
Hincksina synchysia, Berning & Spencer Jones & Vieira, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5081.3.2 |
|
publication LSID |
lsid:zoobank.org:pub:489D7694-F51B-4582-BE02-9C9EBD1F150A |
|
DOI |
https://doi.org/10.5281/zenodo.5776657 |
|
persistent identifier |
https://treatment.plazi.org/id/DFE9C22D-A11B-4010-A70B-89A6D4933FDE |
|
taxon LSID |
lsid:zoobank.org:act:DFE9C22D-A11B-4010-A70B-89A6D4933FDE |
|
treatment provided by |
Plazi |
|
scientific name |
Hincksina synchysia |
| status |
sp. nov. |
Hincksina synchysia View in CoL n. sp.
( Fig. 2 View FIGURE 2 )
urn:lsid:zoobank.org:act:DFE9C22D-A11B-4010-A70B-89A6D4933FDE
Hincksina flustroides View in CoL f. crassispinata Gautier, 1962: 50, fig. 8.
Hincksina flustroides Hincks View in CoL : Harmelin 1988 (part): pl. 1, fig. 4, not fig. 5; Zabala & Maluquer 1988 (part): 80, text-fig. 86, pl. 3, fig. A, not text-fig. 87; Chimenz Gusso et al. 2014: 103, fig. 40a, b.
Material examined. Holotype: MNHN-IB-2017-782, 07/10/1959, Punta Carega , Portofino (Liguria, Italy), 30–40 m, figured by Gautier (1962, fig. 8), one colony on bivalve shell . Paratype: MNHN-IB-2008-10579, locality information as lectotype, one large colony on bivalve shell, http://coldb.mnhn.fr/catalognumber/mnhn/ib/2008-10579.
Etymology. From the Greek sýnchysi (= confusion), alluding to the confusing history of the species.
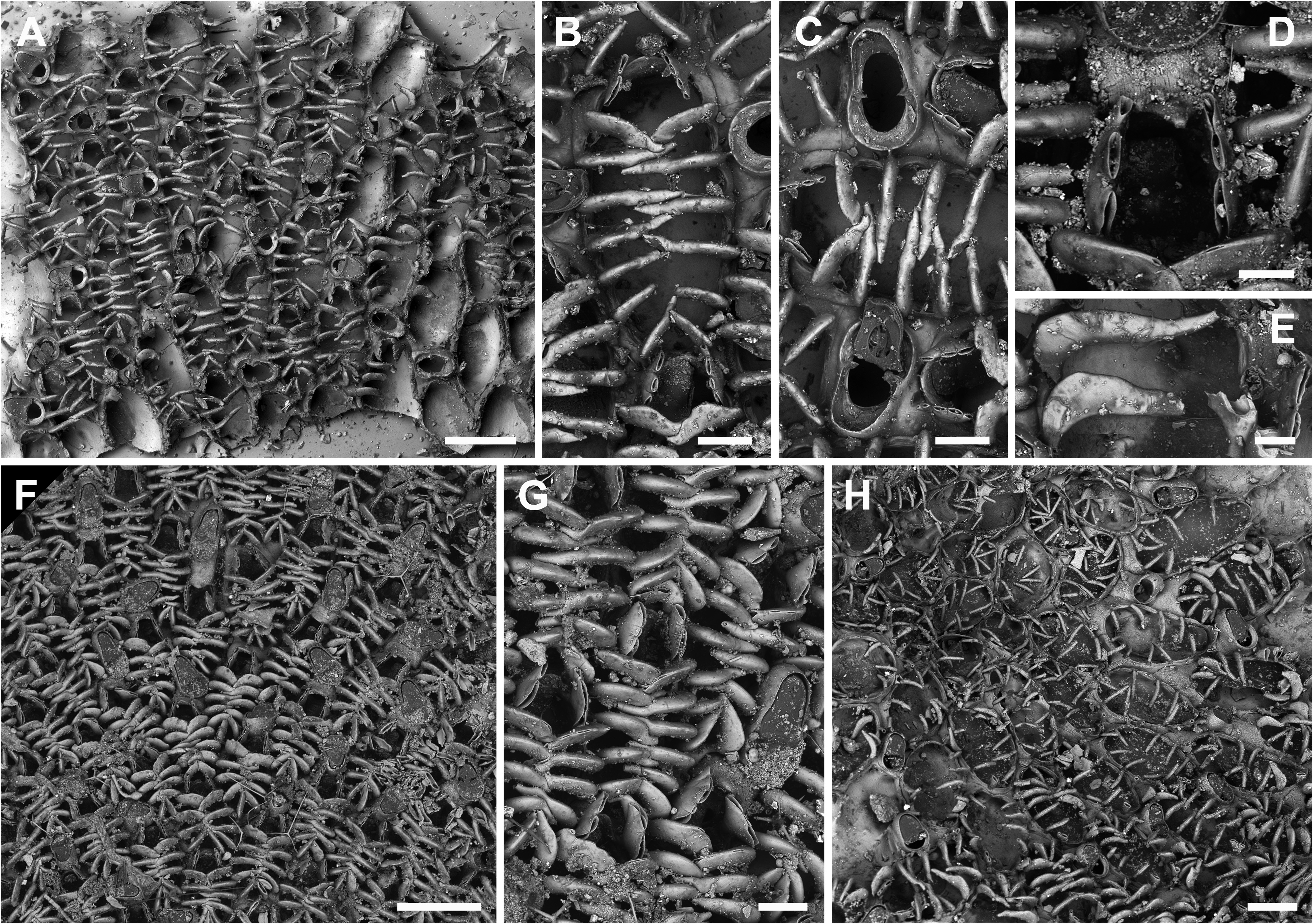
Description. Colony encrusting, unilaminar, multiserial. Autozooids oval to rectangular (ZL 474±47, 406–596, 20; ZW 288±26, 255–356, 20), arranged in irregular series, zooids separated by shallow grooves ( Fig. 2A–D View FIGURE 2 ), whitish translucent when dried. Vertical walls with two or more uniporous pore plates per neighbouring zooid. Development of gymnocyst reduced to proximal region and distolateral corner(s), cryptocyst a narrow band with a nodular surface, opesia extensive (OpL 372±30, 318–445, 20; OpW 205±15, 180–230, 20) ( Fig. 2A, C, D View FIGURE 2 ). Opercular region laterally with a pair of cylindrical to slightly flattened oral spines, more or less directing terminally ( Fig. 2C–E View FIGURE 2 ); frontal membrane in mature zooids overarched by 6–9 cylindrical mural spines that slightly thin towards their tips while not reaching the zooidal mid-line, proximal spines shorter ( Fig. 2A–D View FIGURE 2 ); all spines apparently unjointed at their base.
Ovicell usually endozooidal in distal avicularium but often in distal autozooid ( Fig. 2B, D View FIGURE 2 ), ooecium continuous with the gymnocyst of the avicularian or autozooidal cystid forming a short but broad hemispherical cap (OvL 89±13, 67–115, 20; OvW 182±17, 154–213, 20), entire proximal margin slightly raised but without a distinctive peak ( Fig. 2B, D, E View FIGURE 2 ).
Avicularia interzooidal, distal to many zooids ( Fig. 3A–D View FIGURE 3 ), particularly abundant in fertile colony regions, usually pointing distolaterally but often laterally, distally or even proximally, distinctly longer than wide and oval in outline, widest proximal to condyles, (AL 171±19, 140–213, 20; AW 112±7, 101–128, 20); rostrum semielliptical or rounded-triangular, occasionally with relatively pointed tip ( Fig. 2D View FIGURE 2 ), rostrum at an acute angle to colony surface, mandible hinged on a pair of short triangular condyles delimiting an approximately semicircular proximal area with an immersed calcified shelf (ML 97±12, 77–117, 20; MW 98±6, 87–110, 20), interior mandible with a large central oval lucida ( Fig. 2C View FIGURE 2 ); gymnocyst usually reduced to cystid corners, occasionally better developed all around.
Ancestrula not preserved.
Remarks. There has been considerable confusion as to the identity of this taxon ever since it was introduced by Gautier as Hincksina flustroides f. crassispinata Calvet (MS) in 1962. Initially, Calvet (1902, p. 31, pl. 1, fig. 3) recorded Membranipora flustroides from the Cette region (= Sète, Gulf of Lion, southeastern France), growing on basal thalli of a brown alga of the Cystoseira s.l. group. The figure is detailed enough to distinguish this species from H. flustroides . The ooecia are produced from the distal avicularium (not from the distal autozooid as in H. flustroides ), the proximal ooecial margin lacks the prominent peak, the avicularia and mandibles are distinctly more elongated, and the flattened spines were not recorded to be bifid. Though not figured by Calvet, optical examination of the types furthermore corroborated the presence of an extensive uncalcified region at the spine bases as later depicted by Prenant & Bobin (1966, fig. 62 II.–IV.).
The collection from Sète is now housed at the MNHN where two original specimens were examined optically: MNHN-IB-2008-6104 (a small colony on Cystoseira in ethanol, the label noting “ Membranipora flustroides var. crassispinata , n° 52, Cette”, Brescou Island, Cap d’Agde), and MNHN-IB-2008-8224 (one colony on Cystoseira in ethanol, the label is marked by a red dot and noting “ Membranipora flustroides var. crassispinata Calvet. Station Zoologique de Cette. Corr. Gautier Juin 1960. Hincksina flustroides var. crassispinata ”, Brescou Island, Cap d’Agde). The specimens were studied using a stereomicroscope at the MNHN because preparation for SEM-imaging may be difficult or impossible without damaging the colonies.
Judging by the labels of the two specimens, and according to the remarks in Gautier (1962, p. 51), Calvet himself must have changed his mind at some point after his 1902 publication, and regarded the Sète specimens he recorded as Membranipora flustroides as distinct from the British species. He then re-labelled the specimens to account for the new variety, without, however, publishing this new taxon before he died in 1930.
When revising Calvet’s collection at the Laboratoire de Biologie Marine de Sète in 1960, Y.V. Gautier came across these specimens, which then carried the name of the new variety intended to be given by Calvet. Gautier regarded these specimens to be identical with his own material from the Ligurian coast (N Italy) and the Siculo- Tunisian region, and presented the new form as Hincksina flustroides f. crassispinata, crediting Calvet with the authorship ( Gautier 1962, p. 50). The figure Gautier added to the description (1962, fig. 8), however, was not of Calvet’s but of one of his own specimens from Italy, which has thin cylindrical spines without any apparent organic joints. This is inconsistent with the name crassispinata (from the Latin crassus = thick), with the original drawing by Calvet (1902, pl. 1, fig. 3), and indeed with the morphology of the two Calvet specimens from Sète that do have flattened and distinctly jointed spines. Neither of the two colonies Gautier reported, which grow on the inner side of bivalve shells, contain zooids with flattened spines.
It is impossible to retrace why Gautier thought that his specimens were identical with Calvet’s given the difference in spine morphology. Although spine shape may indeed vary from cylindrical to flattened within the same colony in Hincksina species (see below), Gautier (1962) did not mention this variability to occur in a single colony, while he did state that the morphotype with thin spines (i.e. the new form) is found more often in the Mediterranean Sea than the typical flustroides - type with flattened spines (i.e. Calvet’s material). Thus, he did not realise that the specimens originally recorded as Membranipora flustroides by Calvet (1902) and the specimens he found labelled as Membranipora flustroides var. crassispinata were identical. This is demonstrated by the fact that Gautier regarded Calvet’s (1902) original record of the specimens on Cystoseira as synonymous with the specimens he considered to belong to Hincksina flustroides in the same work on the previous page ( Gautier 1962, p. 49).
Moreover, the measurements Gautier (1962) provided of H. flustroides f. crassispinata do not match with the dimensions of the zooids in his fig. 8. Whereas he reported zooid lengths between 0.65–0.75 mm, the depicted zooids are merely 0.47 mm long on average. Both the scale bar and the drawing he provided are therefore correct as they exactly match the measurements taken on Gautier’s specimen during the present study, while the measurements given by him are clearly erroneous.
Accordingly, Gautier introduced Hincksina flustroides f. crassispinata based on his own specimens and concept, not on Calvet’s, which would make Gautier the author of the form. In the literature, Calvet was occasionally given as author of the form (e.g. De Blauwe 2009), and it is presently regarded as a subspecies of H. flustroides (see Bock & Gordon 2021). The discussion on authorship is vain, however, owing to the fact that Gautier’s form has to be regarded as infrasubspecific because, to our knowledge, it has not been adopted as a valid name of a species or subspecies before 1985. According to Article 45.6.4.1 of the ICZN Code ( ICZN 1999), the name crassispinata is therefore unavailable, and a new species, Hincksina synchysia n. sp., is here introduced for Gautier’s material from Punta Carega on the Italian Ligurian coast.
Hincksina synchysia n. sp. is clearly distinct from Hincks’ H. flustroides in that it has distinctly more elongated avicularia with rounded triangular rostra and mandibles, and the spines were not observed to be bifid in the examined material. Moreover, in Hincksina synchysia n. sp. the ooecium is produced by both distal autozooids and avicularia. In all other previously established species the ooecium is formed either by the distal autozooid ( H. flustroides ) or by the distal avicularium (in the remaining species described herein). The Mediterranean populations figured by Harmelin (1988, pl. 1, fig, 4, but not fig. 5, which is H. calpensis ), Zabala & Maluquer (1988, text-fig. 86, pl. 3, fig. A), and Chimenz Gusso et al. (2014, fig. 40a, b), are here regarded to also belong to H. synchysia n. sp.
Other Mediterranean ( Hayward 1974, NHMUK 2009.10.12.4) and eastern Atlantic records (Madeira: Norman 1909, NHMUK 1911.10.1.346; Canary Islands: Arístegui Ruiz 1984, p. 56, fig. 11) are similar to H. synchysia n. sp. in spine and avicularium morphology but have to be left in open nomenclature as more material needs to be analysed morphologically and genetically before a decision can be made. While Hincksina sp. from southern Portugal, as figured by Souto et al. (2014, fig. 3E, F), differs from H. synchysia n. sp. in a few aspects (e.g. the common presence of kenozooids replacing avicularia; O. Reverter-Gil, pers. comm. 2021), the authors showed that both cylindrical and flattened spines can occur in zooids of the same colony that grow on different sides of a rock. It may thus be possible that Gautier’s and Calvet’s morphotypes are actually synonymous. For instance, the thicker spines in the specimens from Sète could represent an ecophenotype of H. synchysia n. sp. that, when growing on soft erect algae or on the exposed side of a rock, produces thicker spines in order to protect the frontal membrane from touching of the algal blades or from stronger currents with bedload, respectively. In contrast, the colonies on the underside of rocks or on the concave inner sides of bivalve shells, as in Gautier’s material, may be better protected in their microenvironment and thus produce thin cylindrical spines.
On the other hand, bryozoan species growing on soft algal blades or seagrass leaves are often particularly adapted to life on a flexible substratum and specifically different from congeners encrusting stable substrata. The extensive and distinct cuticular area at the base of the jointed spines in Calvet’s colonies that grow on Cystoseira is very likely one such adaptation, preventing breakage of the protective spines. In contrast, the spines in Hincksina synchysia n. sp. are apparently unjointed as the dark bands that indicate organic tissue in SEM images could not be observed in the examined specimens. Considering these differences, we regard the two morphotypes as distinct species for now.
The study of additional material from different regions, environments and microhabitats, assisted by genetic analyses, is necessary in order to assess the morphological variability and ecological preferences of H. synchysia n. sp., and to reveal its relationships with other Mediterranean and NE Atlantic Hincksina populations.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Flustrina |
|
SuperFamily |
Flustridae |
|
Family |
|
|
Genus |
Hincksina synchysia
| Berning, Björn, Spencer Jones, Mary E. & Vieira, Leandro M. 2021 |
Hincksina flustroides
| Gautier, Y. V. 1962: 50 |