
Sylvicanthon proseni (Martínez, 1949)
|
publication ID |
https://doi.org/ 10.5852/ejt.2018.467 |
|
publication LSID |
lsid:zoobank.org:pub:8D27AAB8-B7F2-424C-B1A6-66FEFA66EDFF |
|
DOI |
https://doi.org/10.5281/zenodo.3846303 |
|
persistent identifier |
https://treatment.plazi.org/id/A72C87FB-FFCB-FFD7-0E97-0DA90893928F |
|
treatment provided by |
Valdenar |
|
scientific name |
Sylvicanthon proseni (Martínez, 1949) |
| status |
|
Sylvicanthon proseni (Martínez, 1949) View in CoL revalidation and comb. nov.
Figs 5A View Fig , 6D View Fig , 8B View Fig , 12A, 12C View Fig , 14A, 14C View Fig , 15 View Fig G–H, 17F, 20, 28C–D, 29C–E, 30, 31B
Glaphyrocanthon proseni Martínez, 1949a: 282 View in CoL , 287–291, fig. 2.
Glaphyrocanthon proseni View in CoL – Halffter & Matthews 1966: 43, 293. — Halffter & Martínez 1977: 90–91 (as synonym of C. aequinoctialis View in CoL ). — Solís & Kohlmann 2002 (as synonym of C. aequinoctialis View in CoL ).
Glaphyrocanthon (Glaphyrocanthon) proseni View in CoL – Martínez 1950: 170–171. — Pereira & Martínez 1956: 92, 126, 128. — Martínez et al. 1964: 3, 5, 8, 10–11, 13. — Vulcano & Pereira 1964: 663; 1967: 561.
Canthon (Canthon) proseni – Vaz-de-Mello et al. 2011b: 82, 86, 93. — Chamorro et al. 2018: 87, 92, fig. 10D, F.
Canthon proseni – Jacobs et al. 2008: 642. — Korasaki et al. 2012: 425, 427. — Silva et al. 2014: 348. — Nunes et al. 2014: 410–411. — Silva et al. 2017: 490.
Canthon aequinoctialis View in CoL sensu – Balthasar 1941: 341; 1951: 326. — Escobar 2000a: 206; 2000b: 114, 121 (mixed information with S. aequinoctialis View in CoL comb. nov.). — Medina et al. 2001: 135 (idem). — Celi et al. 2004: 42. — Noriega-Alvarado 2004: 40. — Noriega et al. 2008: 78–79. — Carpio et al. 2009: 462, 464, 469. — Medina & Pulido 2009: 58. — Figueroa & Alvarado 2011: 210–211, fig. 1c. — Noriega 2012: 4. — Ratcliffe et al. 2015: 195.
Canthon aequinoctiale sensu – Vaz-de-Mello 1999: 449–450.
Canthon (Canthon) aequinoctiale sensu – Vaz-de-Mello 2000.
Canthon (Canthon) aequinoctialis View in CoL sensu – Noriega et al. 2007a: 54–55.
Canthon (Canthon) aequinotialis [sic] – Culot et al. 2011: Supporting information, table S1.
Etymology
Eponym after the Argentinian entomologist Alberto F. Prosen (1902–1972).
Material examined
Holotype
BOLIVIA: ♂, La Paz, Nor Yungas, confluence between the Choro and Coroico Rivers, 700 m (Martínez 1949), (“ BOLIVIA / Dep. La Paz / Prov. Nor Yungas / Ríos Coroico y Choro / 700 mts. alt. / Coll. Martínez / Ene.-949 ”, “ HOLOTYPUS ”, “ Glaphyrocanthon / proseni ♂ / sp. nov. / A. Martínez det. 19 49 ”, “MACN-En / 1412”, “ FICHADO ”), genital capsule removed and glued on a square label ( MACN) ( Fig. 29C View Fig )
Paratypes
BOLIVIA: 1 ♂, (“ BOLIVIA / Yungas de Palmar / 1000 M. – Zischka”, “H. &A. Howden / COLLECTION / ex.A. Martinez coll.”, “PARATIPO”, “ Glaphyrocanthon / proseni / sp. n. / Ƌ / A. Martinez – det. 19 49 ”) ( CMNC) (labels examined by photograph, Fig. 29D View Fig ); 1 ♂, (“Prov. del Sara / Cent. Bolivia / 450 m. 1909 26 ” / J. Steinbach”, “H. & A. Howden / COLLECTION / ex. A. Martinez coll.”, “PARATIPO”, “ Glaphyrocanthon / proseni / sp. n. / Ƌ / A. Martinez – det. 19 49 ”) ( CMNC) (labels examined by photograph, Fig. 29D View Fig ).
Additional material (495 ♂♂, 328 ♀♀, 31 unsexed specimens)
BOLIVIA: Cochabamba: 3 ♂♂, 1 ♀, José Carrasco, Valle del Sacta, 17º04′18″ S, 64º46′00″ W, 250 m, 21–22 Dec. 2004, pitfall with human faeces, Mann, Hamel and Simmons leg. ( OUMNH); 2 ♀♀, Chapare, Mar. 1994 ( CEMT).
BRAZIL: 1 unsexed specimen, “Amazon”, H.W. Bates leg. ( BMNH); 1 ♂, Rio Purus, Ehrenreich leg. ( ZMHB); 1 ♂, Rio Xingu, Ponte Nova, Feb. (no year) ( CEMT). – Acre: 14 ♂♂, 20 ♀♀, Mâncio Lima, 07º28.584′ S, 72º54.110′ W, 5 Dec. 2012, H.M.B. Luiz and N.S.G.F. Adem leg. ( CEMT); 40 ♂♂, 15 ♀♀, Manoel Urbano, Parque Estadual Chandless, 09º22′26″ S, 69º55′20″ W, 24 Jun. 2013, T.F. Brito leg. ( CEMT); 41 ♂♂, 14 ♀♀, Manoel Urbano, Parque Estadual Chandless, 09º22′36″ S, 69º55′20″ W, 1 Jul. 2013, T.F. Brito leg ( CEMT); 1 ♀, Rio Branco, Jul. 1996, M. Castro leg. ( CEMT); 1 ♂, 1 ♀, Senador Guiomard, 10º04′ S, 67º37′ W, 14 Apr. 2017, human faeces, Bruna S. Bitencourt leg. ( CEMT); 1 ♂, 1 ♀, Senador Guiomard (“Rio Branco”), Fazenda [Experimental] Catuaba, Feb. 1997 , F.Z. Vaz-de-Mello leg. ( CEMT); 1 ♀, Xapuri, Reserva Chico Mendes, 10º20.975′ S, 68º40.840′ W, 17 Oct. 2008, pitfall with human faeces, J. Silveira leg. ( CEMT); 1 ♂,Xapuri, Reserva Chico Mendes, 10º18.930′ S, 68º40.756′ W, 18 Oct. 2008, pitfall with human faeces, J. Silveira leg. ( CEMT). – Amazonas: 1 ♂, without specific locality, 4 Jun. 1939 ( MZSP); 1 ♂, 1 ♀, Benjamin Constant, Mar.–Apr. 1942, Parko leg. ( MNRJ); 1 ♂, same collecting data as for preceding ( MZSP); 1 ♀, Benjamin Constant, Dec. 1979,A. Lise leg. ( MCNZ); 1 ♂, Benjamin Constant, Rio Javari, Dec. 1960, Dirings leg. ( MZSP); 1 ♂, 1 ♀, Beruri, BR-319, Km 350, 05º12′56,4″ S, 61º50′22,6″ W, 30 Jul.–2 Aug. 2007, H. Gasca leg. ( CEMT); 1 ♂, Manaus, Jan. 1937 ( MZSP); Manaus, II.1941, Parko leg. 1 ♀ ( CEMT); 1 ♀, Manaus, Nov. 1941 ( MZSP); 1 ♂, Manaus, Rio Negro, Dec. 1941, B. Pohl leg. ( MNRJ); 1 ♂, 1 ♀, same collecting data as for preceding ( MZSP); 1 ♂, Manicoré, Rio Madeira, Jun. (no year), J.F. Zikán leg. ( FIOC); 1 ♂, São Paulo de Olivença (“Olivenza”) ( ZMHB); 1 ♀, São Paulo de Olivença, Nov. (no year), J.F. Zikán leg. ( MZSP); 1 ♂, São Paulo de Olivença (“Olivenza”), O. Thieme leg. ( ZMHB); 1 unsexed specimen, Tefé (Ega), H.W. Bates leg. ( BMNH); 1 ♀, Tefé, 1921, J.F. Zikán leg ( FIOC). – Maranhão: 1 ♀, Centro Novo do Maranhão, Igarapé Gurupi-Uma, Aldeia Araçu, 50 km E of Canindé, May 1963, Malkin leg. ( MZSP). – Mato Grosso: 2 ♂♂, 1 ♀, Alta Floresta, 09º52′52″ S, 56º06′12″ W, Mar. 2008, pitfall with human faeces, E. Berenguer leg. ( CEMT); 18 ♂♂, 18 ♀♀, Alta Floresta, 09º50′22″ S, 56º00′21″ W, pitfall with human faeces, Apr. 2008, E. Berenguer leg. ( CEMT); 31 ♂♂, 29 ♀♀, Alta Floresta, 09º47′56″ S, 55º55′33″ W, May 2008, pitfall with human faeces, E. Berenguer leg. ( CEMT); 11 ♂♂, 8 ♀♀, Alta Floresta, 09º53′40″ S, 56º16′35″ W, Jun. 2008, pitfall with human faeces, E. Berenguer leg. ( CEMT); 5 ♂♂, 8 ♀♀, Alta Floresta, CEPLAC, 6 Feb. 2009, pitfall, V. Gonçalves leg. ( CEMT); 3 ♂♂, 3 ♀♀, Alta Floresta, CEPLAC, 24 May 2009, V. Gonçalves leg. ( CEMT); 1 ♀, Alta Floresta, CEPLAC, 4 Dec. 2009, pitfall, V. Gonçalves leg. ( CEMT); 1 ♂, Alta Floresta, CEPLAC, 1 May 2010, human faeces, V. Gonçalves leg. ( CEMT); 2 ♀♀, Alta Floresta, CEPLAC, 9 Jan. 2010, pitfall, V. Gonçalves leg. ( CEMT); 1 ♀,Carlinda, 13 May 2008, C.R. Menezes leg. ( CEMT); 1 ♂, Carlinda, 13 May 2008, J. Ortega leg. ( CEMT); 1 ♂, Carlinda, 13 May 2008, C. Rosseto leg. ( CEMT); 1 ♂, 1 ♀, Carlinda, 14 May 2008, M.E. Ortiz leg. ( CEMT); 1 ♂, 14 May 2008, J.E. Silva leg. ( CEMT); 1 ♂, Carlinda, 15 May 2008, M. Francisco leg. ( CEMT); 1 ♂, Carlinda 16 May 2008, M.L. Santos leg. ( CEMT); 1 ♀, Carlinda, 14 Oct. 2008, E. Macena leg. ( CEMT); 1 ♀, Carlinda, 21 Oct. 2008, B.C. Kain leg. ( CEMT); 2 ♂♂, Carlinda, 22 Oct. 2008, L. Sabino leg. ( CEMT); 1 ♂, Carlinda, 23 Oct. 2008, E.P.P. Cruz leg. ( CEMT); 1 ♂ (dissected), 1 ♀, Carlinda, 23 Oct. 2008, K. Segatto leg. ( CEMT); 1 ♀, Carlinda, 10 Oct. 2009, E. Teles leg. ( CEMT); 1 ♀, Cotriguaçu, May 2011, R.E. Vicente leg. ( CEMT); 4 ♂♂, Cotriguaçu, Fazenda São Nicolau, 09º50′24″ S, 58º15′10″ W, 250 m, 6 Oct. 2009, human faeces, Vaz-de-Mello leg. ( CEMT); 9 ♂♂, 1 ♀, Cotriguaçu, Fazenda São Nicolau, 09º50′24″ S, 58º15′10″ W, 250 m, 8 Oct. 2009, human faeces, Vazde-Mello leg. ( CEMT); 3 ♂♂, Cotriguaçu, Fazenda São Nicolau, 09º50′24″ S, 58º15′10″ W, 250 m, 12 Oct. 2009, flight interception trap, Vaz-de-Mello leg. ( CEMT); 1 ♂, Cotriguaçu, Fazenda São Nicolau, 10 Dec. 2009, black light, M.R. Barreto leg. ( CEMT); 3 ♂♂, 3 ♀♀, Cotriguaçu, Fazenda São Nicolau, 09º50′53″ S, 58º14′36″ W, 11–13 Dec. 2009, human faeces, F.Z. Vaz-de-Mello leg. ( CEMT); 1 ♂, 1 ♀, Cotriguaçu, Fazenda São Nicolau, Castanheira, 09º49′18″ S, 58º17′18″ W, 12–14 Dec. 2009, pitfall with human faeces, F.Z. Vaz-de-Mello leg. ( CEMT); 1 ♂, 1 ♀, Cotriguaçu, Fazenda São Nicolau, Mata do Juruena, 09º52′05″ S, 58º13′35″ W, 202 m, 15 Dec. 2009, pitfall, F.Z. Vaz-de-Mello leg. ( CEMT); 28 ♂♂, 4 ♀♀, Cotriguaçu, Fazenda São Nicolau, Matinha, 09º50′19″ S, 58º15′03″ W, 8 Oct. 2009, pitfall with human faeces, F.Z. Vaz-de-Mello leg. ( CEMT); 2 ♂♂, 3 ♀♀, Cotriguaçu, Fazenda São Nicolau, Matinha, forest edge, 09º50′19″ S, 58º15′03″ W, 13 Dec. 2009, pitfall, Vaz-de-Mello leg. ( CEMT); 2 ♂♂, 1 ♀, Cotriguaçu, Fazenda São Nicolau, Matinha, forest centre, 09º50′19″ S, 58º15′03″ W, 13 Dec. 2009, pitfall, Vaz-de-Mello leg. ( CEMT); 5 ♂♂, 2 ♀♀, Cotriguaçu, Fazenda São Nicolau, Prainha, 09º51′36″ S, 58º12′53″ W, Oct. 2009, pitfall, F.Z. Vaz-de-Mello leg. ( CEMT); 6 ♂♂, 6 ♀♀, Cotriguaçu, Fazenda São Nicolau, Prainha, 09º51′36″ S, 58º12′53″ W, 11–13 Dec. 2009, pitfall with human faeces, F.Z. Vaz-de-Mello leg. ( CEMT); 2 ♂♂, Cotriguaçu, banks of Juruena river, 09º52′47.67″ S, 58º14′07.09″ W, Oct. 2010, pitfall with human faeces, R.V. Nunes and M. Gigliotti leg. ( CEMT); 1 ♂, 1 ♀, Nova Bandeirante, banks of Juruena river, 09º52′47″ S, 58º14′07″ W, Oct. 2010, pitfall with dung, R.V. Nunes and M. Gigliotti leg. ( CEMT); 1 ♀, Novo Mundo, Parque Estadual do Cristalino, May 2013, manual collection, V.S. Corrêa leg. ( CEMT). – Pará: 1 ♂, Almeirim, Monte Dourado, Jan. 1993 ( CEMT); 1 ♂, Alter do Chão, RESEX Tapajós /Arapiuns, 22 Dec. 2008, flight interception trap ( CEMT); 10 ♂♂, 6 ♀♀, Belém, IPEAN, Oct. 1984 [FIT], N. Degallier leg. ( CEMT); 1 ♂, Belém, IPEAN, Nov. 1984, N. Degallier leg. ( CEMT); 3 ♂♂, 1 ♀, Belterra, 15 Jul. 2016, pitfall with human and pig dung, F. França leg. ( CEMT); 1 ♂, Estrada Belém-Brasília, Km 93, Oct. 1959, E. Lobato leg. ( FIOC); 2 ♀♀, Novo Progresso, Fazenda Florentino, 07º08′51″ S, 55º23′38″ W, 230 m, Jan. 2011, pitfall with human faeces, Pelissari leg. ( CEMT); 6 ♂♂, 3 ♀♀ (1 dissected), Novo Progresso, Fazenda Florentino, 06º53′18.18″ S, 55º30′26.25″ W, 2 Jan. 2011, pitfall with human faeces, T.D. Pelissari leg. ( CEMT); 1 ♀, Novo Progresso, Fazenda Florentino, 07º08′00″ S, 55º26.6′00″ W, 230 m, 12 Mar. 2011, pitfall with cow dung, Pelissari leg. ( CEMT); 3 ♂♂, 3 ♀♀, Paragominas, Jan.