
Steinernema apuliae, Triggiani, Oreste, Ek, Zdenek Mrá Č & Reid, Alex, 2004
|
publication ID |
https://doi.org/ 10.5281/zenodo.157848 |
|
publication LSID |
lsid:zoobank.org:pub:25F1DC96-D206-4658-8F6F-9242EC771471 |
|
DOI |
https://doi.org/10.5281/zenodo.5619141 |
|
persistent identifier |
https://treatment.plazi.org/id/AE6F4B07-C27B-FFBE-FEAD-055D945456B0 |
|
treatment provided by |
Plazi |
|
scientific name |
Steinernema apuliae |
| status |
sp. nov. |
Steinernema apuliae sp. n. ( Figures 1–3)
Measurements: Morphometrics of first and second generation paratype males of S. apuliae sp. n. are given in Table 1, morphometrics of first and second generation paratype females in Table 2 View TABLE 2 and morphometrics of the holotype (thirdstage infective juvenile) and of thirdstage paratype juveniles in Table 3 View TABLE 3 .
TABLE 1: Morphometrics of first and secondgeneration paratype males of Steinernema apuliae sp. n. (mean, standard deviation of the mean, range; all measurements in µm).
* spicule length divided by anal body width, ** gubernaculum length divided by spicule length.
* Hyaline tail length in % of total tail length.**Anterior end to excretory pore in % of pharynx length.
Morphology
First generation males: body curved ventrally posteriorly, tapering toward lip region, Jshaped in heatrelaxed specimens. Cuticle with very faint transverse striae, lateral fields absent. Head rounded, almost continuous with body. Six lips fused at base, each lip bearing a labial papilla. Outer circle of four cephalic papillae present. Amphids distinct, located laterally between labial and cephalic circle of papillae ( Fig. 1 B). Excretory pore ( Fig. 1 C) variable in position, rarely slightly anterior, mostly at, and sometimes posterior to nerve ring (ranging from 75% to 92% of the length from anterior body end to base of pharynx). Stoma short, shallow, cheilorhabdions distinct, sclerotized, followed by the sclerotized smaller, sometimes indistinct prorhabdions. Pharynx muscular with procorpus extending into slightly swollen nonvalvate metacorpus, a narrower isthmus region surrounded by a nerve ring, and a rounded basal bulb. Cardia rather long, intestine generally with a wide lumen. Distinct, papilliform deirids situated at or slightly behind pharyngeal bulb. Well developed and reflexed testis. Paired symmetrical, slightly curved spicules and light brown in colour, distally bluntly pointed ( Fig. 1 G). Spicule manubrium distinctly elongated, with length: width ratio of 1.2–1.7:1; manubrium less then one third of spicule length (about 29%); calomus distinct, lamina slightly arched, ventral rostrum absent; velum narrow, indistinct, parallelly arched with lamina, extending from calomus to distal part of lamina. Gubernaculum boatshaped in lateral view, distally much attenuated, proximal part varying from straight and bluntly pointed to slightly bentlike ( Fig. 1 H). Tail bluntly conoid, usually concave at ventral side, mucron absent. There are 23 genital papillae comprising 11 pairs and a single ventral papilla located just in front of the cloacal opening. Of the paired papillae, 6 pairs are located precloacal. Of these 5 pairs situated subventrally and 1 pair laterally. Five pairs postcloacal. Of these 2 pairs located adcloacal and subventral, very close to each other. The last 3 pairs are consistently located near the tail tip ( Fig. 1 D).
Second generation males: similar to the firstgeneration males, but slightly smaller in the body length and other measurements (Table 1; Fig. 2A View FIGURE 2. A ). Greatest body width much smaller. Excretory pore located more anterior than in the first generation (Table 1; Fig. 2A View FIGURE 2. A ). Tail mucron absent ( Fig. 2 View FIGURE 2. A C).
First generation females: body long, usually Cshaped on heat relaxation ( Fig. 1A). Cuticle only faintly striated. Head bluntly rounded, slightly tapering anteriorly. Morphology of head region and pharynx similar to the males ( Fig. 3A View FIGURE 3. A ). Excretory pore located rarely anterior, mostly at, and sometimes posterior to the nerve ring. Genital tract amphidelphic with reflexed ovaries. Vulva asymmetrical and oblique slit to symmetrical and transverse slit, slightly (3 µm) to distinctly (35 µm) protruding from the body contour (depending on the age). Slant and short vagina, leading into paired uteri. Narrow rectum, distinct anal opening. Tail of mature females short and obtuse, with small postanal lobe, almost long as is wide; in young females a longer tail with conicallike tip, which is more prominent in older females with the obtuse tail. The tip bears one large and two or three minute protuberances or papillae protruding from the body contour ( Fig. 3 View FIGURE 3. A B, 3C and 3D). Pigmy forms rarely observed.
Second generation females: similar to first generation females in general morphology, but smaller in dimension ( Table 2 View TABLE 2 ). Excretory pore located more anterior than in the first generation females. Vulva distinctly asymmetrical and oblique slit in all observed specimens ( Fig. 3 View FIGURE 3. A E and 3F). Conical, pointed tail, with a short protuberance or papillalike structures on its tip, but not mucronated; tail longer than body width at anus ( Fig. 3 View FIGURE 3. A G). Pygmy forms not observed.
Infective thirdstage juveniles: body of heatrelaxed specimens slightly Cshaped to almost straight, slender, slightly tapering towards anterior and posterior ends. Morphometrics of infectivestage juveniles are reported on Table 3 View TABLE 3 . Specimens from Galleria cultures often enclosed in secondstage cuticle that is indistinctly annulated and 1.5 to 2 µm thick in midbody region. Lateral fields at midbody region with eight equally distributed ridges (i.e. nine lines); all ridges and lines distinct and at approximately even distances ( Fig. 3 View FIGURE 3. A H). Phasmids weekly developed, posterior to the anus, with their opening approximately 40% of tail length located at ventral edge of the lateral fields. Head portion finely rounded, mostly slightly offset from body contour by a depression of 11–13 µm wide ( Fig. 1 E). Four cephalic papillae and amphids indistinct. Papilliform deirids situated slightly behind pharyngeal bulb. Closed stoma. Pharynx with slightly expanded procorpus, narrower isthmus and subpyriform basal bulb with weakly developed valve apparatus. Basal bulb usually slightly displaced towards dorsal side by the excretory gland. Lentillike hemizonid, about 5 µm long, mostly varying in position from beginning of bulb to its midsection.
Excretory pore posterior to midpharynx region (on average about 66% of the length from the anterior end to the base of the pharynx), cuticularized, about 1 µm wide, excretory duct extending posteriorly to the cardia region ( Fig. 3 View FIGURE 3. A I). Intestine separated from pharynx by a long cardia, which is followed by a slender bacterial vesicle. Gut filled with numerous fat globules, lumen not visible. Straight rectum, almost as long as anal body diameter. Anus 4 5 µm wide, sickleshaped. Tail relatively wide (low ratio C'), conoid, straight, with terminus finely pointed, tip devoid of refractive inclusion ( Fig. 1 F). Hyaline portion occupying on average 41–42% of tail length ( Fig. 3 View FIGURE 3. A L).
DNA analysis
The RFLP (restriction fragment length polymorphism) profiles yielded by digestion of the ITS region with 17 restriction enzymes for Steinernema apuliae sp. n. ( Fig. 4 View FIGURE 4 A) were compared to a database containing over 50 different Steinernema isolates (including 22 described species). Examination of the RFLP profiles showed S. apuliae sp. n. to be unique. S. apuliae sp. n. can be differentiated from S. arenarium ( Fig. 4 View FIGURE 4 B) by 9/17 restriction enzymes (Alu I, Dde I, Hae III, Hha I, Hind III, Hinf I, Pvu II, Rsa I and Sal I) and from S. glaseri ( Fig. 4 View FIGURE 4 C) by 11/17 enzymes (Alu I, Dde I, Hae III, Hha I, Hind III, Hinf I, Hpa II, Pvu II, Rsa I, Sal I and Sau3A I).
Type of locality and host
The type population was collected by using Galleria mellonella (L.) larvae as bait from a soil sample picked up from a saltpan near Margherita di Savoia, in the district of Bari at an altitude of 30 m above sea level, in October 1996. The natural host is unknown.
Type of material
Holotype infective juvenile, paratype thirdstage juveniles, males and females of first and second generations are deposited in the Laboratory of Insect Pathology, Institute of Entomology, Czech Academy of Sciences, Č eské Budĕjovice. Additional paratypes (thirdstage juveniles, males and females) are at the Department of Biology and Chemistry, Section of Entomology and Zoology, University of Bari; paratype thirdstage juveniles are in the Nematode Collection of CABI Bioscience, Egham, U.K.
Differential diagnosis
Steinernema apuliae sp. n. is mainly characterized by the following morphological characteristics: males with light brown, slightly curved spicules with longitudinally shaped manubrium and absence of a tail mucron in both generations. Females possess a characteristically asymmetrical oblique slit vulva distinctly behind the midbody. Position of excretory pore is mostly posterior in both sexes. One distinct and two or three minute protuberances or papillae are located on the first female tail tip. Thirdstage juveniles have long body, on average over 1000 µm. Hyaline tail portion is medium long, slightly over 40% of the tail length. Excretory pore is near the posteriorpharynx region (5877%). Lip region broad, continuous, and rounded. Lateral fields with equally developed eight ridges (equivalent to nine lines) at even distances.
