
Psallus Fieber, 1858
|
publication ID |
https://doi.org/10.11646/zootaxa.5094.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:72F6E1D9-E9E4-41F6-9AC0-97A249F94E68 |
|
DOI |
https://doi.org/10.5281/zenodo.6302537 |
|
persistent identifier |
https://treatment.plazi.org/id/AC7FEA3B-0816-FFF5-FF78-FD7DFA4975B8 |
|
treatment provided by |
Plazi |
|
scientific name |
Psallus Fieber, 1858 |
| status |
|
Psallus Fieber, 1858 View in CoL View at ENA
Discussion. The phyline genus Psallus was established by Fieber (1858) [ type species: P. haematodes (Gmelin, 1790) , a Euro-Siberian element] and is known to comprise approximately 130 species in the Holarctic Region (9 Nearctic species were introduced from Europe, cf. Wheeler & Henry, 1992; Schuh, 2002 – 2013; Wyniger, 2004). As pointed out by several authors (e.g., Yasunaga & Vinokurov; Wyniger, 2004; Yasunaga, 2010), the monophyly of Psallus cannot be elucidated by its current context, sometimes being termed a “garbage group”. It is usually difficult to distinguish species of Psallus from those exhibiting similar appearance in some other genera by the external characters alone.
Six valid subgenera have been proposed for classification of Psallus (cf. Aukema, 2018): Apocremnus Fieber, 1858 , Calopsallus Yasunaga and Vinokurov, 2000 , Hylopsallus Wagner, 1952 , Mesopsallus Wagner, 1970 , Phylidea Reuter, 1899 and Pityopsallus Wagner , in addition to the nominotypical subenus. Some of these subgenera may require generic status, like as Pityopsallus that is treated as a full genus in the present work. Although Wyniger (2004) did not use any subgeneric rank for her classification of Psallus , the subgenera Apocremnus , Calopsallus , Hylopsallus Phylidea and Psallus s. str. are considered to constitute monophyletic groups, on the basis of available external morphology and male genitalic structures (cf. Duwal et al., 2012). However, definitive treatments of the monophyly and relationships are beyond the scope of this study, and eventually a comprehensive phylogenetic analysis and subsequent revision are required to redefine the relevant taxa.
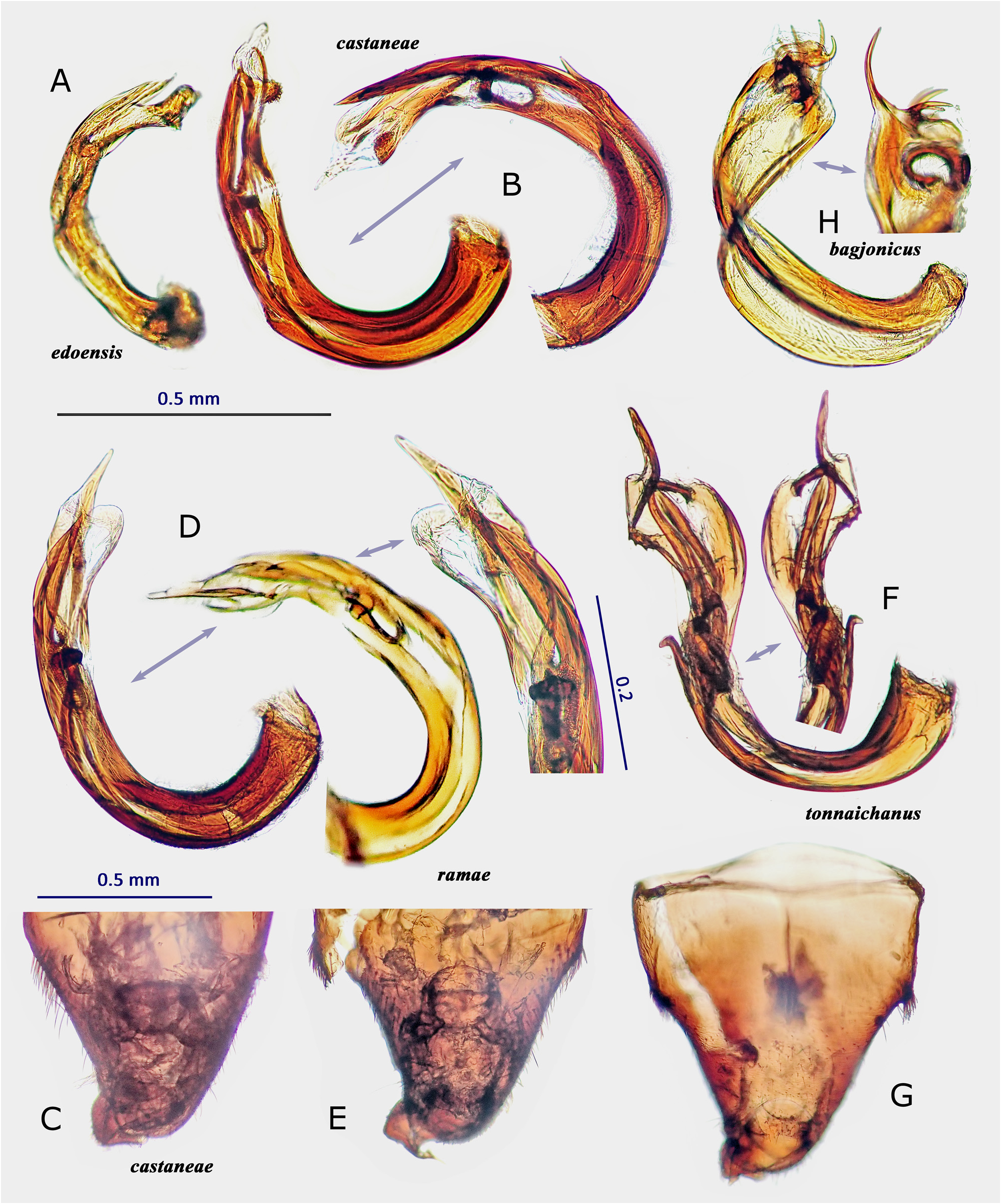
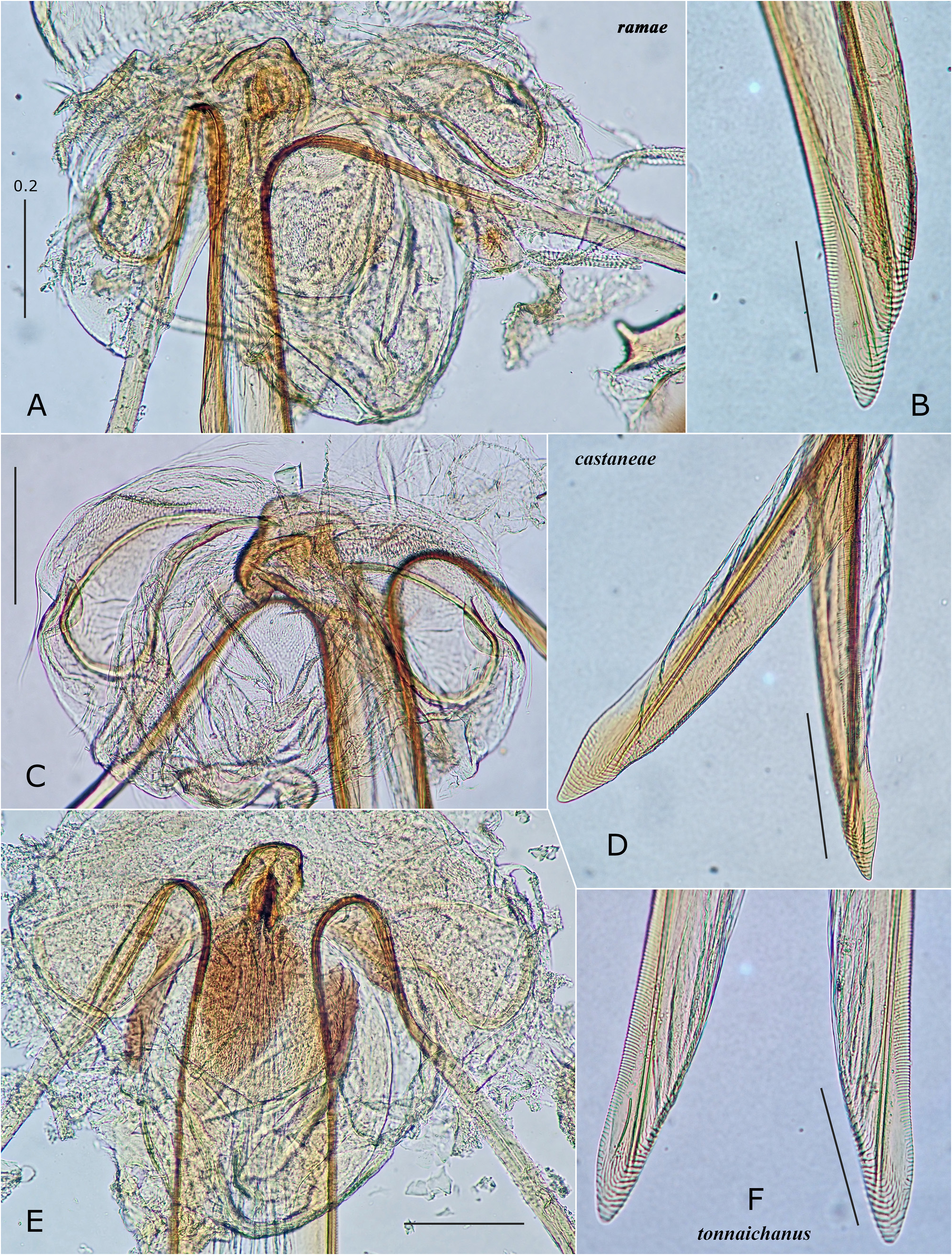
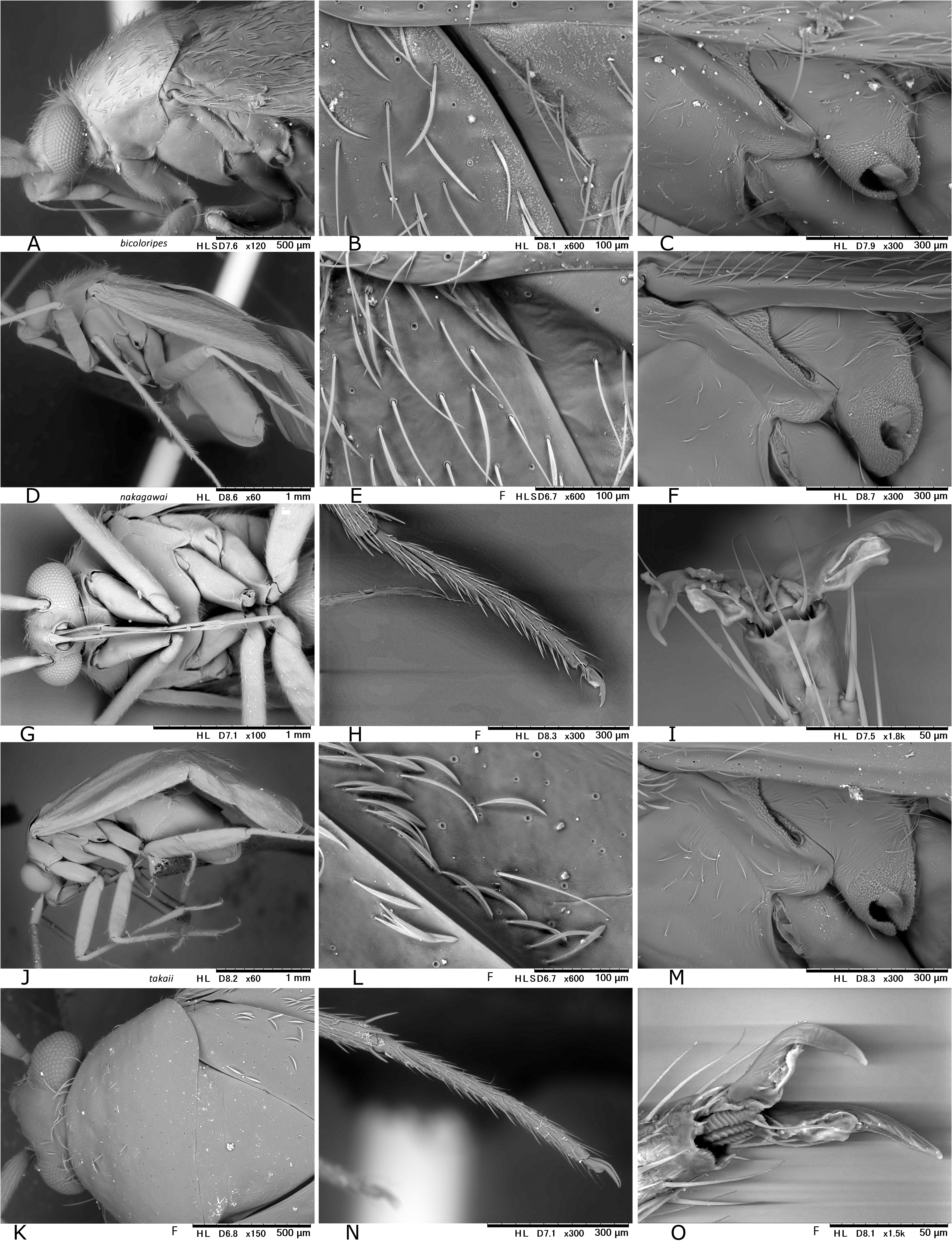
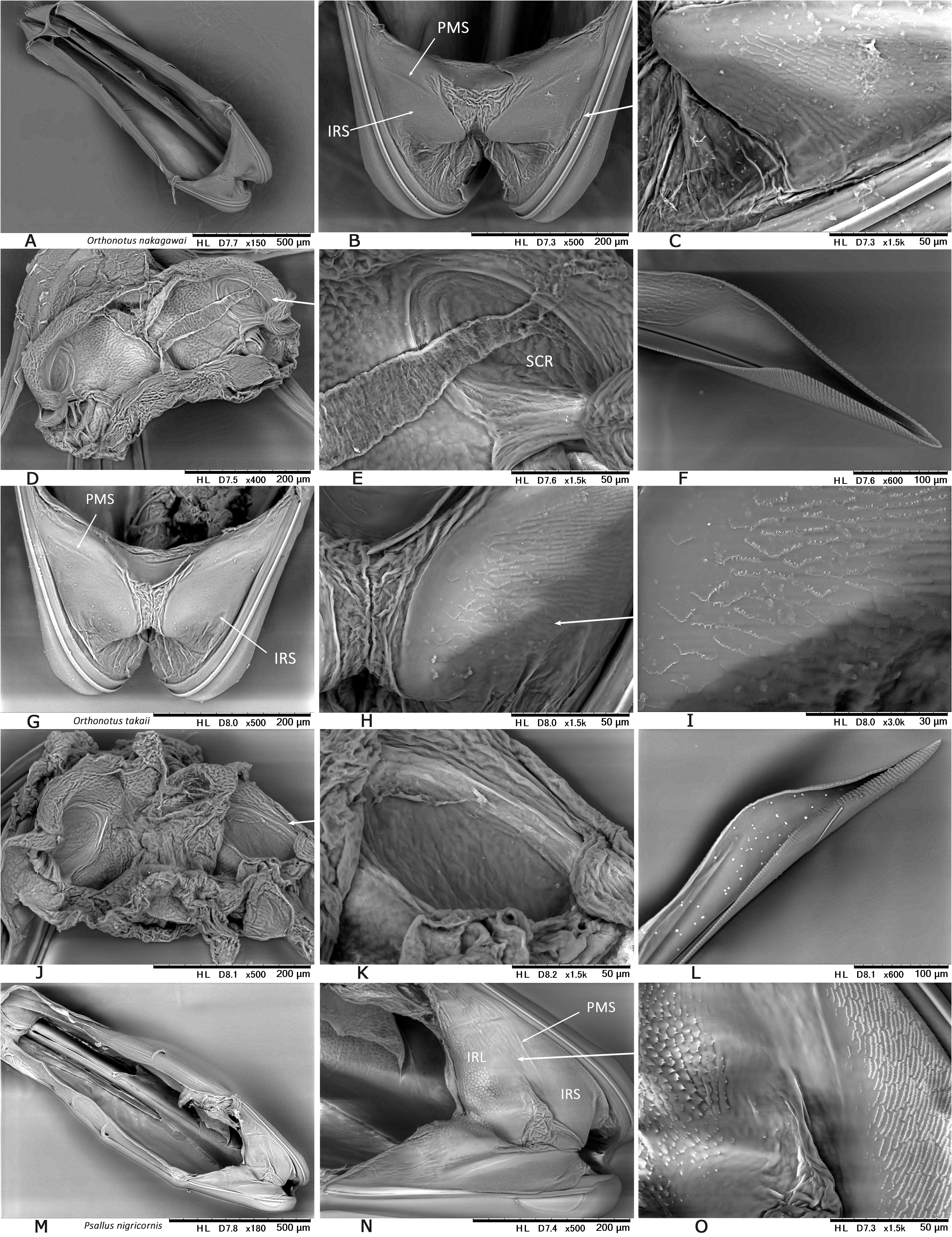
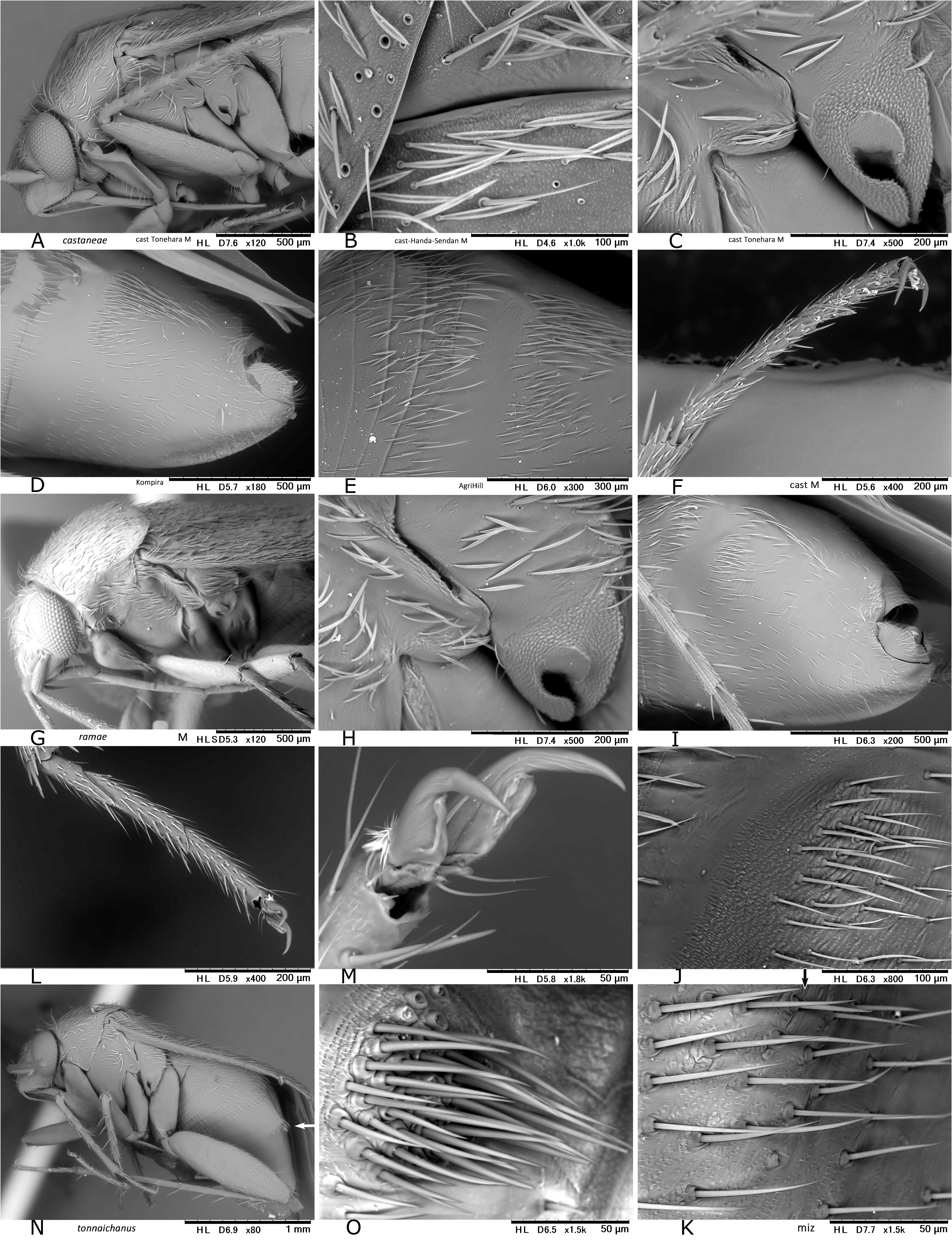
In Japan, 19 species of Psallus were previously recorded ( Yasunaga & Vinokurov, 2000; Yasunaga, 2001d). Subsequent efforts by the author and colleagues have recognized an undescribed species mentioned below, belonging to the subgenus Hylopsallus . The ‘true’ members of Psallus s. str. (the type species P. haematodes and its close relatives) share the following characters: thoracic pleura without silvery seta (except for P. bagjonicus Josifov, 1983 and P. buddha Yasunaga, 2010 with relatively denser silvery setae); blunt-tipped phallotheca, flattened, medially twisted vesica with apically situated, small-sized secondary gonopore and an elongate apical blade (cf. Fig. 14H View FIGURE 14 ) that is usually notched laterally ( Yasunaga & Vinokurov, 2000); posterior wall with noticeable spinulate scalelike microstructures on (undifferentiated) interramal lobe and narrow posterior margin sclerite ( Fig. 25M–O View FIGURE 25 ); and rounded, smaller sclerotized rings (cf. Yasunaga, 2010, Fig. 23 View FIGURE 23 ). On the other hand, species included in Hylopsallus have the silvery setae on the thoracic pleura ( Figs. 13C, F View FIGURE 13 ; 29A, D, G, L View FIGURE 29 ), sharply pointed phallotheca (e.g., Fig. 31B, E View FIGURE 31 ), stout, relatively tubular vesica with 3 distinct processes ( Fig. 31F, I, O–M View FIGURE 31 ) and thick-rimmed, elongateovoid secondary gonopore (e.g., Figs. 14A–B View FIGURE 14 ), relatively smooth interramal lobe (e.g., Fig. 32C–D View FIGURE 32 ), and wide sclerotized rings that are often asymmetrical to each other (cf. Fig. 15C, E View FIGURE 15 ; Wyniger, 2004). Psallus castaneae Josifov, 1983 that has been placed in the subgenus Phylidea , is herein regarded as a member of Hylopsallus , because of sharing the above-mentioned characters. Accordingly, five species of Psallus (Hylopsallus) are now recognized in Japan: P. (H.) aterrimus Yasunaga & Vinokurov, 2000 ; P. (H.) castaneae Josifov, 1983 ; P. (H.) edoensis Yasunaga & Vinokurov, 2020 ; P. (H.) ramae n. sp.; and P. (H.) tonnaichanus Muramoto, 1973 .
The following key will distinguish Japanese members of the subgenus Hylopsallus including a new species as well as a superficially very similar congener, P. bagjonicus Josifov, 1983 (for identification of all other Japanese Psallus species , see Yasunaga & Vinokurov, 2000).
Key to Japanese species of Hylopsallus and Psallus bagjonicus
1. Antennal segment I almost entirely fuscous................................................................ 2
– Antennal segment I entirely yellow or pale brown, or its base narrowly infuscate................................... 3
2. Metafemur uniformly dark brown; male pygophore (genital segment) only with simple setae, lacking silvery seta ( Fig. 29B View FIGURE 29 )........................................................................................ P. (H.) aterrimus View in CoL
– Ventral surface of metafemur chestnut or reddish brown, with large, fuscous spots; pygophore with silvery setae ( Fig. 29H View FIGURE 29 )........................................................................................ P. (P.) bagjonicus View in CoL
3. Base of antennal segment I usually darkened; protibia with dark spots at bases of spines; male genital segment with distinct ventral mesal keel from middle part to apex ( Fig. 30D View FIGURE 30 )....................................................... 4
– Antennal segment I almost entirely pale, or its extreme base faintly obscured; bases of protibial spines lacking dark spots; ventral mesal keel on male genital segment reduced, forming a narrow carina at apical 1/2–2/3 of segment ( Fig. 29M View FIGURE 29 )..... 5
4. Width of pronotum greater than length of metafemur in female; metatibial dark spots smaller, separated from each other; setae on male genital segment not aggregated or clustered at left-dorsolateral base ( Fig. 30D–E View FIGURE 30 ).............. P. (H.) castaneae View in CoL
– Width of pronotum slightly less than length of metafemur in female; metatibial dark spots often enlarged (some are fused as in Fig. 13D–E View FIGURE 13 ); setae on male genital segment somewhat aggregated at left-lateral base ( Fig. 30J View FIGURE 30 )........ P. (H.) ramae n. sp.
5. Hemelytra blackish brown; male genital segment with a pair of clustered stiff setae ( Fig. 30N–O View FIGURE 30 )....... P. (H.) tonnaichanus View in CoL
– Hemelytra chestnut brown; setae on male genital segment not aggregated ( Fig. 31D View FIGURE 31 ); spots at bases of metatibial spines small, obscure................................................................................. P. (H.) edoensis View in CoL




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Tribe |
Phylini |
|
SubTribe |
Phylina |