– Jul. 2011, Solar et al. leg. ( CEMT); 1 ♂, Redenção, Pinkaiti-Aik, 07º46′ S, 51º58′ W, Jun. 1999, P.Y. Scheffler leg. ( CEMT); 1 ♂, Redenção, Pinkaiti-Aik, 07º46′ S, 51º58′ W, Nov. 1999, P.Y. Scheffler leg. ( CEMT); 1 ♂, 1 ♀, Santarém, Mar. 1923, Boy leg. ( FIOC); 1 ♀, Santarém Jun. 1924, H.C. Boy leg. ( MNRJ); 3 ♂♂, 2 ♀♀, Santarém, May 2010, V.H. Oliveira leg. ( CEMT); 1 ♂, 1 ♀, Santarém, 03º00′01″ S, 54º52′38″ W, 300 m, 8 Jun. 2010, pitfall with dung, Victor H.F. Oliveira leg. ( UFPA); 1 ♂, Santarém, Reserva Tapajós , 2º36.662′ S, 55º36.513′ W, 7 Jan. 2009, pitfall with human faeces, R. Andrade leg. ( CEMT); 1 ♂, São Félix do Xingu, Pinkaití Reserve, 07º45′ S, 51º57′ W, 31 Oct. 1998, P.Y. Scheffler leg. ( CEMT); 2 ♂♂, 2 ♀♀, Tailândia, “Empresa Agropalma”, 9–15 Jul. 2016, F. Silva leg. ( CEMT); 3 unsexed specimens, Tapajós, H.W. Bates leg. ( BMNH). – Rondônia: 1 ♂, Cacaulândia (“Caucalandia” sic), 10º32′ S, 62º48′ W, 160–350 m, 10–11 Oct. 1991, John R. Macdonald leg. ( CEMT); 2 ♂♂, Cacaulândia, Fazenda Rancho Grande, Feb. 1997, Vaz-de-Mello leg. ( CEMT); 4 ♂♂, Cacaulândia (“ 62 km So. Ariquemes”), Fazenda Rancho Grande, 18–29 Sep. 1996, D.G. Marques leg. ( TAMU); 8 ♂♂, 7 ♀♀, Guajará-Mirim, 10º44′53.56″ S, 65º17′31.1″ W, 14–16 Feb. 2010, pitfall with human faeces, Fabricio Coletti leg. ( CEMT); 2 ♂♂ (1 dissected), 3 ♀♀, Guajará-Mirim, 10º44′57.62″ S, 65º19′0.32″ W, 138 m, 24 Feb. 2010, cow spleen, F. Coletti leg. ( CEMT); 1 ♂, Guajará-Mirim, Fazenda Agropecuária A.J.B., 10º37′59″ S, 64º59′58″ W, 180 m, 15 Jan. 2010, cow spleen, F. Coletti leg. ( CEMT); 1 ♂, 2 ♀♀, Itapuã do Oeste, Floresta Nacional [“FloNa”] do Jamari, 09º11′25″ S, 63º06′49″ W, 20 Feb. 2013, Mazama gouazoubira dung, J.F. Cerveira leg. ( CEMT); 3 ♂♂, Itapuã do Oeste, Floresta Nacional [“FloNa”] do Jamari, 09º05′20″ S, 63º09′47″ W, 24 Feb. 2013, Mazama nana dung, J.F. Cerveira leg. ( CEMT); 1 ♂, Ji-Paraná (“V. Rondônia ”, or Vila de Rondônia), 25 Jan.–9 Feb. 1961, Pereira and A. Machado leg. ( MZSP); 1 ♂, “Marmelo”, Rio Abunã, Nov. 1962, W. Bokermann leg. ( MZSP); 1 ♂, 1 ♀, Porto Velho, Abunã, 09º36′38″ S, 65º21′33″ W, 200 m, 19 Nov. 2010, pitfall with human faeces, J.C.F. Falcão leg. ( CEMT); 2 ♂♂, Porto Velho, Abunã, 09º35′03″ S, 65º22′36″ W, 7 Apr. 2011, pitfall with human faeces, L.R. Silva and M.F. Souza leg. ( CEMT); 1 ♂, 3 ♀♀, Porto Velho, Bom Jesus, Rio das Garças, 08º49′47″ S, 63º46′51″ W, 5 Apr. 2017, flight interception trap, D.C. Santos and K.K.G. Silva leg. ( CEMT); 3 ♂♂, 3 ♀♀, Porto Velho, Caiçara, 09º26′59″ S, 64º48′42″ W, 27 Jun. 2010, flight interception trap, J.C.F. Falcão and L.R. Silva leg. ( CEMT); 1 ♂, Porto Velho, Caiçara, 09º26′42″ S, 64º49′41″ W, 20 Mar. 2011, pitfall with human faeces, M.F. Souza and L.R. Silva leg. ( CEMT); 2 ♂♂, 6 ♀♀, Porto Velho, Caiçara, 09º26′42″ S, 64º49′41″ W, 29 Mar. 2011, pitfall with human faeces, M.F. Souza and L.R. Silva leg. ( CEMT); 1 ♂, 1 ♀, Porto Velho, Caiçara, 09º26′59″ S, 64º48′42″ W, 30 Mar. 2011, flight interception trap, L.R. Silva and M.F. Souza leg. ( CEMT); 1 ♂, Porto Velho, Caiçara, 09º26′10″ S, 64º49′59″ W, 10 Jan. 2012, flight interception trap, M.F. Souza leg. ( CEMT); 3 ♂♂, 3 ♀♀, Porto Velho, ESEC Cuniã, 08º04′11.82″ S, 63º28′34.64″ W, 4–6 Dec. 2011, pitfall with bananas, M.A.P.A. Silveira leg. ( CEMT); 1 ♂, Porto Velho, ESEC Cuniã, 08º04′11.82″ S, 63º28′34.64″ W, 4–6 Dec. 2011, pitfall with human faeces, M.A.P.A. Silveira leg. ( CEMT); 1 ♂, Porto Velho, ESEC Cuniã, 08º04′11.82″ S, 63º28′34.64″ W, 5–7 Dec. 2011, pitfall with bananas, M.A.P.A. Silveira leg. ( CEMT); 9 ♂♂, 10 ♀♀, Porto Velho, ESEC Cuniã, 08º04′11.82″ S, 63º28′34.64″ W, 9–11 Dec. 2011, pitfall with chicken meat, M.A.P.A. Silveira leg. ( CEMT); 1 ♂, Porto Velho, ESEC Cuniã, 08º04′11.82″ S, 63º28′34.64″ W, 8–12 Aug. 2012, pitfall with bananas, M.A.P.A. Silveira leg. ( CEMT); 2 ♂♂, Porto Velho, ESEC Cuniã, 08º04′11.82″ S, 63º28′34.64″ W, 15 Aug. 2012, pitfall with human faeces, M.A.P.A. Silveira leg. ( CEMT); 1 ♂, 1 ♀, Porto Velho, ESEC Cuniã, 08º04′11.82″ S, 63º28′34.64″ W, 19 Oct. 2012, pitfall with human faeces, M.A.P.A. Silveira leg. ( CEMT); 1 ♂, 1 ♀, Porto Velho, Mutum-Paraná [“Mutum”], 09º38′36″ S, 65º26′54″ W, 14 May 2010, pitfall with human faeces, F. Albertoni and L.R. Silva leg. ( CEMT); 2 ♂♂, 2 ♀♀, Porto Velho, Mutum- Paraná [“Mutum”], 09º35′ S, 65º03′ W, 250 m, 15–17 Sep. 2010, human faeces, J.C.F. Falcão and L.R. Silva leg. ( CEMT); 3 ♂♂, 1 ♀, Porto Velho, near Mutum-Paraná, “Prainha”, 12 May 2010, cow dung, F.F. Albertoni and L.R. Silva leg. ( CEMT); 1 ♀, Porto Velho, Nova Mutum-Paraná, 09º26′25″ S, 60º13′19″ W, 27–28 Feb. 2010, human faeces, L.R. Silva and R.L.R. Silva leg. ( CEMT); 4 ♂♂, 2 ♀♀, Porto Velho, Nova Mutum-Paraná, 09º26′55″ S, 61º09′57″ W, 1–2 Mar. 2010, human faeces, L.R. Silva and R.L.R. Silva leg. ( CEMT); 3 ♂♂, 4 ♀♀, Porto Velho, Nova Mutum-Paraná [“Mutum”], 09º35′46″ S, 65º02′27″ W, Jan. 2012, R.V. Nunes leg. ( CEMT); 1 ♂, Porto Velho, Nova Mutum-Paraná [“Mutum”], 09º38′36″ S, 65º26′54″ W, 3 Apr. 2012, flight interception trap, M.S.G. Carvalho leg. ( CEMT); 1 ♂, Porto Velho, Nova Mutum-Paraná [“Mutum”], 09º34′49″ S, 65º03′13″ W, 3 Apr. 2012, pitfall with human faeces, M.S.G. Carvalho leg. ( CEMT); 7 ♂♂, 4 ♀♀, Porto Velho, Nova Mutum-Paraná [“Mutum”], 09º35′46″ S, 65º02′27″ W, 250 m, Nov. 2012, pitfall with human faeces, R.V. Nunes leg. ( CEMT); 2 ♂♂, Rolim de Moura, 11º44′4.33″ S, 61º55′10.39″ W, 218 m, 8–10 Dec. 2015, pitfall with human faeces, D.C. Castro and N.S.G.F. Adem leg. ( CEMT); 1 ♂, 1 ♀, Rolim de Moura, 11º44′05″ S, 61º55′11.62″ W, 219 m, 8–10 Dec. 2015, pitfall with human faeces, D.C. Castro and N.S.G.F. Adem leg. ( CEMT); 3 ♂♂, 5 ♀♀, Rolim de Moura, 11º44′3.83″ S, 61º55′10.97″ W, 220 m, 8–10 Dec. 2015, pitfall with human faeces, D.C. Castro and N.S.G.F. Adem leg. ( CEMT); 4 ♂♂, 2 ♀♀, Rolim de Moura, 11º44′3.83″ S, 61º5510.97″ W, 221 m, 8–10 Dec. 2015, pitfall with human faeces, D.C. Castro and N.S.G.F. Adem leg. ( CEMT); 3 ♂♂, 4 ♀♀, Rolim de Moura, 11º44′3.91″ S, 61º55′12.11″ W, 222 m, 8–10 Dec. 2015, pitfall with human faeces, D.C. Castro and N.S.G.F. Adem leg. ( CEMT); 3 ♂♂, 2 ♀♀, Rolim de Moura, 11º44′3.33″ S, 61º55′11.53″ W, 223 m, 8–10 Dec. 2015, pitfall with human faeces, D.C. Castro and N.S.G.F. Adem leg. ( CEMT); 2 ♀♀, Vilhena, Nov. 1987, O. Roppa and P. Magno leg. ( MNRJ).
COLOMBIA: Amazonas: 1 ♀, Leticia, Monilla Amena, 27 Jun. 1997, pitfall with human faeces during night time, I. Quintero leg. ( CEMT). – Guaviare: 1 ♂, El Retorno, Elí farm., 02º21′46″ N, 72º38′29″ W, 15 Jul. 2007, Lagothrix lagotricha dung, Zárate leg. ( CEMT); 1 ♂, San José del Guaviare, Finca La Esmeralda, 02º33′ N, 72º37′ W, 240 m, 6 Oct. 2003 ( TAMU); 1 ♂, San José del Guaviare, Finca La Esmeralda, 02º33′ N, 72º37′ W, 240 m, 7 Oct. 2003 ( TAMU); 1 ♂, San José del Guaviare, Finca La Esmeralda, 02º33′ N, 72º37′ W, 240 m, 12 Jul. 2007 ( TAMU). – Meta: 1 ♂, 1 ♀, Parque Nacional Natural Tinigua, Centro de Investigaciones Ecológicas La Macarena, Rio Duda, 02º40′ N, 74º10′ W, 350 m, Jan. 1996, human faeces, I. Jiménez leg. ( CEMT).
ECUADOR: Orellana: 1 ♂, 1 ♀, Estación Científica Yasuní, 215 m, 5–10 Sep. 1999, E.G. Riley leg. ( CEMT); 19 ♂♂, 14 ♀♀, same collecting data as for preceding ( TAMU); 5 ♂♂, 1 ♀♀, Parque Nacional Yasuní (“Scyasuni”), 200 m, 25–27 Oct. 1997, D. Padilla and I. Tapia leg. ( CEMT); 1 ♀, same collecting data as for preceding ( MCNZ); 2 unsexed specimens, Payamino Research Station, 0º29′36″ S, 77º17′29″ W, 300 m, Jul.–Aug. 2007, Gillett and Orellana leg. ( OUMNH); 2 ♂♂, Puerto Francisco de Orellana (“Coca”), R. Haensch leg. ( ZMBH); 5 ♂♂, 5 ♀♀, Rodrigo Borja, IAMOE, 4 Jun. 2000, pitfall with human faeces, A. Dávalos leg. ( CEMT); 5 ♂♂, 3 ♀♀, 21 unsexed specimens, Tiputini Biodiversity Station, 0º38′ S 76º09′ W, 220 m, Sep. 2000, pitfall with human faeces, D. Inward leg. ( BMNH); 1 ♂, 1 ♀, same collecting data as for preceding ( MUSM); 2 ♂♂, 6 ♀♀, Yasuni Scientific Research Station, 9–17 Sep. 1999, D.G. Marqua leg. ( TAMU).
PERU: 1 unsexed specimen, Río Tambopata, Jan.–Apr. 1995, B.J. Selman leg. ( BMNH). – Cuzco: 2 ♀♀, La Convención, Echarate, Comunidad Nativa de Timpia, 12º06′41.38″ S, 72º49′59.34″ W, 429 m, 22 Oct. 2009, M. Alvarado and E. Rázuri leg. ( MUSM); 1 ♀, La Convención, Echarate, Comunidad Nativa de Timpia, 12º06′47.61″ S, 72º49′32.67″ W, 536 m, 25–31 Jan. 2010, E. Razuri and C. Espinoza leg. ( MUSM). – Huánuco: 1 ♀, Estación Biológica Panguana (Forschungsstation Panguana), Rio Pachitea, Rio Yuyapichis, 09º37′ S, 74º56′ W, 260 m, 28 Aug.–14 Sep. 1986, Listabarth leg. ( NHMW); 1 ♂, Puerto Inca, Clayton, 09º11′55.37″ S, 74º55′12.1″ W, 243 m, 16–18 Jun. 2009, C. Carranza leg. ( MUSM). – Junín: 1 ♀, Satipo, Oct.–Nov. 2002 ( CEMT); 2 ♂♂, Satipo, 600 m, 23 May–3 Jun. 2004, A. Santibañez leg. ( TAMU); 2 ♂♂, 2 ♀♀, Satipo, 1100 m, 4–15 Apr. 2003 ( CEMT); 2 ♂♂, 1 ♀, Satipo, Fundo Santa Teresa, 2 Oct. 2007, M. Alvarado and L. Figueroa leg. ( MUSM); 1 ♂, Satipo, Rio Tambo, Campamento Kampitiare, Comunidad Nativa Shevoja, 11º42′ S, 73º01′ W, 250 m, 10 Jan.–7 Feb. 2005, Williams Paredes leg. ( MUSM). – Loreto: 1 ♀, Loreto, Urarinas, right bank of Urituyacu river, 140 m, 18 Nov. 2009, L. Sulca leg. ( MUSM); 2 ♂♂, Loreto, Urarinas, Rio Copalyacu, 03º42′59″ S, 75º26′00″ W, 161 m, 7–8 Oct. 2009, L. Sulca leg. ( MUSM); 1 ♂, Maynas, Iquitos, 4 Nov. 2005, L. Culot leg. ( MUSM); 1 unsexed specimen, Maynas, Iquitos, 03º96′46″ S 73º15′49″ W, Dec. 1997, M.V.L. Barclay leg. ( BMNH); 1 ♂, Maynas, Iquitos, Agua Blanca, 02º55′16.3″ S, 74º49′0.1″ W, 160–177 m, ‘bait trap’, Felipe Meza Velez leg. ( MUSM); 1 ♂, Maynas, Miraflores, 03º47′4.8″ S, 73º41′43″ W, 69 m, 10 May 2012, C. Ampudia leg. ( MUSM); 1 ♂, Maynas, Napo, Rio Curacay, Gaseoducto Barrett, 16–18 Mar. 2008, W. Paredes leg. ( MUSM); 1 ♂, Requena, Masisea, Comunidad Nativa Santa Teresa, 06º00′55.07″ S, 74º04′59.95″ W, 87 m, 12 Aug. 2008, C. Carranza leg. ( MUSM); 1 ♂, Requena, Sierra del Divisor (“a 5 km del Rio Capanagua”), 06º19′30″ S, 73º46′09″ W, 160 m, 20–21 Oct. 2008, A. García leg. ( MUSM); 1 ♀, Requena, Sierra del Divisor (“cerca de Rio Punga”), 06º55′11.2″ S, 73º50′47″ W, 222 m, 22–24 Oct. 2008, C. Espinoza leg. ( MUSM); 3 ♂♂, 2 ♀♀, Rio Ungumayo, 04º13′56″ S, 76º48′43″ W, 171 m, 29–30 Nov. 2009, L. Sulca leg. ( MUSM); 1 ♂, 1 ♀, Ucayali, Sierra del Divisor (“cerca de Sarayacu”), 06º57′21.06″ S, 74º01′27.4″ W, 195 m, 10–11 Oct. 2008, C. Espinoza leg. ( MUSM). – Madre De Díos: 10 ♂♂, 2 ♀♀,Parque Nacional del Manu, Estación Biológica Cocha Cashu, 11º55′ S, 77º18′ W, 380 m, 18 Aug.–5 Sep. 1986, D.C. Darling and A.B. Forsyth leg. ( MUSM); 1 ♀, Parque Nacional del Manu, Estación Biológica Cocha Cashu, 11º55′ S, 77º18′ W, 380 m, 20–22 Aug. 1986, D.C. Darling leg. ( MUSM); 1 ♂, Parque Nacional del Manu, Pakitza, 11º53′ S, 70º58′ W, 400 m, 16 Nov. 1990, J. Macdonald leg. ( CEMT); 1 ♂, Manu, Centro de Investigación y Capacitación Río Los Amigos ( CICRA), Yugunturo trail, 12º33′ S, 70º05′ W, 283 m, 16 Nov. 2006, flight interception trap, A. Asenjo leg. ( MUSM); 1 ♂, Manu, CICRA, 12º34′ S, 70º05′ W, 280 m, 10–14 Jul. 2010, C. Chaboo leg. ( MUSM); 1 ♂, Manu, Reserva Comunal Amarakaeri, naciente Qda. Pinquiri, 12º55′43″ S, 70º51′41″ W, 421 m, 4 Jun. 2011, M. Vilchez leg. ( MUSM); 1 ♀, Reserva Biológica CICRA [“Centro de Investigación y Capacitación Río Los Amigos”], 23 Jan. 2007, flight interception trap, Angelico Asenjo leg. ( CEMT); 1 ♂, Rio Palma Real Grande, Limon, 12º32′20″ S, 68º51′41″ W, 400 m, 5–6 Apr. 1999, pitfall with human faeces, T. Larsen leg. ( MUSM); 1 ♀, Rio Palma Real Grande, Limon, 12º32′20″ S, 68º51′40″ W, 220 m, pitfall with human faeces, T. Larsen leg. ( MUSM); 2 ♂♂, 1 ♀, Tambopata, 12º27′49.27″ S, 69º07′30.69″ W, 233 m, 17–19 Apr. 2011, O. Huaches leg. ( MUSM); 1 ♀,Tambopata, 300 m, 18–22 Apr. 1983, E. Perez leg. ( MUSM); 1 ♂, Tambopata, Inambari, Primavera Baja, 12º54′ S, 70º05′ W, 234 m, 4–11 Nov. 2009, C. Castillo and E. Martinez leg. ( MUSM); 1 ♂, 1 ♀, Tambopata, Puerto Maldonado, Madama, 12º31′20″ S, 69º03′44″ W, 19–20 Jul. 2009, 182 m, M. Alvarado leg. ( MUSM); 1 ♂, 1 ♀, Tambopata, Puerto Maldonado, sector Triunfo, 12º33′42″ S, 69º11′47″ W, 198 m, 23 Jul. 2009, M. Alvarado leg. ( MUSM); 1 ♂, 1 ♀, Tambopata, Puerto Madonado, Sudadero, 12º21′19″ S, 69º01′48″ W, 221 m, 21–22 Jul. 2009, M. Alvarado leg. ( MUSM); 2 ♂♂, 1 ♀, Tambopata, Reserva Nacional Tambopata, Explorer’s Inn, 12º50′30″ S, 69º17′31″ W, 161 m, 15–18 May 2009, L. Figueroa and M. Alvarado leg. ( MUSM); 1 ♂, Tambopata, Rio Tambopata, Colpa de Guacamayos, 13º08.5′ S, 69º36.4′ W, 300 m, Oct. 1995, A. Forsyth leg. ( MUSM). – San Martín: 1 ♂, Mariscal Cáceres, Juanjuí, 7 Jul. 2007, P. Melendes leg. ( TAMU); 1 ♀, San Martín, El Porvenir, Bosque El Pelejo, 06º19′28.3″ S, 81º50′38.4″ W, 161 m, 17–19 May 2010, J. Robledo leg. ( MUSM); 1 unsexed specimen, San Pedro de Cumbasa (“Cumbase”) ( MNHN). – Ucayali: 1 ♂, Coronel Portillo, 08º20′40.04″ S, 73º42′42.64″ W, 212 m, 10–12 Oct. 2012, P. Sanchez leg. ( MUSM); 1 ♀, Coronel Portillo, Binocura, 08º53′16.9″ S, 74º00′0.21″ W, 140 m, 15–16 Jul. 208, M. Alvarado leg. ( MUSM); 2 ♂♂, 2 ♀♀, Coronel Portillo, Puerto Alegre, 08º44′6.76″ S, 74º09′4.54″ W, 196 m, 21–22 May 2008, L. Figueroa leg. ( MUSM); 1 ♀, Coronel Portillo, Puerto Purin, 08º45′16.7″ S, 74º10′16.8″ W, 125 m, 17–18 May 2008, L. Figueroa leg. ( MUSM); 2 ♂♂, 4 ♀♀, Coronel Portillo, Puerto Purin, 08º44′59.2″ S, 74º08′19.52″ W, 122 m, 19 May 2008, L. Figueroa leg. ( MUSM); 3 ♂♂, 1 ♀, Coronel Portillo, Rio Inamapuya, 08º44′33.7″ S, 74º06′15.9″ W, 135 m, 23–24 May 2008, L. Figueroa leg. ( MUSM); 1 ♀, Coronel Portillo, Rio Tamaya, 08º52′32.3″ S, 74º06′52.1″ W, 155 m, 11–12 May 2008, L. Figueroa leg. ( MUSM); 1 ♂, Coronel Portillo, Rio Tamaya, 08º52′39.2″ S, 74º07′7.1″ W, 155 m, 13–14 May 2008, L. Figueroa leg. ( MUSM); 1 ♀, Coronel Portillo, Rio Tamaya, 08º51′41.9″ S, 74º04′20.6″ W, 150 m, 15–16 May 2008, L. Figueroa leg. ( MUSM); 1 ♂, Padre Abad, Alexander von Humboldt, 08º48′46.2″ S, 75º04′15.7″ W, 245 m, 10–11 Jun. 2009, C. Carranza leg. ( MUSM).
No data: 3 ♂♂ ( CEMT – ex V.M. Diéguez M. coll.); 1 ♂ ( ISNB).
Redescription
COLOURATION. Head, pronotum, elytra, pygidium, and underside – including ventral surface of femora – with diffuse shine. Head, pronotum, and pygidium with blue, purple, or greenish reflections; in some specimens, pronotum with strong olive green shine. Elytra dark and with silky aspect; usually dark blue or purple; in some specimens, totally black or with greenish reflections. Meso- and metafemora dark brown and usually with greenish reflections.
HEAD. Tegument with silky aspect and micropunctation almost absent, more evident only on frons ( Fig. 6D View Fig ). Clypeus with two small apical teeth obtuse and contiguous at base; with a single transverse row of short setae covering base of both teeth. Genae with acute tooth immediately behind clypeal-genal juncture. Posterior edge of head usually with a fine line between eyes which can be effaced on the sides, or even totally absent.
THORAX. Pronotum with tegument at centre ranging from bright with dense micropunctation and without microsculpture to with strong alveolar microsculpture obliterating micropunctation; lateral region always without micropunctation, with alveolar microsculpture present and tegument perceptibly more matte than at centre. Posterior edge without fine transverse line at centre. Hypomeral cavity with very short yellowish setae at centre. Metaventrite entirely glabrous (occasionally, with some setae on the sides close to metacoxae); micropunctation indistinct at centre and visible only on region adjacent to apices of metacoxae.
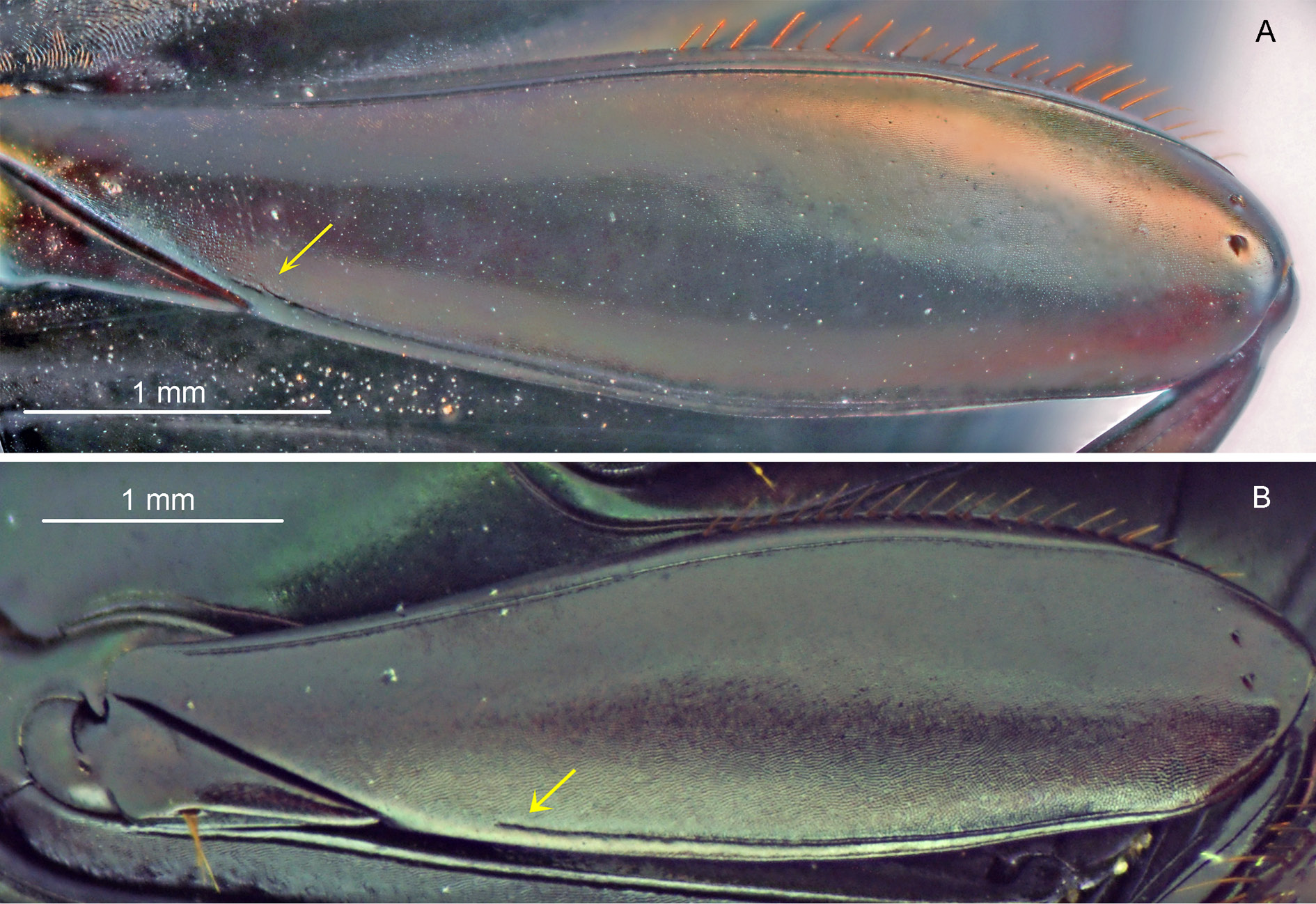
LEGS. Ventral surface of all femora and tibiae with diffuse shine of silky aspect. Protibiae with three obstuse or slightly acute teeth at their apical third, the two apical teeth of similar size and larger than the basal one. Mesofemora margined anteriorly only at their basal half; unmargined portion of anterior edge with row of very short setae. Metafemora with both anterior and posterior magins; posterior margin incomplete, always present at apical half and ranging in length at basal half, but never reaching trochanter ( Fig. 31B View Fig ). Metatarsomeres II and V subequal in length and longer than the others; metatarsomere IV shorter than the others.
ELYTRA. Nine fine elytral striae only weakly marked, almost imperceptible. Base of striae I–V carinulate and well marked. Tegument of interstriae with a silky shine and with dense three-dimensional alveolar microsculpture obliterating indistinct micropunctation. Fine humeral carina distinct and long, sometimes present throughout basal fourth of elytra ( Fig. 12C View Fig ).
ABDOMEN. Ventrites V–VI smooth, shiny and with micropunctation at centre. Lateral foveae absent in both sexes. Pygidium with diffuse shine, with well-marked alveolar microsculpture and without micropunctation.
AEDEAGUS. Parameres long, little shorter than phallobase and clearly asymmetrical: left paramere laterally with apical depression much deeper and wider than depression on right paramere (difference more easialy seen in dorsal view). In lateral view, parameres with central angulosity and without ventral keel and notch ( Fig. 17F View Fig ).
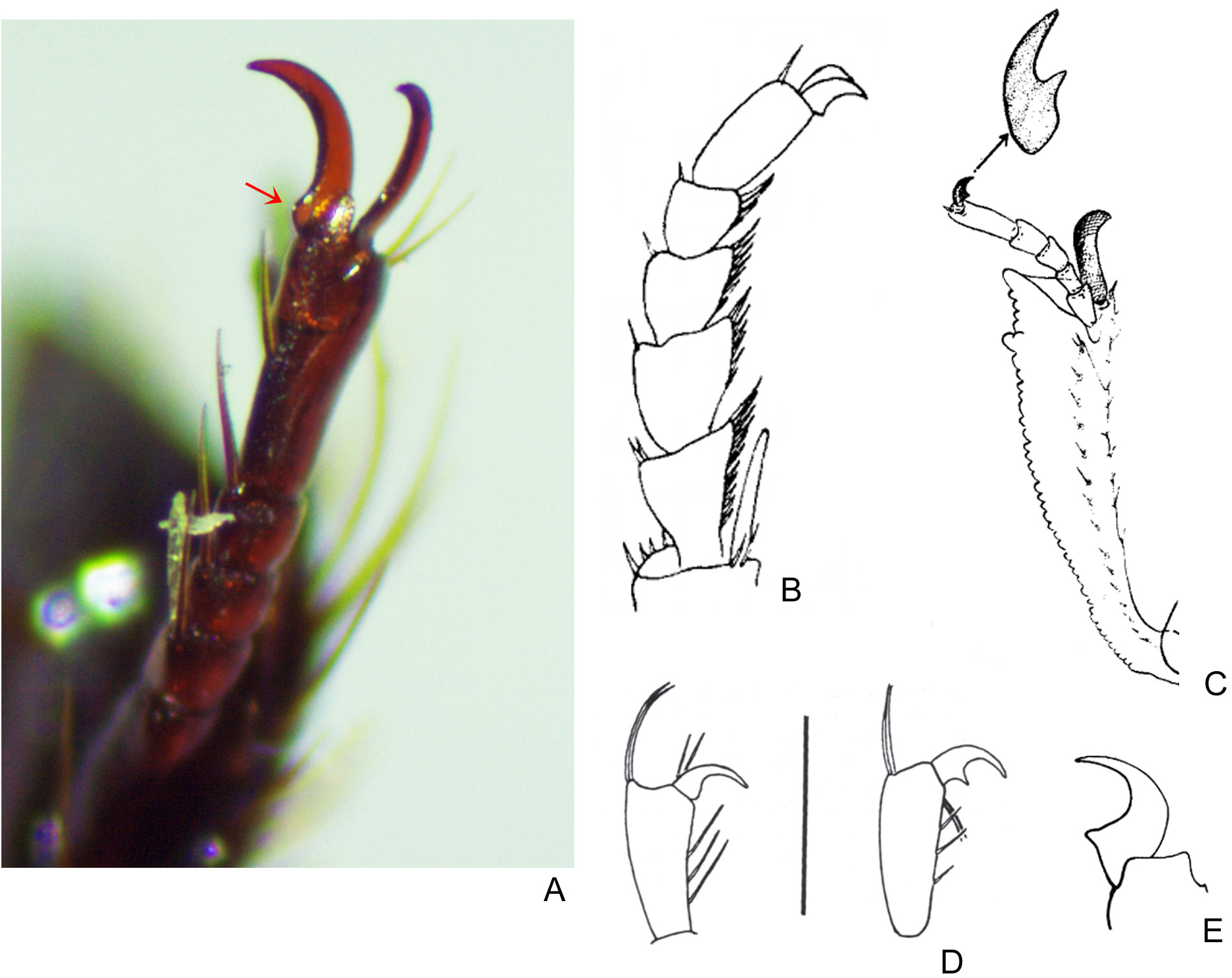
SEXUAL DIMORPHISM. Males: Protibial spur broad and foliaceous, external edge extended into an acute long spine, while internal edge has no prolongation or has much shorter spine; area between both spines straight or slightly excavated ( Fig. 15G View Fig ). Pygidium very long (length between 1.9 and 2.9 mm) and convex; apical margin of pygidium much wider than lateral ones. Ventrite VI strongly narrowed at middle. Females: Protibial spur wide and with strong apical incision, with two long spines of uneven length, the internal spine shorter than the external one ( Fig. 15H View Fig ). Pygidium shorter (between 1.5 and 2.2. mm) and flat; apical margin only slightly wider than lateral ones. Ventrite VI as wide at middle as on sides.
Measurements
Males (N= 40). TL: AV: 12.1 ± 0.78; MX: 15; MN: 10.5. EW: AV: 8.2 ± 0.36; MX: 8.8; MN: 7.4. PL: AV: 4 ± 0.25; MX: 4.5; MN: 3.4. PW: AV: 7.5 ± 0.41; MX: 8.5; MN: 6.5. PgL: AV: 2.2 ± 0.18; MX: 2.9; MN: 1.9. PgW: AV: 3.8 ± 0.22; MX: 4.3; MN: 3.1.
Females (N = 20). TL: AV: 11.8 ± 1; MX: 14; MN: 9.8. EW: AV: 7.9 ± 0.6; MX: 8.7; MN: 6.1. PL: AV: 3.8 ± 0.31; MX: 4.2; MN: 2.9. PW: AV: 7.1 ± 0.54; MX: 7.9; MN: 5.7. PgL: AV: 1.9 ± 0.18; MX: 2.2; MN: 1.5. PgW: AV: 3.8 ± 0.28; MX: 4.2; MN: 3.
Geographical distribution
Widespread in the Amazon Basin.
Ecoregions
Negro-Branco Moist Forests, Caquetá Moist Forests, Napo Moist Forests, Cordilleira Oriental Montane Forest , Ucayali Moist Forests, Iquitos Varzea, Southwest Amazon Moist Forests, Purus Varzea, Bolivian Yungas, Purus-Madeira Moist Forests, Monte Alegre Varzea, Madeira-Tapajós Moist Forests, Mato Grosso Tropical Dry Forests, Tapajós-Xingu Moist Forests, Xingu-Tocantins-Araguaia Moist Forests, Tocantis-Pindaré Moist Forests.
Collecting sites ( Fig. 30 View Fig )
COLOMBIA. Meta: Parque Nacional Natural Tinigua (Centro de Investigaciones Ecológicas La Macarena). Guaviare: El Retorno, Parque Nacional Natural Nukak, San José del Guaviare. Amazonas: Leticia (Monilla Amena).
ECUADOR. Sucumbíos: “Chiruisla Station”. Orellana: Parque Nacional Yasuní, Payamino Research Station, Puerto Francisco Orellana, Tiputini Biodiversity Station. Morona Santiago: Cordillera de Cutucú.
PERU. Loreto: Loreto (Urarinas), Maynas (Iquitos, Miraflores, Napo), Requena (Masisea: Comunidad Nativa Santa Teresa; Sierra del Divisor), Ucayali (Sierra del Divisor). San Martín: Mariscal Cáceres (Juanjuí), San Martín (El Porvenir), San Pedro de Cumbasa. Huánuco: Estación Biológica Panguana, Puerto Inca (Clayton). Ucayali: Coronel Portillo (Puerto Alegre, Puerto Purin), Padre Abad (Alexander von Humboldt). Junín: Satipo. Cuzco: La Convención (Echarate). Madre de Dios: Manu (Centro de Investigación y Capacitación Río Los Amigos, Reserva Comunal Amarakaeri), Parque Nacional del Manu (Estación Biológica Cocha Cashu), Reserva Biológica “Centro de Investigación y Capacitación Río Los Amigos” (CICRA), Tambopata (Inambari: Primavera Baja; Puerto Maldonado; Reserva Nacional Tambopata).
BRAZIL. Amazonas: Benjamin Constant, Beruri, Manaus, Manicoré, São Paulo de Olivença, Tefé. Acre: Mâncio Lima, Manoel Urbano (Parque Estadual Chandless), Rio Branco, Senador Guiomard, Xapuri (Reserva Extrativista Chico Mendes). Rondônia: Cacaulândia, Guajará-Mirim, Itapuã do Oeste (Floresta Nacional do Jamari), Ji-Paraná, Rolim de Moura, Vilhena. Pará: Almeirim (Monte Dourado), Alter do Chão, Belém, Belterra, Novo Progresso, Paragominas, Redenção, Santarém, São Félix do Xingu, Tailândia. Maranhão: Centro Novo do Maranhão. Mato Grosso: Alta Floresta, Carlinda, Cotriguaçu, Nova Bandeirante, Novo Mundo (Parque Estadual do Cristalino).
BOLIVIA. La Paz: Nor Yungas. Cochabamba: José Carrasco (Valle del Sacta), Chapare. Santa Cruz: Sara.
Intraspecific variation and taxonomic discussion
Although formally described only in 1949, it is possible to trace back the beginning of the taxonomic history of S. proseni to the late 19 th century, when Bates (1887) was the first author to realize that the South American populations of ‘ Canthon aequinoctialis ’ differed from the Central American ones in having individuals of considerable different size: “Central-American differ from South-American exemples only in being smaller (11 millim.), the latter varying little from 14 millim” ( Bates 1887: 33). Indeed, Henry Walter Bates himself collected some specimens of S. proseni in the Amazon region which were examined for this work. Nonetheless, over the following 60 years, no author noticed those differences or discovered others, treating under the same name C. aequinoctialis specimens coming from both Central America and the Amazon Basin, including the revisions of Schmidt (1922) and Balthasar (1939).
Eventually, already in the mid-20 th century, Martínez (1949) described Glaphyrocanthon proseni stating his new species was close to C. aequinoctialis , but distinct from this latter species by the metafemora with a margin on both edges (while C. aequinoctialis would have only the anterior margin) and colouration. It is clear, however, that Martínez did not have on hand specimens of S. aequinoctialis , since both species have metafemora with posterior margin and, actually, this margin is even longer in S. aequinoctialis than in S. proseni ( Fig. 31 View Fig ) (indeed, Martínez wrote “S egún la descripción ” when referred to C. aequinoctialis ).
Later, however, Pereira & Martínez (1956), in a key to the species then-positioned in Glaphyrocanthon , noted colouration differences between the species, G. proseni being blue-violet and opaque (“azul violeta e opaco”) and G. aequinoctialis being black and bright (“preto e brilhante”). Martínez et al. (1964) refined those descriptions writing that G. proseni had indistinct elytral striae, and the elytra, pygidium and ventral surface of femora opaque, while G. aequinoctialis had fine, but perceptible elytra striae and the entire body bright; they were also the first authors to note that S. aequinoctialis was restricted to Central and northern South America . But despite these considerations, they also wrote that, the differences between the two taxa being restricted to the microsculpture and elytral striae only, G. proseni would possibly represent a southern subspecies of G. aequinoctialis instead of a full species. Then, Vulcano & Pereira (1967), in a key to the dung beetle species of Amazonia, distinguished the two species, apart from their distribution, only by colouration and size (resuming, for the first time and, probably independently, the observation taken by Bates 1887). Lastly, Halffter & Martínez (1977), after having studied some type specimens of both names ( Fig. 29 View Fig ), synonymized C. aequinoctialis and G. proseni without presenting, however, the reasons for that act, although it is reasonable to imagine they did so based on the arguments given by Martínez et al. (1964). In this way, except for some recent faunistic inventories that treated C. proseni as a valid species (e.g., Vaz-de-Mello et al. 2011b; Korasaki et al. 2012; Nunes et al. 2014; Chamorro et al. 2018), these two names have remained in synonymy ever since. Next, we present evidence that, in our view, supports the present revalidation of S. proseni and demonstrates its independence from S. aequinoctialis .
As noted by Bates (1887) (despite having considered them different populations of a same species), the average body size of S. aequinoctialis and S. proseni is truly distinct. The average body length of S. proseni is 12 mm, ranging from individuals measuring only 9.8 mm to those of 15 mm, while S. aequinoctialis has an average length of 10 mm, with the smallest specimens measure 8.7 mm to the largest ones 12.3 mm. It is interesting to note that the lowest value recorded for S. proseni is only slightly lower than the average of S. aequinoctialis , while the largest S. aequinoctialis examined is only a little longer than the average of the specimens of S. proseni . This pattern is repeated in all the other measurements taken for this work, including the elytral width (average of 8.1 mm for S. proseni , with a maximum of 8.8, a minimum of 5.7 and an average of 7.0 mm for S. aequinoctialis , with a maximum of 7.8 and a minimum of 5.7 mm) and the greatest width of the pronotum (an average of 7.3 mm, with a maximum of 8.2. and a minimum of 5.7 mm for S. proseni , and an average of 6.3, with a maximum of 7.4 and a minimum of 5.3 for S. aequinoctialis ).
Also part of the group of characteristics already used in the past to distinguish both species are the differences in microsculpture and other aspects of the tegument, which form one of the strongest indications for the evolutionary independence of S. aequinoctialis and S. proseni . By seeing them with the naked eye, it is possible to realize that the pronotum, ventral surface of femora, pygidium and especially the elytra of S. aequinoctialis are very bright and have a smooth surface, while they reflect light in a more diffuse way and, consequently, have a more opaque, silky appearance in S. proseni . This is so thanks to the microsculpture present in the tegument of these two species. In S. aequinoctialis , the microsculpture is absent or very effaced at the centre of the pronotum and on the elytra, and, although clearly present, it is very fine (i.e., the microsculpture’s ‘alveoli’ are very small) on the femora and pygidium. This condition makes the tegument of this species (or gives the appearance of being) much smoother than that of S. proseni , which has a strong alveolar microsculpture covering the entire pronotum, femora, elytra and pygidium ( Fig. 12A View Fig ).
The elytral striae are also much more marked and visible in S. proseni than in S. aequinoctialis , where they are almost imperceptible; this difference is more easily seen looking at the base of elytra, where the striae are finely marked and expanded in S. proseni and completely indistinct in S. aequinoctialis . Regarding the micropunctation, both species differ in the way the micropunctures are marked. In the majority of the specimens of S. proseni (but not all; see below), the pronotum, elytra and pygidium have a very visible micropunctation, which, in general, is not strongly obliterated by the microsculpture. In S. aequinoctialis , on the other hand, punctures may be completely absent or, more commonly, they are only weakly marked and are almost imperceptible, having the appearance of being shallower than those of S. proseni . Also in relation to the tegument, we see differences between S. aequinoctialis and S. proseni in the humeral carina ( Fig. 12C View Fig ) (which is longer and more strongly marked in the latter species), on the posterior margin of the metafemora (the margin extends from the apex of metafemur to beyond or immediately before the trochanter in S. aequinoctialis ( Fig. 31A View Fig ), while it never reaches the trochanter and, in general, is present only slightly beyond the middle of metafemur in S. proseni , Fig. 31B View Fig ) and on the posterior margin of the head (which is always present in S. aequinoctialis , and may be incomplete or even completely absent is some specimens of S. proseni ).
Despite all the differences listed above, the strongest evidence for the isolation between these two species rests on the shape of the female protibial spur. In S. aequinoctialis , this spur is spiniform and has an acuminate apex ( Fig. 15F View Fig ), while females of S. proseni have a wider and profoundly bifid spur which has the external branch longer than the internal one (separation between both branches ‘V’-shaped, Fig. 15H View Fig ). In fact, the presence of this bifid spur distinguishes females of S. proseni from all the other females in the genus and is here reported for the first time.
Lastly, the disjunct geographical distribution of S. aequinoctialis and S. proseni , separated by the great barrier constituted by the Andes (Fig. 113), adds to the morphological evidence discussed above. Sylvicanthon aequinoctialis is present from Honduras in the north, south to the Choco region on the Pacific coast of South America , and in the great valley between the central and western mountain ranges in Colombia. In turn, S. proseni is distributed throughout most of the Amazon Basin, from Maranhão in the east to the Yungas on the eastern slopes of the Andes in Bolivia, Peru and Ecuador; in Colombia, the latter species was recorded only from the lower areas in the Amazon forest. Ecologically, both species do not seem to be very distinct from one another. Both inhabit a large variety of forest habitats with a very remarkable altitudinal range, living from low and coastal regions (e.g., S. proseni in Belém, on the banks of the Baía de Marajó, Brazil; S. aequinoctialis in La Ceiba on the Caribbean coast of Honduras) to the Andean regions above 1000 m (e.g., S. proseni in Satipo, Peru, at 1110 m a.s.l.; S. aequinoctialis in the Reserva Natural Cañon del Río Claro, Colombia, at 1440 m). In all those places, S. aequinoctialis and S. proseni are among the most abundant dung beetle species, and there they exploit a wide array of food sources (dung, carrion and rotten fruits). Therefore, despite the differences in morphology, both species seem to have little diverged in their life habits.
Being such an abundant and widely-distributed species, Sylvicanthon proseni would be expected to show a great degree of instraspecific variation, and this is indeed what occurs. The variation on the density of pronotal micropunctation is easily seen: even within a single given population it is possible to encounter specimens with a very densely-punctated tegument (and with a very bright pronotum) and others with a very fine, almost imperceptible micropunctation. The most common pattern, however, is the one with fine, but easily visible micropunctation. The size of the dorsal portion of the eyes also varies intraspecifically, the width ranging from approximately one-fifth to one-sixth of the interocular space. Lastly, a very important variation is seen on the posterior edge of the head: in the majority of the specimens a fine line runs across the entire interocular space. In some specimens, however, this line is effaced near the eyes and is marked only at the middle; progressively, this central line disappears, and in some individuals, it is completely absent.
Comments
Because this species was confused with S. aequinoctialis for such a long time, especially after the synonymy proposed by Halffter & Martínez (1977), most of what is known about S. proseni is published under the name Canthon aequinoctialis . Concerning specifically its geographical distribution, the records of Balthasar (1941, 1951) from “ Mishujacu, Iquitos ” ( Peru), Vaz-de-Mello (1999) from Acre ( Brazil), Celi et al. (2004) and Carpio et al. (2009) from Ecuador, Noriega-Alvarado (2004), Noriega et al. (2007a) and Noriega (2012) from Meta ( Colombia), Noriega et al. (2008) from Amazonas ( Colombia), and Figueroa & Alvarado (2011) from Tambopata ( Peru) certainly refer to S. proseni , not to S. aequinoctialis as originally mentioned.
The village of San Pedro de Cumbasa, in the department of San Martín, Peru, was included in the distribution of S. proseni based on a specimen in MNHN labelled “ Cumbase ” without any further information. Takiya et al. (2003), in their study of the leafhopper genus Hyogonia China, 1927 ( Hemiptera , Cicadellidae ), also studied a specimen labelled “Cumbase” and, according to them, this word probably refers to San Pedro de Cumbasa. Similarly, the register from Novo Centro do Maranhão, the only known locality of this species in the Brazilian state of Maranhão, was obtained from a single specimen housed at the MZSP labelled “ Igarapé Gurupi-Uma, Aldeia Araçu, 50 km E. de Canindé ”, information that probably refers to that locality according to Pinto & Lamas (2011).
Natural history
Sylvicanthon proseni is widely distributed in the Amazon Basin and seems to be a very common species in all the regions where it lives. In several dung beetle community inventories carried out in the Amazon region, S. proseni was one of the most abundant species (e.g., Noriega et al. 2007a: in Meta, Colombia, it corresponded to 4.66% of the collected specimens; Korasaki et al. 2012: Amazonas, Brazil, 5.6%; Vaz-de-Mello 1999: Acre, Brazil, 23%; Noriega et al. 2008: Amazonas, Colombia, 29.9%; Carpio et al. 2009: Sucumbíos, Ecuador, 55%). This species seems to be absent only in grasslands and agricultural fields, but, where some small patch of forest is left standing or there is secondary regeneration, S. proseni can be found ( Vaz-de-Mello 1999; Noriega et al. 2007a; Korasaki et al. 2012), including river floodplain areas ( Escobar 2000b; Nunes et al. 2014). These observations clearly show the species’ flexibility and its considerable resistance against anthropic impact.
The individuals studied for this work were collected in all months and within an altitudinal range between 68 and 1100 m (nonetheless, the greater part of the specimens was collected between 100 and 500 m). Although we are aware of just a single time-activity record – which was taken from the label of a specimen from Leticia, Colombia, collected at night –, the information that specimens were collected with a light trap (Vaz-de-Mello et al. 2011) and our knowledge about the other species of Sylvicanthon (especially S. aequinoctialis ) allows us to confidently believe that S. proseni is a nocturnal species.
Sylvicanthon proseni was mainly collected using human faeces as bait and it seems to be preferably coprophagous (Martínez et al. 1949; Vaz-de-Mello 1999; Noriega et al. 2007 a, 2008; Figueroa & Alvarado 2011; Korasaki et al. 2012; Nunes et al. 2015; and information from specimen labels). Some individuals collected in Porto Velho (Rondônia, Brazil), however, were caught using pitfall traps baited with chicken meat and rotten bananas. Furthermore, S. proseni was also collected on dung of the woolly monkey [ Lagothrix lagotricha (Humboldt, 1812) ] ( Noriega 2012) and cows (specimen labels information), with a flight interception trap ( Vaz-de-Mello 1999) and a light trap (Vaz-de-Mello et al. 2011).
Pereira & Martínez (1956) and Martínez et al. (1964) reported a very interesting case of phoretic behaviour between S. proseni and tapirs [ Tapirus terrestris (Linnaeus, 1758) ]. Nevertheless, neither work stated the primary source for that information (authors’ observation?) or any further detail (e.g., place of observation, number of individuals, or whether the tapir was alive or dead when the observation was made, which could rule out the possibility of simple necrophagy). Therefore, whether this information is reliable or not is yet to be resolved. Nevertheless, phoresy has indeed been observed in a great number of other dung beetles, including the relationship between two distinct lineages of Onthophagus and macropod marsupials (wallabies and rat-kangaroos) in Australia ( Matthews 1972). In the Neotropical region, phoresy was observed between species of the dung beetle genera Uroxys and Bradypodidium Vaz-de-Mello, 2008 with three-toed sloths [ Bradypus Linnaeus, 1758 ] ( Ratcliffe 1980; Howden & Young 1981; Vaz-de-Mello 2008), and species of Canthidium with the coppery titi monkey, Callicebus cupreus (Spix, 1823) ( Herrera et al. 2002). In Deltochilini, Pereira & Martínez (1956) reported phoretic behaviour of Glaphyrocanthon subhyalinus ( Harold, 1867) (cited as “ Canthon hyalinus ”) in the brown titi monkey, Callicebus brunneus (Wagner, 1842) , while Jacobs et al. (2008) described in detail the relationship between Glaphyrocanthon aff. quadriguttatus (Olivier, 1789) and both Callicebus brunneus and the saki monkey species Pithecia irrorate Gray, 1842 .
As hypothesized by Herrera et al. (2002) and Jacobs et al. (2008), this phoretic strategy may be related to the behaviour known as the ‘sit and wait’ strategy, which is largely beneficial to the dung beetles that employ it because, being mounted on the dung-producing animal itself and positioned around its anus, those beetles can reach the dung as soon as it is released. In doing so, they can start the consumption of the dung before it dries out and before the arrival of competitors. In tropical forest with a low density of large mammals such as those in South America , competition between dung beetles can be intense and, consequently, arriving first at the food source brings a considerable advantage. If the alleged phoresy of S. proseni in tapirs is confirmed, it will be the phoretic relationship between the largest host and the largest dung beetle yet discovered. Ratcliffe (1980) and Jacob et al. (2008) give an excellent discussion on the phoretic relationship between Scarabaeinae and mammals.








| MACN |
Museo Argentino de Ciencias Naturales Bernardino Rivadavia |
| MZSP |
Sao Paulo, Museu de Zoologia da Universidade de Sao Paulo |
| MNRJ |
Museu Nacional/Universidade Federal de Rio de Janeiro |
| MCNZ |
Porto Alegre, Museu de Ciencias Naturais da Fundacao Zoo-Botanica do Rio Grande do Sul |
| FIOC |
Fundacao Instituto Oswaldo Cruz |
| NHMW |
Naturhistorisches Museum, Wien |
| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Sylvicanthon proseni (Martínez, 1949)
| Cupello, Mario & Vaz-De, Fernando Z. 2018 |
Canthon (Canthon) proseni
| Chamorro W. & Marin-Armijos D. & Granda V. & Vaz-de-Mello F. Z. 2018: 87 |
| Vaz-de-Mello F. Z. & Silva R. L. R. & Nunes L. G. O. A. & Correa P. R. O. A. 2011: 82 |
Canthon proseni
| Silva R. J. & Pelissari T. D. & Krinski D. & Canale G. & Vaz-de-Mello F. Z. 2017: 490 |
| Silva R. J. & Coletti F. & Costa D. A. & Vaz-de-Mello F. Z. 2014: 348 |
| Nunes R. V. & Carvalho M. S. G. & Vaz-de-Mello F. Z. & Dattilo W. & Izzo T. J. 2014: 410 |
| Korasaki V. & Vaz-de-Mello F. Z. & Braga R. F. & Zanetti R. & Louzada J. 2012: 425 |
| Jacobs J. & Nole I. & Palminteri S. & Ratcliffe B. 2008: 642 |
Canthon (Canthon) aequinoctialis
| Noriega J. A. & Realpe E. & Fagua G. 2007: 54 |
Canthon aequinoctiale
| Vaz-de-Mello F. Z. 1999: 449 |
Glaphyrocanthon proseni
| Halffter G. & Martinez A. 1977: 90 |
| Halffter G. & Matthews E. G. 1966: 43 |
Glaphyrocanthon (Glaphyrocanthon) proseni
| Vulcano M. A. & Pereira F. S. 1967: 561 |
| Martinez A. & Halffter G. & Halffter V. 1964: 3 |
| Vulcano M. A. & Pereira F. S. 1964: 663 |
| Martinez A. & Pereira F. S. 1956: 92 |
| Martinez A. 1950: 170 |
Glaphyrocanthon proseni Martínez, 1949a: 282
| Martinez A. 1949: 282 |
Canthon aequinoctialis
| Ratcliffe B. C. & Jameson M. L. & Figueroa L. & Cave R. D. & Paulsen M. J. & Cano E. B. & Beza-Beza C. & Jimenez-Ferbans L. & Reyes-Castillo P. 2015: 195 |
| Noriega J. A. 2012: 4 |
| Figueroa L. & Alvarado M. 2011: 210 |
| Carpio C. & Donoso D. A. & Ramon G. & Dangles O. 2009: 462 |
| Medina C. A. & Pulido L. A. 2009: 58 |
| Noriega J. A. & Cubillos A. M. & Castaneda C. & Sanchez A. M. 2008: 78 |
| Celi J. & Terneus E. & Torres J. & Ortega M. 2004: 42 |
| Noriega-Alvarado J. A. 2004: 40 |
| Medina C. A. & Lopera-Toro A. & Vitolo A. & Gill B. 2001: 135 |
| Escobar F. 2000: 206 |
| Escobar F. 2000: 114 |
| Balthasar V. 1951: 326 |
| Balthasar V. 1941: 341 |