
Microphallus
|
publication ID |
https://doi.org/ 10.5281/zenodo.210378 |
|
DOI |
https://doi.org/10.5281/zenodo.6166271 |
|
persistent identifier |
https://treatment.plazi.org/id/A872878D-FFF0-FFE1-FF73-3D54FE42FEB0 |
|
treatment provided by |
Plazi |
|
scientific name |
Microphallus |
| status |
|
Microphallus View in CoL sp. “livelyi”
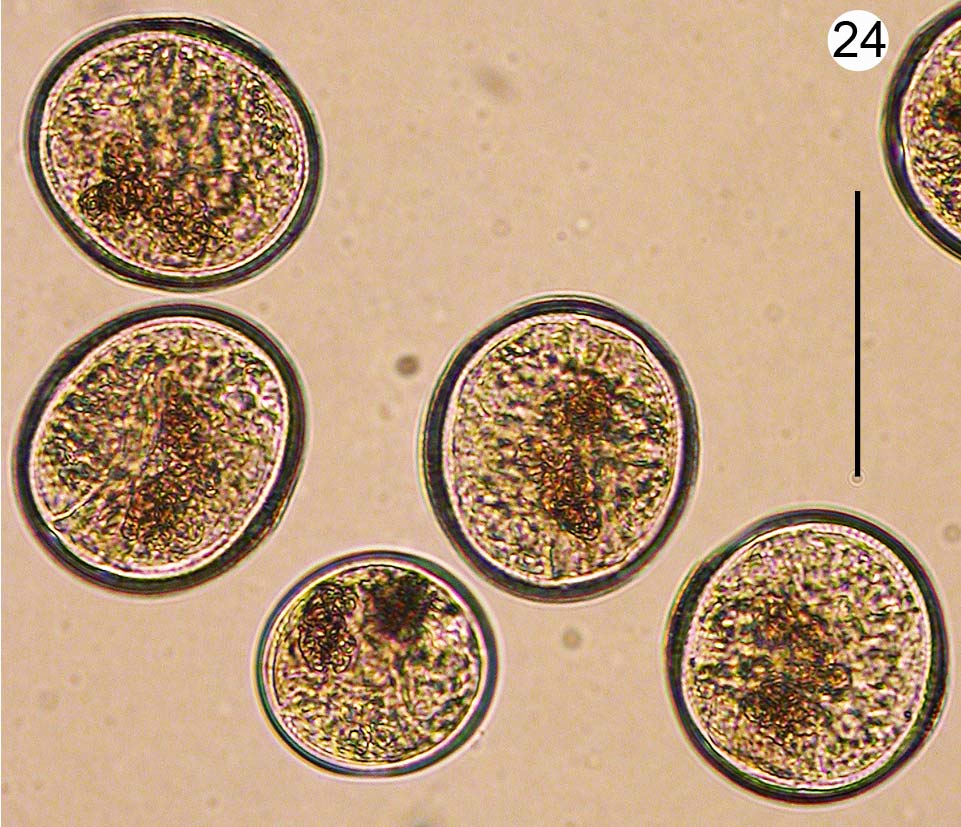
(13. Mili; Fig. 24 View FIGURE 24 )
Stages: embryonic and encysted metacercariae
Host: Potamopyrgus antipodarum
Site in host: embryonic and encysted metacercariae in gonad, digestive gland, and basal visceral mass
Voucher locality: Monowai Stream, South Island (-45.8067o lat, 167.5308o long)
Prevalence: 7% (n = 100)
Specimens deposited: USNPC #: 105687
Diagnosis: Parthenitae not-present. Cercariae not present. Initial infection with hundreds of germ balls and embryonic metacercariae that appear to develop directly to encysted metacercariae that fill most of distal visceral mass (gonad and digestive gland region) and much of basal visceral mass. Developed encysted metacercaria spherical to ovoid, ~100–120 diameter, with developing vitelline follicles apparent as two dark brown blotches.
Remarks: This species corresponds to Winterbourn’s Metacercaria A. I never detected daughter sporocysts for these infections, including in very early-stage infections. As mentioned by other workers (e.g., Krist & Lively 1998), developing infections progress from only having unencysted germinal balls and embryos, to having an increasing proportion of well-developed, fully-encysted metacercariae. All stages appear to develop freely in snail tissues or hemocoelomic spaces. Future histological work could confirm whether this species truly lacks daughter sporocysts. My colleagues and I are currently maintaining this species in the laboratory, and I am working to resolve its specific identity.
Microphallus View in CoL sp. “livelyi” possesses an evolutionarily modified lifecycle, using the first intermediate host as second intermediate host. Hence, it is a “trophically transmitted parasitic castrator,” sensu Lafferty and Kuris (2002). Adults use birds, particularly ducks, as final hosts ( Osnas & Lively 2011). Domestic mice serve as suitable laboratory hosts ( Lively & McKenzie 1991).
Other microphallids that have similar lifecycles wherein the first intermediate host individual also serves as second intermediate host. For such species, metacercaria embryos have variously been termed “blastocercariae” (e.g., Deblock 1974) or “cryptocercariae” ( Galaktionov & Dobrovolskij 2003). However, those embryos appear to always form within sporocysts. If Microphallus View in CoL sp. “livelyi” (or its ancestor) has indeed lost daughter sporocysts, it represents a variation on the theme characterizing these abbreviated live cycles.
Explaining my working name, this is the “ Microphallus View in CoL sp.” extensively worked on by Curt Lively and colleagues (e.g., Mark Dybdahl, Jukka Jokela, Ed Levri, Kayla King). This research, using this trematode-snail host interaction, includes classic empirical evidence for parasitism underlying the evolutionary maintenance of sexual reproduction, local adaptation of parasite host specificity, and coevolutionary cycling of hosts and parasites (e.g., Dybdahl & Lively 1998; Jokela & Lively 1995; Lively 1987; Lively 1989). Additionally, this trematode appears to adaptively modify the behavior of infected snails to increase trophic transmission to final host birds (e.g., see Levri 1999; Levri & Lively 1996). Infection also decreases growth in juvenile snails ( Krist & Lively 1998) and is also related to shell shape and defense morphology ( Levri et al. 2005). Please consult the literature for additional work on this system.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |