
Aphelochaeta clippertonensis, Blake, 2019
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4629.2.1 |
|
publication LSID |
lsid:zoobank.org:pub:89B34FE2-BCB0-4F13-B29C-3FDEABD8E15D |
|
DOI |
https://doi.org/10.5281/zenodo.5619229 |
|
persistent identifier |
https://treatment.plazi.org/id/CE40005E-0928-44AA-B800-57903EDFCB0F |
|
taxon LSID |
lsid:zoobank.org:act:CE40005E-0928-44AA-B800-57903EDFCB0F |
|
treatment provided by |
Plazi |
|
scientific name |
Aphelochaeta clippertonensis |
| status |
sp. nov. |
Aphelochaeta clippertonensis View in CoL new species
Figures 3–4 View FIGURE 3 View FIGURE 4
urn:lsid:zoobank.org:act:CE40005E-0928-44AA-B800-57903EDFCB0F
Tharyx sp. C: Wilson & Hessler 1987: Appendix E (in part).
Material examined. North Equatorial Pacific Ocean, abyssal plain, Clarion-Clipperton Fracture Zone, NOAA BIE Project site, Sta. DDT-9-93, veg. 13, 5– 10 cm fraction, 03 Sep 1993, 12°56.280′N, 128°35.440′W, 4860 m, holotype ( USNM 1557530 View Materials ) GoogleMaps ; Sta. DDT 2-93, veg 20, 2– 5 cm fraction, 10 Aug 1993, 12°56.166′N, 128°35.520′W, 4869 m, 1 specimen ( USNM 1557532 View Materials ) GoogleMaps ; Sta. DDT 5-93, veg 14, 2– 5 cm fraction, 13 Aug 1993, 12°56.566′N, 128°35.408′W, 4870 m, 1 paratype ( USNM 1557533 View Materials ) GoogleMaps ; Sta. DDT 6-93, veg 13, 0–2 cm fraction, 01 Sep 1993, 12°55.780′N, 128°35.881′W, 4858 m, 1 specimen ( USNM 1557534 View Materials ) GoogleMaps ; Sta. DDT-8-93, veg. 24, 0–2 cm fraction, 02 Sep 1993, 12°55.633′N, 128°36.011′W, 4843 m, 1 paratype ( USNM 1557531 View Materials ).— ECHO I GoogleMaps , DOMES Site C, R/ V Melville cruise, coll. R. Hessler, 0.25 m 2 Sandia box core, Sta. H 350, 1– 5 cm fraction, 14 Jun 1983, 14°38.1226′N, 125°26.8208′W, 4506 m, 1 paratype ( LACM-AHF Poly 11259) GoogleMaps .
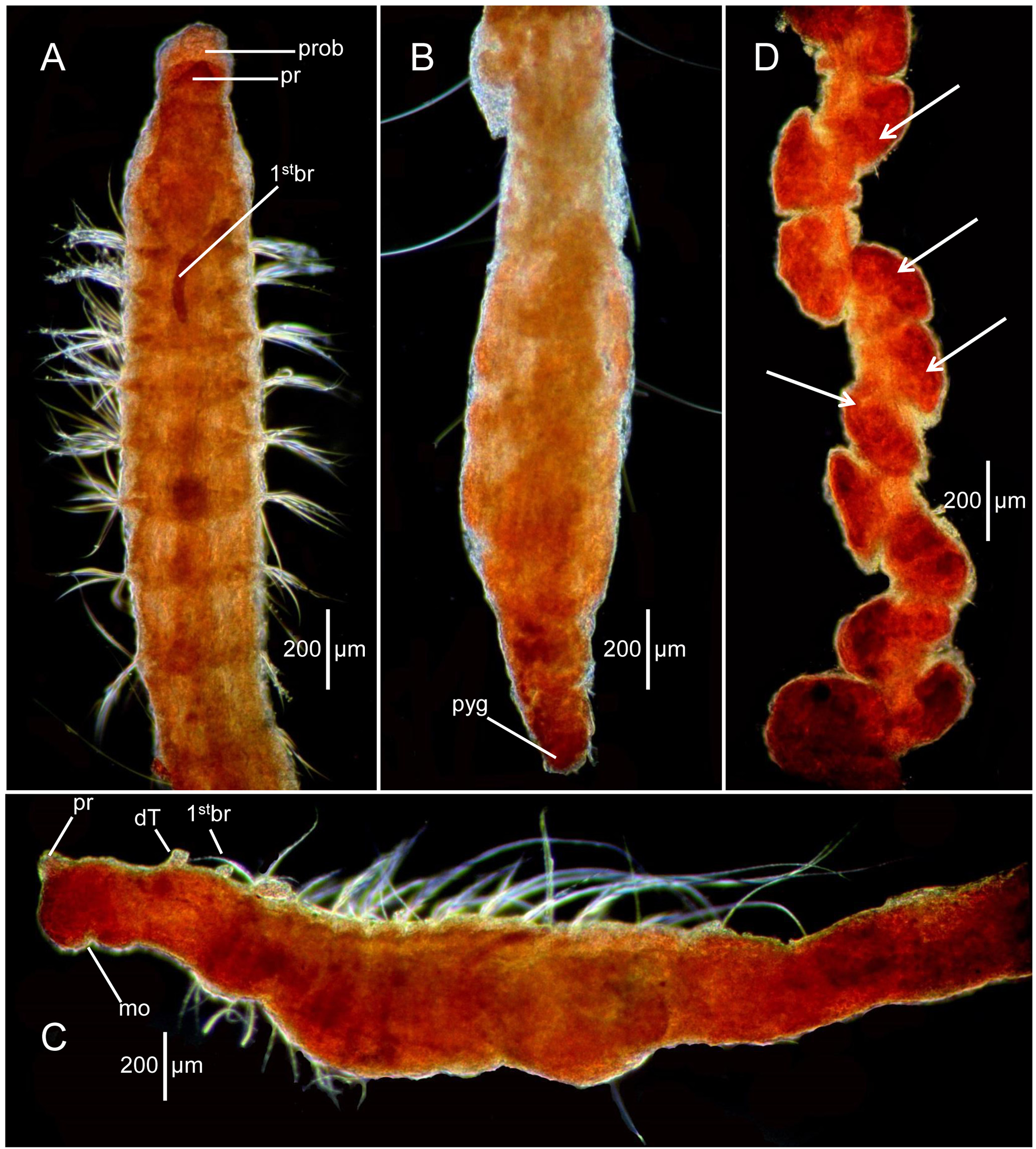
Description. A small, elongate, threadlike species. Holotype complete, 4.0 mm long, 0.3 mm wide across anterior segments with 20 setigers; paratype from Sta. DDT-5-93 complete, 1.83 mm long, 0.213 mm wide with 16 setigers. Body slightly expanded in anterior and far posterior segments ( Figs. 3 View FIGURE 3 A–B, 4A–C), but generally consistently narrow along entire body. Individual segments visibly separated from one another by setal fascicles; segmental furrows poorly developed ( Fig. 3 View FIGURE 3 A–B); no moniliform segments. Parapodial shoulders not apparent in anterior segments or elsewhere along body. First 6–8 segments narrow, about three times wider than long ( Figs. 3A View FIGURE 3 , 4A, C View FIGURE 4 ); middle body segments longer, about 1.5 times as wide as long ( Fig. 4C View FIGURE 4 ); posterior segments longer, about 1.8 times as wide as long ( Figs. 3B View FIGURE 3 , 4B View FIGURE 4 . Dorsal and ventral grooves absent. Paratype from Sta. H350 with posterior segments bearing elongate embryos, two per segment, each approximately 85 µm long and 35 µm wide ( Fig. 4D View FIGURE 4 ). Each embryo composed of numerous cells, but not differentiated. Color in alcohol opaque white.
