
Hincksina alice (Jullien in Jullien & Calvet, 1903 )
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5081.3.2 |
|
publication LSID |
lsid:zoobank.org:pub:489D7694-F51B-4582-BE02-9C9EBD1F150A |
|
DOI |
https://doi.org/10.5281/zenodo.5776697 |
|
persistent identifier |
https://treatment.plazi.org/id/AE591B12-FF9F-0D58-FF0F-FB87B5A5D1E2 |
|
treatment provided by |
Plazi |
|
scientific name |
Hincksina alice (Jullien in Jullien & Calvet, 1903 ) |
| status |
|
Hincksina alice (Jullien in Jullien & Calvet, 1903) View in CoL n. comb.
( Fig. 5 View FIGURE 5 )
Membraniporella alice Jullien in Jullien & Calvet, 1903: 37, pl. 5, fig. 1; Calvet in Jullien & Calvet 1903: 124; Calvet 1907: 397.
part Membranipora albida View in CoL (?) Hincks: Busk 1884: 63, pl. 15, fig. 4.
part Membranipora lineata (Linné) : Waters 1898: 678, pl. 47, fig. 11; not pl. 48, fig. 9.
Material examined. Lectotype (here designated): MOM INV-22545, Hirondelle, 14/8/1888, Stn 226, 38°31.32’N 28°34.52’W, Calvet coll., Faial-Pico Channel (Azores), 130 m, colony fragment on bivalve shell, mounted on slide. Paralectotypes (all with the same locality data as lectotype): MOM INV-22461, colony on bioclast with ancestrula and early astogenetic stages, on slide; MOM INV-22520, one colony on bioclast, in vial; MOM INV-22545, one colony on bioclast, on slide; MOM INV-22590, one colony on bivalve together with Puellina sp. , Micropora sp. and Buffonellaria sp. , in ethanol; MOM INV-22622, one large colony on bivalve shell, in glass vial; MOM INV- 22623-22648, each with one colony fragment on slide. Non-type material examined using SEM: MNHN-IB-2008- 1996, Talisman , 13/8/1883, Stn 125, 38°29.1’N 28°37.3’W, Faial-Pico Channel (Azores), 80–115 m, five small colonies on bivalve shell fragments, ocurring together with Figularia sp. and Micropora sp. , Calvet coll., http:// coldb.mnhn.fr/catalognumber/mnhn/ib/2008-1996; MNHN-IB-2008-4009 (same locality data as MNHN-IB-2008- 1996), two colonies on bivalve shell fragments, Calvet coll., http://coldb.mnhn.fr/catalognumber/mnhn/ib/2008- 4009; NHMUK 1887.12.9.308, Challenger, 2/7/1873, Stn 75, 38°38’N 28°28’W, N of Pico Island (Azores), 820 m, volcanic mud and sand, two colonies on fragments of an erect bryozoan [formerly one large fragment figured by Busk (1884) on pl. 15, fig. 4b]; OLL 2021/3, Biaçores, 17/10/1971, Stn 88, 39°02.5’N 28°06.5’W, off Graciosa (Azores), 400–450 m, two colonies on a worn fragment of the erect Buskea ovalis ( Busk, 1881) , occurring together with Hemicyclopora canalifera ( Busk, 1884) , Hippothoa sp. , Puellina sp. and? Pyripora sp.
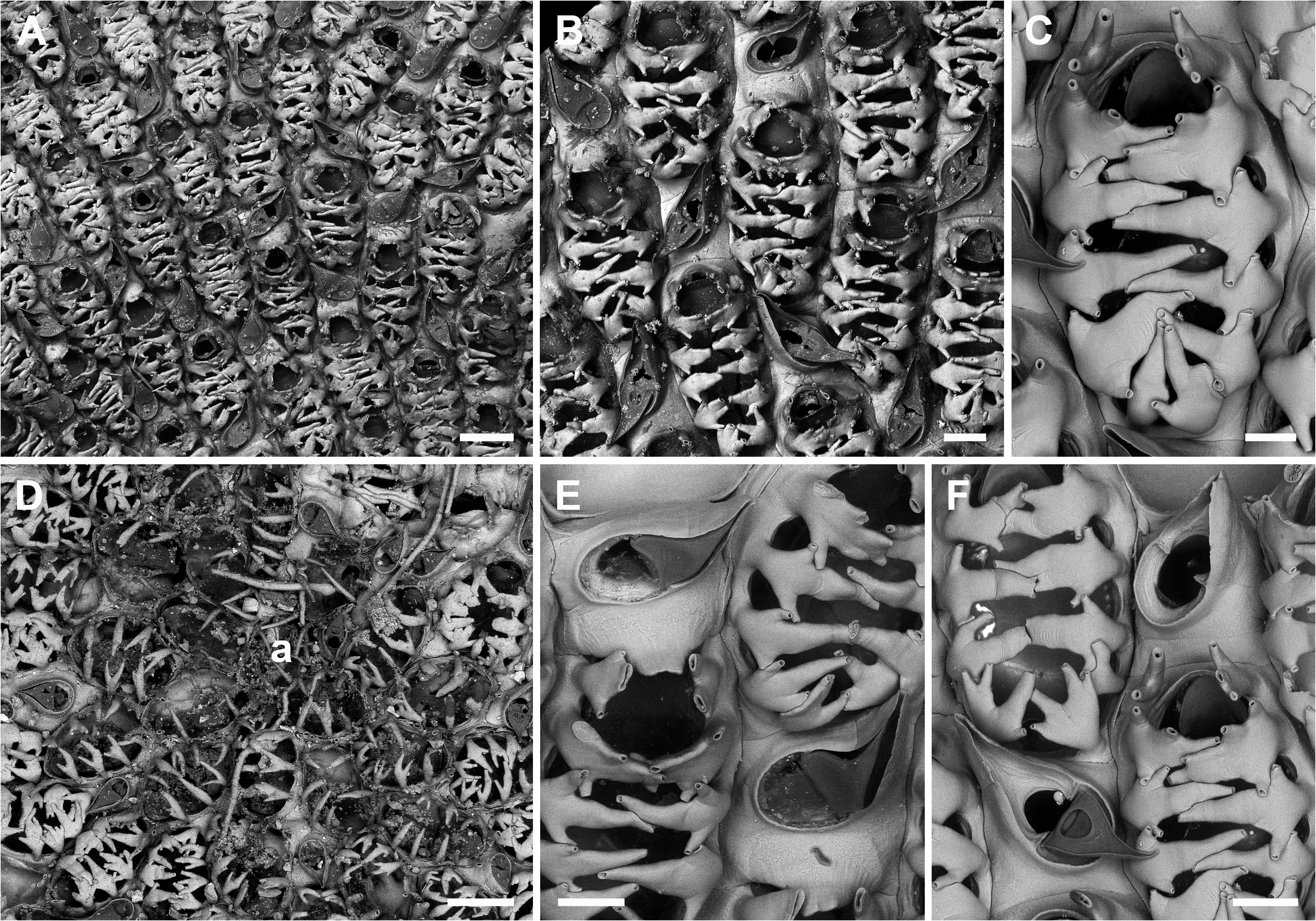
Description. Colony encrusting, unilaminar, multiserial. Autozooids oval (ZL 431±33, 364–492, 20; ZW 267±27, 234–324, 20), arranged in irregular series ( Fig. 5A View FIGURE 5 ), dried skeleton translucent, zooids separated by shallow grooves. Vertical walls with two or more uniporous pore plates per neighbouring zooid. Development of circum-zooecial gymnocyst extremely reduced or absent and often restricted to zooecial corners yet mural rim thickly calcified around spine bases ( Fig. 5B, C, E, F View FIGURE 5 ), cryptocyst practically absent, opesia therefore almost as long as zooecium (OpL 370±31, 310–433, 20; OpW 199±13, 170–222, 20). Opercular region laterally framed by a pair of flattened, fan-shaped or vaguely palmicorn oral spines, aligned oblique to zooidal midline, ramifying into 2–4 short branches, terminations straight and uncalcified ( Fig. 5B, C, E, F View FIGURE 5 ); frontal membrane in mature zooids overarched by 8–12 (most often 9–10) stout horizontally compressed mural spines that are very broad but vary in width between zooids and are of trident or vaguely palmicorn shape, growing vertically from their base and then bending inwards at a 90° angle over the frontal membrane while broadening and producing a pair of short and narrow lateral branches pointing disto- and proximomedially (with respect to zooid alignment) with the main median spine steadily narrowing towards its tip which extends beyond the zooidal mid line; all spine ends interdigitating, superimposing or occasionally colliding with the neighbouring/opposite spines; proximal mural spines shorter, distalmost spine pair broadest with their distal part upturned to provide space for tentacle extrusion; all spines in late astogenetic zooids unjointed ( Fig. 5B, C, E, F View FIGURE 5 ).
Ovicell subimmersed in distal avicularium, ooecium continuous with the gymnocyst of the avicularian cystid forming a short but broad hemispherical cap (OvL 93±15, 69–124, 20; OvW 172±21, 135–212, 20), proximal margin raised to produce a central peak ( Fig. 5E View FIGURE 5 ).
Avicularia interzooidal, distal to almost every autozooid ( Fig. 5A, B View FIGURE 5 ), pointing distolaterally, over twice as long as wide (AL 215±23, 186–252, 20; AW 97±10, 76–117, 20), positioned on a square cystid, smooth gymnocyst occasionally relatively extensive proximally, confined to the corner opposite of the rostrum distally ( Fig. 5B, F View FIGURE 5 ); rostrum elongate triangular and often gently curved, at an acute angle to colony surface with the tip slightly downcurved, mandible hinged on a pair of short triangular condyles delimiting an approximately semicircular proximal area (ML 134±14, 113–157, 20; MW 79±6, 68–92, 20), entire opesia oval, framed by an immersed calcified shelf slightly thinning laterally and increasing in width distally, avicularium widest proximal to condyles ( Fig. 5B, E, F View FIGURE 5 ), interior mandible marked by a large subcircular lucida ( Fig. 5F View FIGURE 5 ).
Ancestrula tatiform, periancestrular zooids with slender and cylindrical jointed spines arching over the frontal membrane, with slightly flattened and bifurcating spines in second- or third-generation zooids, occurrence of fully developed spines in sixth-generation zooids, avicularia distinctly shorter and not curved ( Fig. 5D View FIGURE 5 ).
Remarks. For the same reasons as given above for Hincksina neptuni , this species has to be transferred from the Cribrilinidae to the Flustridae . Hincksina alice is closely related with H. neptuni owing to the pointed avicularia. The characteristic mural spines of H. alice are also similar to those of H. sceletos and H. neptuni as regards their varied shape, but instead of being vertically flattened as in these species they are compressed horizontally in H. alice . Likewise, the flattened, bifurcating oral spines are shared with H. neptuni , H. sceletos as well as with H. calpensis .
Jullien & Calvet (1903, p. 37) did not indicate type material for the species while stating that several specimens were found at Hirondelle Stn 226; none of the material is present at the MNHN where Jullien’s collection is kept ( Tricart & d’Hondt 2009; B. Berning, pers. observ.). A large number of specimens from the same station exist, however, in Calvet’s collection at the MOM. As it is impossible to judge from the original drawing if (and precisely which) one of these specimens was figured, one of the best-preserved colonies is here designated as lectotype of H. alice (MOM INV-22462).
Part of the material (i.e. the specimens from the Azores) that was recorded by Busk (1884, p. 63) as Membranipora albida (?) Hincks, 1880c (a species originally described from Singapore) belongs to H. alice . Slightly more problematic to interpret is Waters’ (1898) account on what is supposed to be the same material [identified as Membranipora lineata ( Linné, 1758) , which is now placed in the genus Callopora Gray, 1848 ]. Whereas his drawing of one specimen (pl. 47, fig. 11) presumably depicts H. alice , his second drawing of pointed teeth within basal pore chambers is most likely based on a different taxon as Hincksina does not produce basal pore chambers. Both authors were presumably misled in their systematic interpretation by the fact that in all but one zooid the spines are abraded in the colonies from Challenger Stn 75.
As H. neptuni , H. alice is also known only from the Azorean islands of Faial, Pico and Graciosa (central group), and has been collected from depths between 80 and 820 m where it encrusts rocks, shells and other biogenic substrata. The colonies from the deep Biaçores Stn 88 (400–450 m) and Challenger Stn 75 (820 m) are, however, dead and abraded, possibly indicating downslope transport. Living colonies have only been recovered from 80 to 130 m depth. When introduced, H. neptuni and H. alice were found to occur sympatrically in the Faial-Pico Channel ( Jullien & Calvet 1903). Surprisingly, however, neither of the species were recorded from settlement panels deployed in this region at depths between 0 and 500 m for up to two years in 2006–2008 ( Wisshak et al. 2015).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
SubOrder |
Flustrina |
|
SuperFamily |
Flustridae |
|
Family |
|
|
Genus |
Hincksina alice (Jullien in Jullien & Calvet, 1903 )
| Berning, Björn, Spencer Jones, Mary E. & Vieira, Leandro M. 2021 |
Membraniporella alice
| Calvet, L. 1907: 397 |
| Jullien, J. & Calvet, L. 1903: 37 |
| Jullien, J. & Calvet, L. 1903: 124 |
Membranipora lineata (Linné)
| Waters, A. W. 1898: 678 |
Membranipora albida
| Busk, G. 1884: 63 |