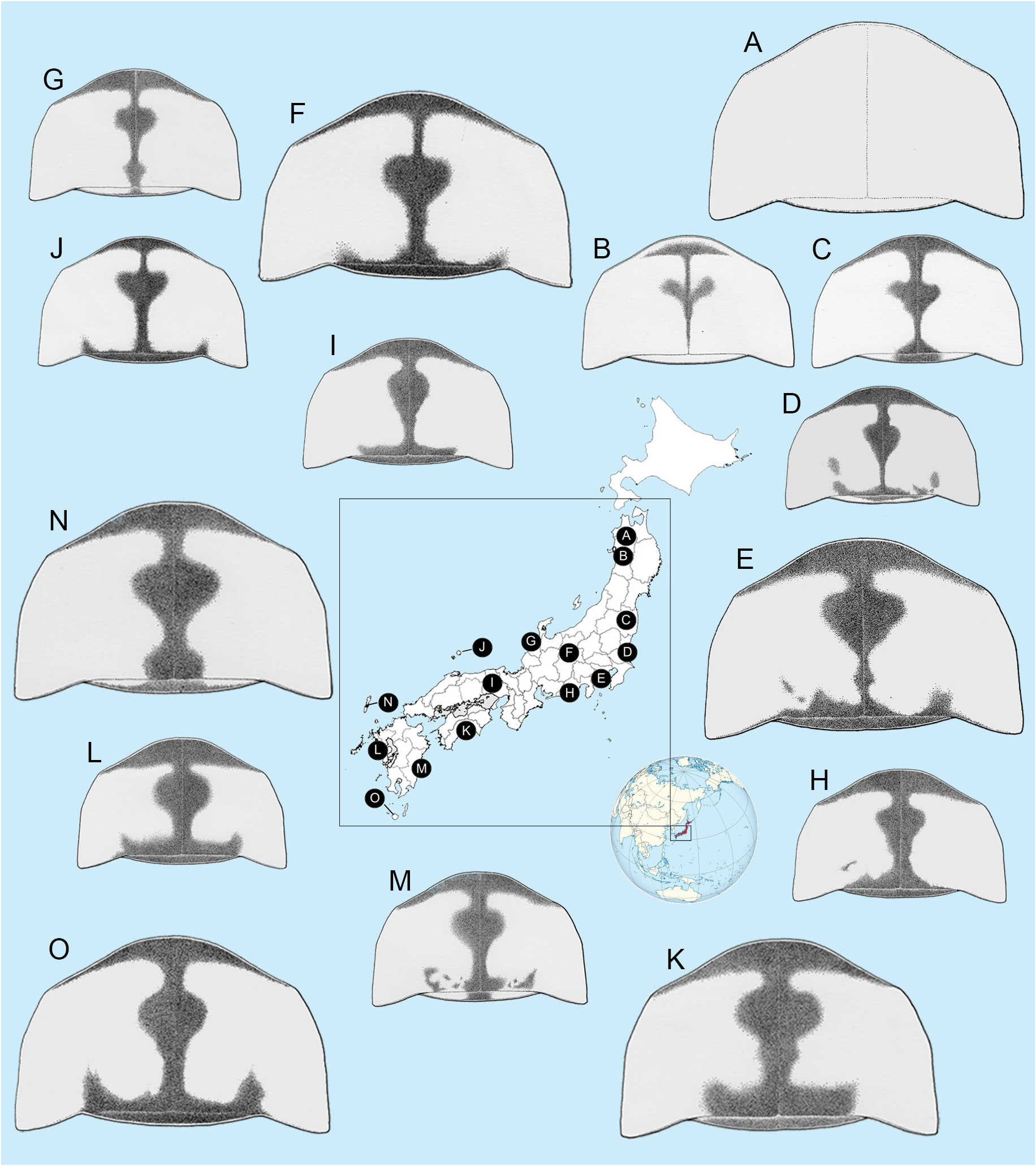
PRONOTUM ( Figs 1–2 View Fig View Fig ). Punctation dense; anterior margin broadly rounded and projecting beyond rounded obtuse anterolateral corners, slightly so in owadai ; lateral margins slightly divergent, the pronotum being wider across its posterior area than elsewhere (C>A, B), width (C) subequal to humeral width; posterolateral corners narrowed, apically rounded, slightly less than 90°, projecting posteriorly slightly beyond median posterior margin, which is not emarginated, and separated from it by welldefined emarginations. Pronotal colour variations depicted in Fig. 6 View Fig are discussed under Nipponoluciola cruciata gen. et comb. nov.
HYPOMERA. Not well flattened; pronotal width/GHW 1.43–1.47.
ELYTRON ( Figs 1–3 View Fig View Fig View Fig ). Punctation dense, not linear, not as large as that of pronotum, nor widely and evenly spaced; apices not deflexed, nor emarginated; epipleuron and sutural ridge extend beyond mid-point, almost to apex but not extending around apex, neither thickened in apical half; with three interstitial lines of which the inner two interstitial lines are well defined; in horizontal specimen viewed from below epipleuron at elytral base narrow with humerus visible; viewed from above in horizontal specimen anterior margin of epipleuron arises well behind the posterior margin of MS ( Fig. 3A, C View Fig ); epipleuron developed as a lateral ridge along most of length; sutural margins approximate along most of length in closed elytra; lateral margins parallel-sided.
HEAD ( Figs 1–2 View Fig View Fig ). Gently depressed between eyes; at least anterior portion exposed in front of pronotum, not capable of complete retraction within prothoracic cavity at rest; eyes moderately separated beneath at level of posterior margin of mouthpart complex; frons-vertex junction not defined this area rounded, posterolateral eye excavation not developed; antennal sockets on head between eyes, close but not contiguous, ASD subequal to ASW; clypeolabral suture present, flexible, not in front of anterior eye margin when head viewed with labrum horizontal; outer edges of labrum reach inner edges of closed mandibles. Mouthparts: functional; apical labial palpomere flattened, shaped like a broad triangle, longer than wide, with inner edge dentate. Antennae ( Fig. 1 View Fig ) 11 segmented; approximately 2× GHW; FS simple elongate; GHW/SIW 3.70–4.50.
LEGS ( Figs 1–2 View Fig View Fig ). Without MFC; no legs with curved or expanded femora or tibiae; no basitarsi expanded or excavated.
ABDOMEN ( Fig. 2F View Fig ). No ventrites with curved posterior margins nor extending anteriorly into emarginated posterior margin of anterior segment. LO in V7 entire, LO present in V6, occupying almost all V6 ( Jeng et al. 2003: figs 7, 9). V7 with elongate apically rounded MPP and angulate lateral areas which are not prolonged and correspond to where PLP would originate in other species. T7 without prolonged anterolateral corners; T8 not heavily sclerotised; no concealed anterolateral arms of T8 and the anterolateral corners scarcely produced (see Jeng et al. 2003: figs 21–22); lateral margins subparallel sided; posterior margin evenly rounded and entire; T8 ventral surface smooth even without any ridges or projections.
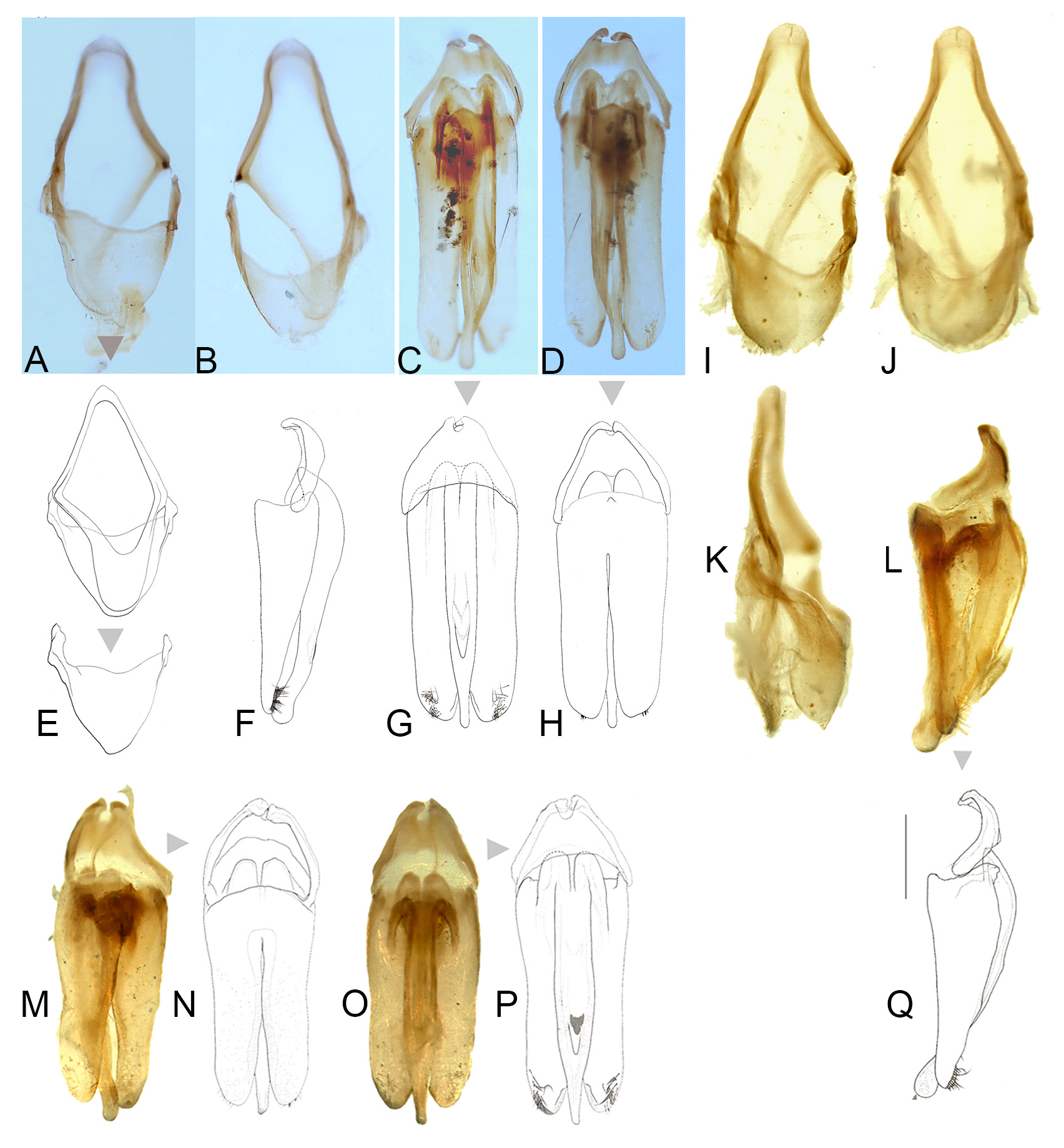
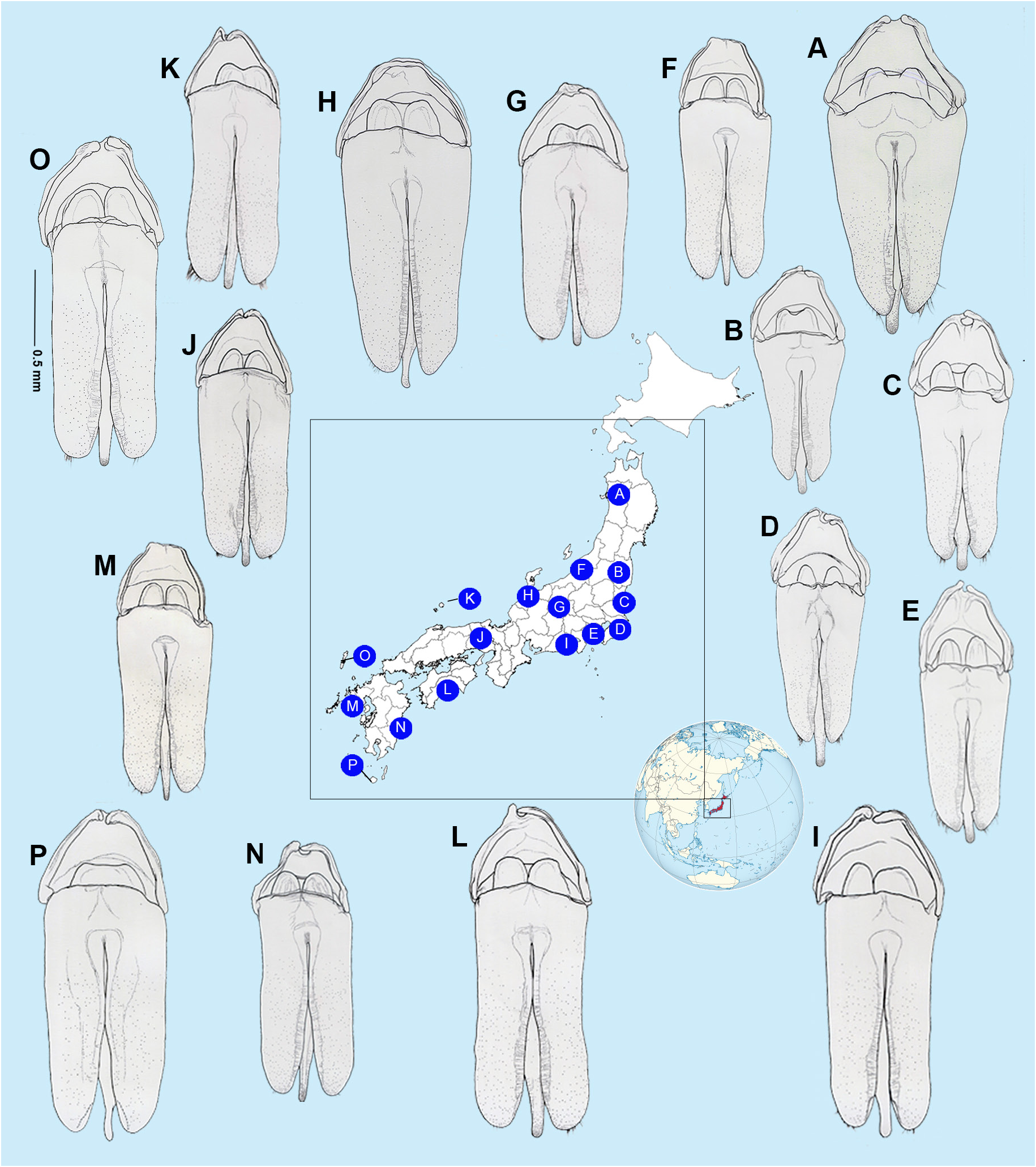
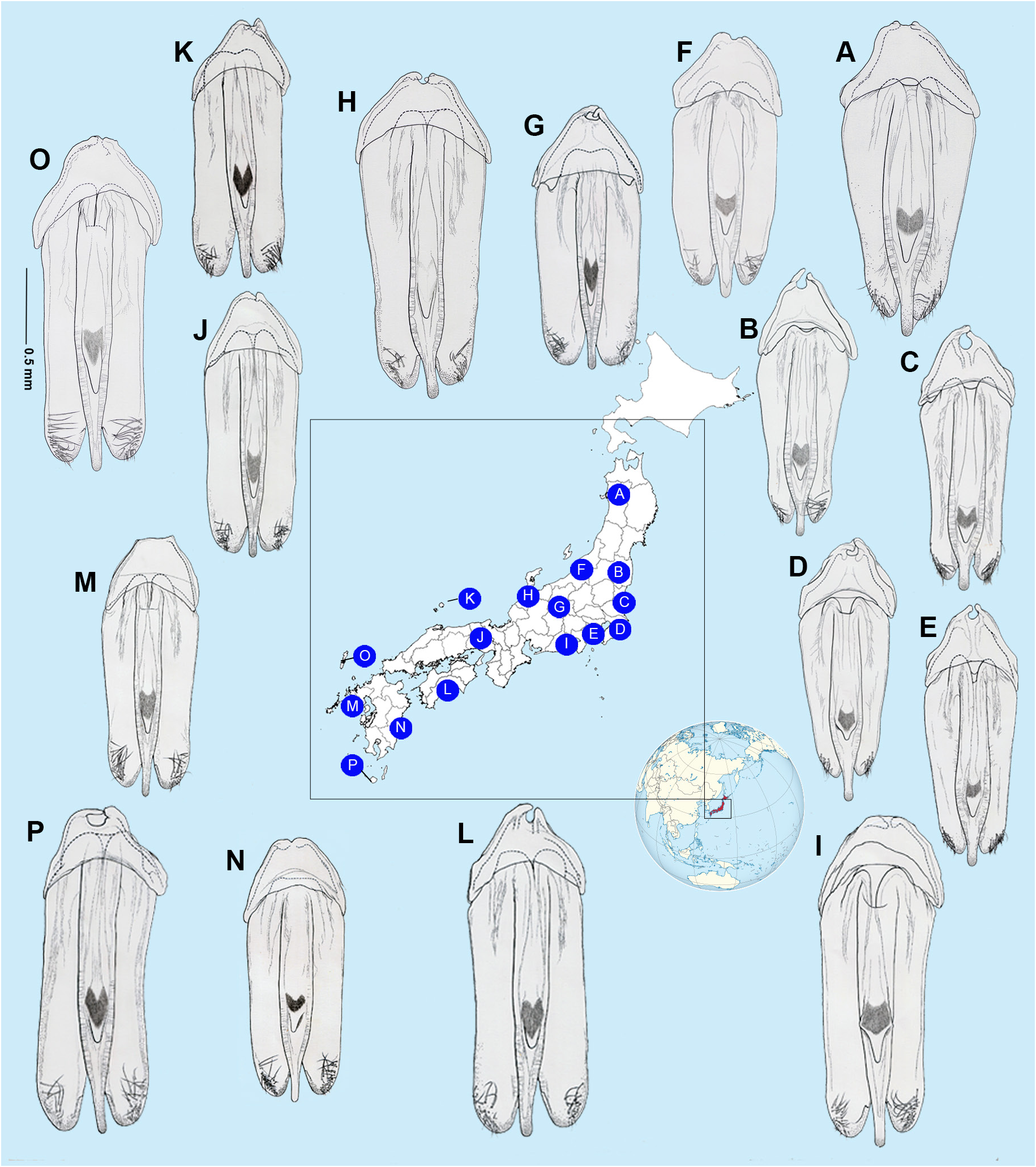
AEDEAGAL SHEATH ( Fig. 5 View Fig ). Approx. 1.94–2.12 times as long as wide; without bulbous paraprocts; symmetrical in anterior half where sheath sternite tapers evenly to a broadly rounded apex; lateral arms of sheath tergite widely visible from below on left, narrowly on right; posterior area of sternite (posterior to the articulation of the tergite arms) narrow, emarginated on both sides (depending on orientation it may appear to be more deeply emarginated on right side) with margin even, without projections, and sternite apex rounded; tergite broad with anterior margin slightly and evenly emarginated; no obvious subdivision into more than one segment observed; posterior margin entire and evenly rounded; tergite projecting about as far as apex of sternite.
AEDEAGUS ( Figs 5 View Fig , 10–12 View Fig View Fig View Fig ). Subparallel sided; basal piece well defined and usually with distinctive, asymmetrical median notch; LL widely visible beside ML; LL of similar length, slightly shorter than ML; width across LL, (measured from beneath), much wider than width across widest part of median lobe (3‒3.3); ML slender gradually narrowing in apical 1/7; apices of LL with ventral surfaces broad, slightly curved and hairy. Differences in aedeagal patterns in Nipponoluciola cruciata gen. et comb. nov. are addressed under that species in Figs 10‒12 View Fig View Fig View Fig .
Female ( Fig. 7 View Fig )
Macropterous and flight capable.
BODY LENGTH. 14.0– 18.4 mm ( Jeng et al. (2003) listed a range of 15.0– 18.6 mm long).
COLOUR ( Fig. 7A, C–D View Fig ). As for male except white LO restricted to V6, pinkish fat bodies beneath V 7 in cruciata , or V7 completely black and underlying fat body not visible in owadai ; black V8 with paired semitransparent areas; all tergites black except for T7 which has a median dark band and semitransparent lateral areas under which pink fat bodies are visible in cruciata .
PRONOTUM ( Fig. 7 View Fig ). Outline as for male; pronotal width/GHW 1.88–2.20.
HEAD. GHW/SIW 2.67–2.90.
ABDOMEN ( Fig. 7C–D, I View Fig ). Lateral margins of V8 taper posteriorly, V8 with shallow median posterior emargination; anterior apodeme of V8 well sclerotised and separated from posterior area of V8 by clear transparent cuticle ( Fig. 7I View Fig ).
REPRODUCTIVE SYSTEM ( Fig. 7E‒H, J View Fig ). Spermatophore digesting gland well defined usually with particulate remnants inside; spermatheca with expanded non sclerotised base; no obvious bursa plates; median oviduct plate well defined ( Fig. 7G–H View Fig ), anterior margin curved and irregular, lateral margins straight and converging slightly posteriorly, posterior margin slightly and broadly emarginated; lying transversely in the median oviduct and curved possibly to accommodate to the outline of the duct. Ovipositor: elongate slender ( Fig. 7J View Fig ).
Larva ( Figs 8A–B View Fig , 9A‒H View Fig ) (see also Fu et al. 2012b: figs 81–82)
DORSAL SURFACE ( Figs 8–9 View Fig View Fig ). With 3 thoracic and 9 obvious abdominal segments of which abdominal segments 1–8 bear lateral tracheal gills which are forked and compound, with a non-functional spiracle located at the end of the shorter branch. A narrow ring of cuticle at the posterior end of segment 9 has been attributed to segment 10 ( Ballantyne & Menayah 2002: figs 2–3; Nada et al. 2021) and may be completely covered and concealed by the posterior margin of segment 9 from above in aquatic species, but visible from beneath. Lawrence & Ślipiński (2013) consider that segment 10 has been transformed into the holdfast organ (pygopodia) at the end of the abdomen. We refer to abdominal terga 1‒9 only as these are clearly visible. Elongate, tapering at front and behind, membranous and very soft bodied, and somewhat flattened. The thorax and abdomen are almost membranous, with ventral sclerites degenerate and extent barely demarcated by wrinkles or folds, except for a single coloured dorsal plate on both the prothorax and abdominal segment 9, and paired well sclerotised and coloured tergites on the meso and metathorax, and abdominal segments 1–8. A wide membranous area/line extends from the anterior margin of the mesothorax to the posterior margin of abdominal segment 8. Without laterally explanate tergal margins thus laterotergites and gills visible from above.
VENTRAL SURFACE ( Fig. 9B, D, F View Fig ). The extent of the following areas in very soft bodied aquatic larvae is often difficult to determine. A lateral pleural suture delimits laterotergites above in the thorax and abdomen (e.g., Ballantyne & Menayah 2002: fig. 1; Fu et al. 2012b: figs 81–82). In the thorax the median ventral surface of both meso and metathorax is subdivided into two areas by a sternocostal suture running anterior to the coxae, an anterior basisternum with laterotergites at the sides in the mesothorax; (in terrestrial larvae these mesothoracic laterotergites bear functional spiracles, but spiracles were not clearly visible in the larvae examined); a posterior median subrectangular sternellum bears the legs and is margined laterally by laterotergites; the episterna and epimera of both the meso and metathorax are visible above the coxae of the meso and metathoracic legs as thin dark sclerotised plates (see interpretation in Fu et al. 2012b: 5). This subdivision does not extend to the dorsal surface. In the abdominal segments 1–8 a median subrectangular area is attributed to the sternum; this is margined laterally by elongate narrow paired laterosternites, which are delimited by folds from the median sternal plate below, and the laterotergites above; in this aquatic genus the absence of any additional colour on the ventral surface makes interpretation of these areas difficult, and they may be incompletely distinguished only by folds in the membrane; laterotergites of segments 1–8 bear branched gills. Eversible branched defensive organs arise at the sides of the meso and metathorax, and abdominal segments 1–8, in the membrane at the sides of the terga of those segments and above the laterotergites of the abdomen (see below).
HEAD ( Fig. 8C View Fig ) ( Kanda 1935: fig. 13; Hara 1962: figs 1, 5; Fu et al. 2012b: figs 25–30, 69). Subparallel sided, dorsoventrally flattened, prognathous, well sclerotised, with an ocellus at each side; not visible when retracted into the prothoracic cavity; extensible neck membrane forming two layered envelope around retracted head; capable of considerable extension beyond the anterior protergal margin; head capsule divisible into median dorsal frontoclypeus, bounded laterally by the U shaped frontal arms of the ecdysial line (= epicranial suture); lateral parietal plates at the sides of the frontoclypeus are separated behind by the epicranial stem and are reflexed ventrally but not meeting; maxillae and labium fused forming a maxillolabial complex covering most of ventral head area.
ANTENNAE ( Fu et al. 2012b: figs 27–29). Slender, cylindrical, 3 segmented, with elongated scape and pedicel, and apical very short (3 rd) segment (the flagellum), with apical sensilla, subequal in length to adjacent sense cone; elongate ‘articulating membrane’ forms two layered envelope around retracted antenna.
MOUTHPARTS ( Fu et al. 2012b: figs 25, 29, 69). Well developed; mandibles falcate, strongly sclerotised; densely covered in fine hair along outer margins; densely pubescent along basal half of inner margins; perforated along length by a fine canal that opens on the outer margin just behind the apex; without a basal retinaculum; fine reticulate sculpture on the dorsal surface; with a row of elongate curved setae arising in a line just inside the outer margin. Maxillae with short, squat, four segmented palp bearing sense organs at the apex of segment 4; basal segment (which may be interpreted as the palpifer) large and well defined, segments 2 and 3 very short and diminishing in width towards apex, apical segment longer and narrower than palpomere 3, with apical sense organs; palp may obscure galea, which is long, thin, two segmented, bearing long and short setae at apex, and on its inner margin an elongate, flattened, dense profusion of anteromedially directed hairs (lacinia); cardo well defined, articulating with broad elongate stipes; fused along its outer edges with the reflexed head margins, and along its inner margins with the median labium. Labium with two segmented labial palpi, bearing sense organs at the apex of segment 2, and arising at anterolateral corners of small prementum which lacks a ligula; apical palpomere elongate conical with two or three peg-like sensilla on the sides; anterior area of prementum densely covered by minute spines on both sides of dorsal surface; postmentum elongate, not well sclerotised and colourless, and joined along sides by membrane to the cardines.