Pre-setiger region narrow, tapering from setiger 1 to tip of prostomium, about 1.5 times longer than wide ( Figs. 3A View FIGURE 3 , 4A View FIGURE 4 ). Prostomium triangular, tapering to pointed apex; eyespots absent; nuchal organs not observed; proboscis with thin papillated epidermis, everted on holotype ( Figs. 3A View FIGURE 3 , 4A View FIGURE 4 ). Peristomium with three weakly developed annular rings only evident laterally by shallow furrows; dorsal surface smooth with no separate dorsal crest ( Fig. 3A View FIGURE 3 ). Dorsal tentacles present on anterior border of third peristomial ring ( Figs. 3A View FIGURE 3 , 4 View FIGURE 4 A–B); first pair of branchiae present posterior to tentacles at anterior border with setiger 1 ( Figs. 3A View FIGURE 3 , A–B). Second pair of branchiae on setiger 1 dorsal to notosetae; subsequent branchiae in similar location on following segments ( Fig. 3A View FIGURE 3 ). Most branchiae missing but evident as stubs or scars through about setiger 10.
Parapodia poorly developed throughout, with podial lobes reduced and with setae emerging directly from body wall. All setae thin capillaries. Notosetae 7–8 per fascicle on first 5–6 setigers, then reduced to 3–5 per fascicle over next 5–6 setigers; posterior setigers with 1–2 long natatory-like capillaries ( Figs. 3B View FIGURE 3 , 4C View FIGURE 4 ); neurosetae of anterior setigers similar to notosetae, numbering 6–8 per fascicle, then reduced to 3–4 in following segments; posterior segments with 1–2 capillaries where long notosetae occur. Individual capillaries thin, with no marginal fibrils evident in light microscopy.
Posterior end terminating in simple conical-shaped pygidium bearing numerous minute papillae ( Figs. 3B View FIGURE 3 , 4C View FIGURE 4 ).
Methyl Green stain. No pattern.
Etymology. The name clippertonensis is taken from the second part of the name of the Clarion-Clipperton Facture Zone where the species was collected.
Remarks. Aphelochaeta clippertonensis n. sp. is one of several small bitentaculate cirratulids from abyssal sediments in the Clarion-Clipperton Fracture Zone with a slender threadlike body. Among species of Aphelochaeta identified in this project, A. clippertonensis n. sp. is the only one to have reduced segmental furrows that result in the individual segments being defined only by the location of setal fascicles. Other species in this study have individual segments well defined with segmental furrows and distinctly separate from one another along the body. An exception is the paratype from Sta. H350 that has elongate brooding embryos within segments in the posterior half of the body producing segments that appear swollen, lumpy, and rounded ( Fig. 4D View FIGURE 4 ). There appear to be two embryos per segment that are similar in appearance to eggs and embryos observed for Tharyx moniliformis Blake, 2018 from slope and abyssal depths in the Weddell Sea, Antarctica. These observations suggest that deep-sea cirratulids have evolved viviparity or other means of brooding as part of their life cycles.
Distribution. Abyssal Pacific Ocean, 4506–4870 m.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |