
Chrysopidae, Schneider, 1851
|
publication ID |
https://doi.org/10.11646/zootaxa.5133.3.1 |
|
publication LSID |
lsid:zoobank.org:pub:16A54EFD-028D-42FF-BD62-B8D35B7BA4DC |
|
DOI |
https://doi.org/10.5281/zenodo.6522485 |
|
persistent identifier |
https://treatment.plazi.org/id/AD76878E-FFC9-FFCA-E292-A308CBDD724F |
|
treatment provided by |
Plazi |
|
scientific name |
Chrysopidae |
| status |
|
Eocene Chrysopidae View in CoL View at ENA and the Florissant chrysopid assemblage
The oldest known Cenozoic species of Chrysopidae is Sinonothochrysa zhangi Huang et Nel in Huang et al. (2021) from the Paleocene of southern China ( Huang et al. 2021). Its venation is characteristic of the subfamily Nothochrysinae (see its diagnosis of Makarkin & Archibald 2013), including the long Sc, the short intramedian cell, and the proximal position of the crossvein 2m-cu. It is interesting that the hind wing MA in this oldest Nothochrysinae is apomorphic, fused for some distance with RP as in most other genera of the subfamily.
The only other Eocene Chrysopidae known outside of Europe and North America is Asiachrysa tadushiella Makarkin, 2014 from the Tadushi Formation of Pacific coastal Russia, estimated to be early Eocene ( Popov & Grebennikov 2001; Makarkin 2014). As mentioned above, the forewing venation of this genus is most similar to that of Cimbrochrysa .
All other Eocene Chrysopidae are from North America and Europe.
Three monotypic genera of Nothochrysinae ( Danochrysa , Cimbrochrysa , and Stephenbrooksia ) and one of Limaiinae ( Protochrysa Willmann et Brooks, 1991 ) are recorded from the early Ypresian (earliest Eocene) of Denmark (Mo-clay; all probably from the Fur Formation) ( Schlüter 1982; Willmann & Brooks 1991; Willmann 1993). The subfamily affinity of Brooksiochrysa Rust, 1999 ( nomen nudum) is unclear (Rust 1999: Pl. 23, Fig. f). The forewing of its only species has three gradate series, while two in other Mo-clay Chrysopidae .
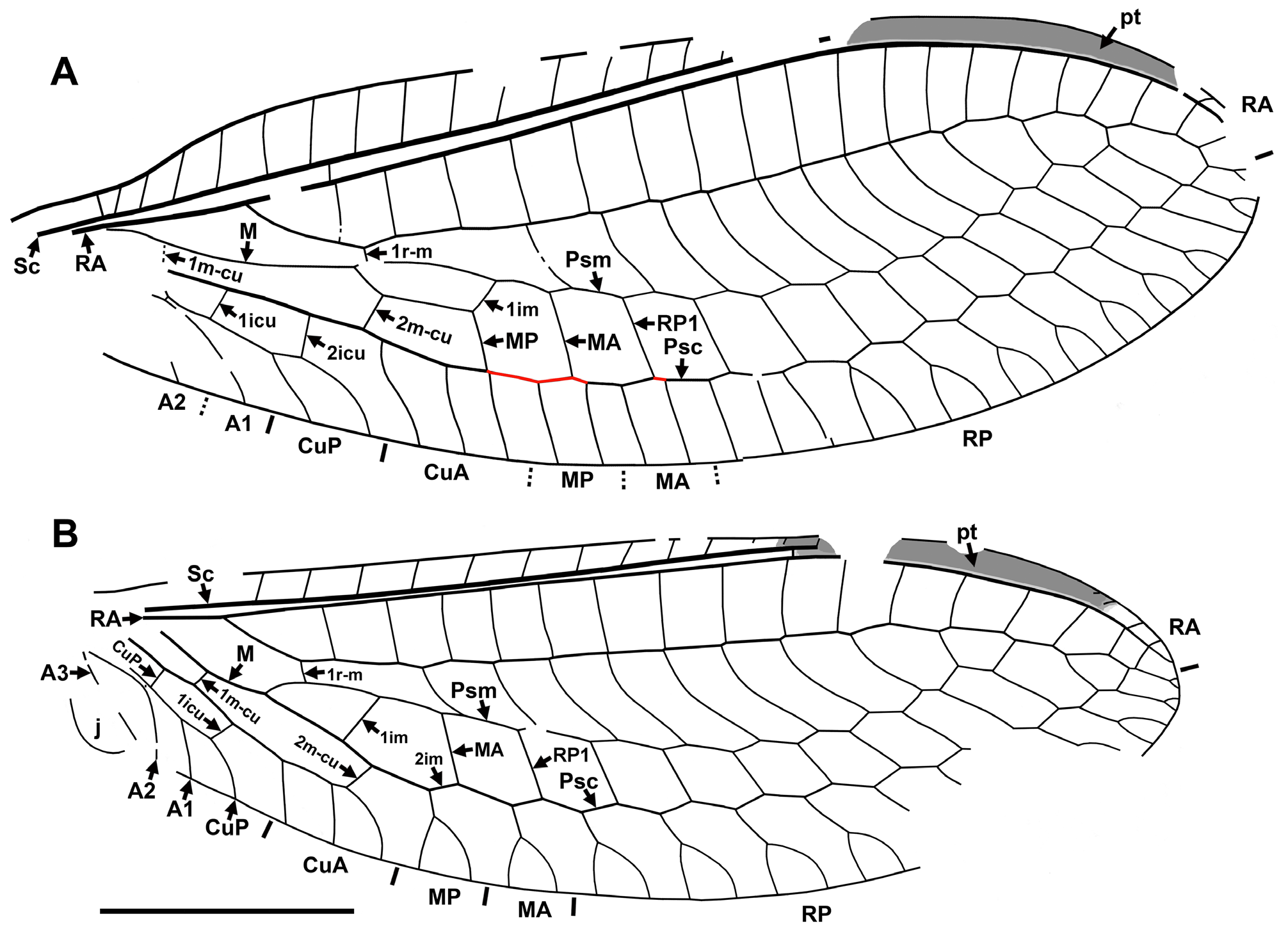
The genus Stephenbrooksia is similar to the North American Archaeochrysa (and extant monotypic Leptochrysa ) by the presence of the basal crossvein 1r-m, a plesiomorphic condition ( Fig. 2 View FIGURE 2 ). In other genera of Nothochrysinae (fossil and extant), this crossvein is absent as RP and MA are fused for some distance, an apomorphic condition ( Fig. 28 View FIGURE 28 ). We agree with Adams (1967) that genera possessing this plesiomorphic condition are more ‘archaic’ in the subfamily; the majority of the Mesozoic Limaiinae possess this character state.
The venation of Cimbrochrysa here first recorded from North America is generally similar to that of the Florissant genera Palaeochrysa and Lithochrysa , differing mainly by the shape of its intramedian cell. The forewing venation of the genus Danochrysa is also somewhat similar to that of Cimbrochrysa , but differs in that all anal veins and both branches of CuP are forked. These character states are unique in the subfamily.
Five genera and at least six species of Nothochrysinae ( Archaeochrysa , Adamsochrysa , Lithochrysa , Okanaganochrysa , Pseudochrysopa ), and the Limaiinae genus Protochrysa (one species) are known from the early Eocene Okanagan Highlands of far western North America ( Makarkin & Archibald 2013; Archibald & Makarkin 2015, 2017). These chrysopids are most morphologically diverse compared to other Cenozoic assemblages. Adamsochrysa and Pseudochrysopa , most characteristic of that assemblage, have venation very dissimilar to that of other Eocene chrysopids. The wings of Adamsochrysa are the most densely veined, with four or five gradate series of crossveins, while those of Pseudochrysopa have some of the simplest venation of chrysopids in which Psm and Psc are absent.
Six genera and 13 species are known from Florissant: Archaeochrysa (three species), Cimbrochrysa (two species), Dyspetochrysa (one species), Lithochrysa (four species), Palaeochrysa (one species) and Tribochrysa (two species). The validity of the genus Dyspetochrysa is not clear (see above). This assemblage is less morphologically diverse than those of the early Eocene. Palaeochrysa is probably the most derived among these genera with hind venation similar to that of Chrysopinae (above), the most derived subfamily.
The Chrysopidae View in CoL of European Baltic amber include the chrysopine genus Pseudosencera Makarkin et al. 2018 (three known specimens) and fragmentarily preserved specimens of Nothochrysinae most similar to Nothochrysa View in CoL (one known specimen) ( Makarkin et al. 2018; pers. obs.). Therefore, the taxonomic compositions of these late Eocene localities are very different. This is also characteristic of their raphidiopteran assemblages (see Archibald & Makarkin 2021).
Chrysopidae View in CoL from other late Eocene European localities show the same pattern. It appears that Chrysopinae were the dominant chrysopids in the late Eocene of Europe. Green lacewings of the latest Eocene Bembridge Marls ( UK) are represented by two specimens. One is so fragmentary that any conclusions are impossible ( Chrysopidae View in CoL Species A: Jarzembowski 1980: Fig. 42); the hind wing venation of another ( Chrysopidae View in CoL Genus undetermined species B: Nel & Jarzembowski 2019: Figs 2c View FIGURE 2 , 3 View FIGURE 3 ) is similar to that of the chrysopine Pseudosencera baltica Makarkin et al. 2018 . Paleochrysopa Séméria et Nel, 1990 View in CoL from Monteils ( France) belongs probably to Chrysopinae ( Séméria & Nel 1990) .
The Eocene chrysopid genera Protochrysa (Limaiinae) View in CoL and Cimbrochrysa (Nothochrysine) View in CoL have been found in both Europe and North America. Other shared neuropteran genera include the polystoechotid Palaeopsychops Andersen, 2001 View in CoL ( Archibald & Makarkin 2006); the hemerobiid Proneuronema Makarkin et al., 2016 View in CoL ( Makarkin et al. 2016); and the osmylid Osmylidia Cockerell, 1908b View in CoL ( Makarkin et al. 2021). These support evidence from other insect orders for Eocene transatlantic faunal connections ( e.g., Archibald et al. 2011 and references therein, and closely related odonates: Petrulevičius et al. 2007; Archibald & Cannings 2019; Simonsen et al. 2022).
Key to species of Florissant Chrysopidae
( Lithochrysa concinnula is not included, as its description is incomplete and the holotype is obscured by Canadian balsam)
1. One crossvein present between RA and RP proximad 1r-m in forewing. M and RP in hind wing not fused ( Archaeochrysa View in CoL ). 2
- Crossveins absent between RA and RP proximad 1r-m in forewing. M and RP in hind wing fused for some distance...... 4
2. Anterior and posterior sides of im (MA, MP) in forewing converging basally at low angle.......... Archaeochrysa fracta View in CoL
- Anterior and posterior sides of im in forewing parallel and converging basally at steeper angle........................ 3
3. Forewing costal margin convex through costal space.............................. Archaeochrysa cockerelli sp. nov.
- Forewing costal margin of forewing not convex in middle of costal space................... Archaeochrysa paranervis View in CoL
4. Forewing RP1 originating distad two proximal crossveins between RA and RP................. Dyspetochrysa vetuscula View in CoL
- Forewing RP1 originating distad at most one proximal crossvein between RA and RP............................... 5
5. Inner gradate series of crossveins in fore- and hind wings arranged in a more or less smooth line...................... 6
- Inner gradate series of crossveins in fore- and hind wings arranged in strongly broken line (shifted anteriad at termination of Psm).............................................................................................. 10
6. In forewing, im broadly pentagonal; 2m-cu located at proximal 1/3 to 1/2 of im length.............................. 7
- In forewing, im rather narrow, elongate; 2m-cu located at proximal 1/4 of im length................................ 8
7. Forewing RP with <10 branches; 1r-m in more distal position within im (at proximal one-third length).......................................................................................... Cimbrochrysa americana sp. nov.
- Forewing RP with ≥10 branches; 1r-m in more proximal position (at proximal one-fifth to one-fourteenth length)..................................................................................... Cimbrochrysa major sp. nov.
8. In forewing, cells formed by branches of MA, MP, CuA between Psc and hind margin broad, 1.20–1.25 as long as wide...................................................................................... Lithochrysa wickhami View in CoL
- In forewing, cells formed by branches of MA, MP, CuA between Psc and hind margin narrow, 1.5–2.0 as long as wide.... 9
9. Forewing RP with 12 branches; 1r-m in more proximal location ( ca. at level of 2m-cu)........ Lithochrysa meyeri sp. nov.
- Forewing RP with ≤10 branches; 1r-m in more distal location (clearly distad 2m-cu)............. Lithochrysa ferruginea
10. In forewing, Psm formed by five branches of RP and nearly straight, very slightly zigzagged......... Palaeochrysa stricta View in CoL
- In forewing, Psm formed by two-three branches of RP and strongly zigzagged................................... 11
11. Forewing subcostal veinlets more closely spaced (seven between origin of RP and third ra-rp); RP with 10 branches........................................................................................ Tribochrysa inaequalis
- Forewing subcostal veinlets more closely spaced (five– six between origin of RP and third ra-rp); RP with 8 branches.......................................................................................... Tribochrysa firmata View in CoL


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Nothochrysinae |
|
Genus |
Chrysopidae
| Makarkin, Vladimir N., Antell, Gwen S. & Archibald, S. Bruce 2022 |
Pseudosencera
| Makarkin 2018 |
Pseudosencera baltica
| Makarkin 2018 |
Proneuronema
| Makarkin 2016 |
Palaeopsychops
| Andersen 2001 |
Paleochrysopa Séméria et Nel, 1990
| Semeria et Nel 1990 |
Nothochrysinae
| Navas 1910 |
Osmylidia
| Cockerell 1908 |
Nothochrysa
| McLachlan 1868 |
Chrysopidae
| Schneider 1851 |
Chrysopidae
| Schneider 1851 |
Chrysopidae
| Schneider 1851 |
Chrysopidae
| Schneider 1851 |