THORAX ( Figs 8–9 View Fig View Fig ) ( Fu et al. 2012b: figs 81–82). Prothorax longer than wide, and containing retracted head within; with ventral surface little differentiated. Protergum elongate or elliptical, with lateral margins arcuate and shallowly expanded laterad, divergent posteriorly; anterior and posterior margins rounded and widely arcuate, connecting directly to lateral margins without forming corners. Each tergite of meso- and metathorax as are those of abdominal segments 1–8 separated from each other as a pair of plates, subequal in size; the shape more or less round pentagonal; any corners more or less rounded; the margins lightly pale margined, broadened at the lateral margins to form pale spots; lateral margins arcuate, expanded to laterad; hind margins also arcuate; inner margins straight and parallel-sided to each other. Legs are four segmented – short cylindrical coxae with bases widely separated; elongate trochanters joining femora obliquely; tibiotarsus terminated by a single claw. In contrast with previous treatments the leg segments are attributed to coxa, trochanter, femur and tibiotarsus.
ABDOMEN ( Figs 8–9 View Fig View Fig ) ( Fu et al. 2012b: figs 81–82). Each tergite of abdominal segments 1–8, as are those of the meso- and metathorax, is separated as a pair of plates, similar form to those of meso- and metathoracic terga, more or less round pentagonal, diminishing slightly in both length and width toward posterior ones, not forming any corners; the margins lightly pale margined, broadened at the lateral margins to form pale spots; segment 9 has a single round tergal plate, with the margin also lightly pale margined as in segments 1–8; pale posterior margin often widens slightly to form a narrow pale spot; abdominal tergal width decreasing a little towards posterior end, ventral surface of all but terminal abdominal segment (segment 9) with paired laterotergites bearing gills at the sides (visible along sides of body as lateral projections); median sternal element in each of abdominal segments 1–8 margined by elongate slender laterosternites but these areas difficult to discern in soft bodied larvae; (laterosternites appear membranous on segment 8 which houses the light organs).
DEFENSIVE ORGANS. With 10 pairs of similar pale white, forked eversible defensive organs ( Okada 1928: fig. 1a; Kanda 1935: 57–58, pl. 5-5; Trice et al. 2004: 2; Fu et al. 2009: fig. 4; 2012b: figs 7–12) located laterally on the meso- and metathorax, and above the tracheal gills on each of the 8 abdominal segments; these organs arise in the thorax in folds of membrane on the upper side of the laterotergites and just below the lateral margins of the tergal plates, and in the abdomen above the abdominal laterotergites and below the lateral margins of the tergal plates; organs have protuberances on their external surface, each composed of a well-developed globular body attached to a long thick stalk; globular bodies consist of secretory cells.
PYGOPODIA ( Fig. 8 View Fig , partly visible in Fig. 9A View Fig ). On the external surfaces of the tubular larval eversible holdfast organs, numerous hook-shaped protuberances grow in circular rows on each tube.
List of species of Nipponoluciola gen. nov.
Nipponoluciola cruciata ( Motschulsky, 1854) gen. et comb. nov.
Nipponoluciola owadai ( Satô & Kimura, 1994) gen. et comb. nov.
Key to species of Nipponoluciola gen. nov. using males
1. Widely distributed in Japan; pronotum usually with pinkish fat body visible through cuticle, and variable median blackish marking which may be expanded in the median area to assume a cross like formation; MS and elytra black ( Figs 1–2 View Fig View Fig ) ......................................................................................... ......................... N. cruciata ( Motschulsky, 1854) (Honshu, Shikoku, Kyushu and their sub-islands)
– A rare and protected species known only from the Okinawa Islands group; pronotum yellowish orange, without median dark markings, and fat body without pinkish colour; MS yellowish orange and elytra black ( Figs 1–2 View Fig View Fig ) ................................................................................................................. ....... N. owadai ( Satô & Kimura, 1994) (only Kume-jima Is. of the Okinawa Isls, Middle Ryukyus)







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.