Steinernema glaseri and S. arenarium are the only described Steinernema species found in Europe closely resembling S. apuliae sp. n. in morphometrics and other morphological characters ( Poinar, 1978; de Doucet & Gabarra, 1994; Mráček et al., 1999). However, females of S. apuliae sp. n. have an asymmetrical, oblique slit vulva, whereas S. glaseri and S. arenarium females have a more symmetrical and transverse slit vulva. Vulval opening is slightly posterior in S. apuliae females. Males of S. glaseri possess a hooked spicule tip that was never observed in S. apuliae and S. arenarium . Hyaline tail portion of IJs of S. apuliae seems to be slightly shorter. Pair of marginal ridges is slightly annulated in S. glaseri and smooth in S. arenarium and S. apuliae . Finally, PCRRFLP analysis can be used to clearly differentiate these species.
More species, such as S. cubanum Mráček, Hernández & Boemare 1994 , S. puertoricense Roman & Figueroa 1994 , S. longicaudum Shen & Wang 1992 , S. karii Waturu, Hunt & Reid 1997 , S. loci Phan, Nguyen & Moens, 2001 and S. diaprepesi Nguyen & Duncan, 2002 , belong to the “glaserigroup”. There is an important zoogeographical barrier for torrid species of S. cubanum , S. puertoricense , S. karii , and S. loci . The infective juvenile ratio (D%) is very low for S. karii (56.6%) and S. loci (57%), whereas 66% for S. apuliae .
The ratio c´for S. longicaudum IJs is very high (4.22) due to the tail length over 90 µm tail length, as it is for S. puertoricense . S. diaprepesi has shorter IJs, about 1000 µm, and a ratio (D%) close to 50%.
The morphology of males is similar among these species; however, S. apuliae possesses a strongly asymmetrical vulva that differs from that with a double flapped epiptygma of S. puertoricense . S. cubanum is morphologically and morphometrically very similar to S. apuliae ; however, its vulva position is about 50% that of the body length whereas in S. apuliae it is significantly, behind the midbody, varying mostly between 55% to 57% of the body length.
Life cycle and crossbreeding
Steinernema apuliae sp. n. is easily reared on G. mellonella larvae in the laboratory at 20 to 22°C. The Galleria larvae were killed after two days of exposition to IJs; first generation of adults appeared three days after host death occurred and new IJs started to emerge after one week. There were no positive interbreedings of S. apuliae with S. glaseri and S. arenarium .
Distribution and ecology
Steinernema apuliae sp. n. has been found exclusively in southern Italy in the Apulia and Basilicata regions ( Fig. 5 View FIGURE 5 ) ( Triggiani & Tarasco, 2002). Isolates were only found in sandy soil with one exception in silty soil (Table 4). They prevailed on seacoast with scant vegetation and a pinewood forests where Thaumetopoea pityocampa species ( Lepidoptera , Thaumetopoeidae ) is common.
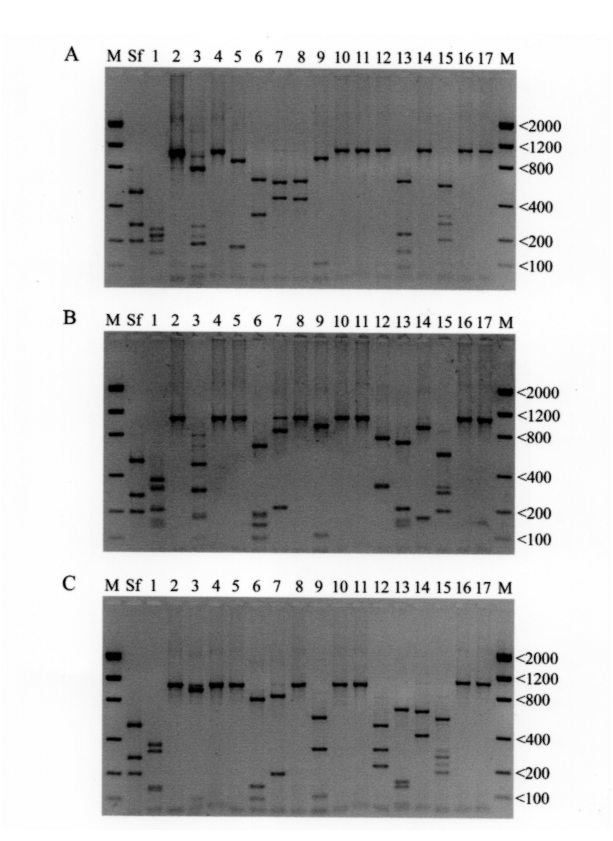
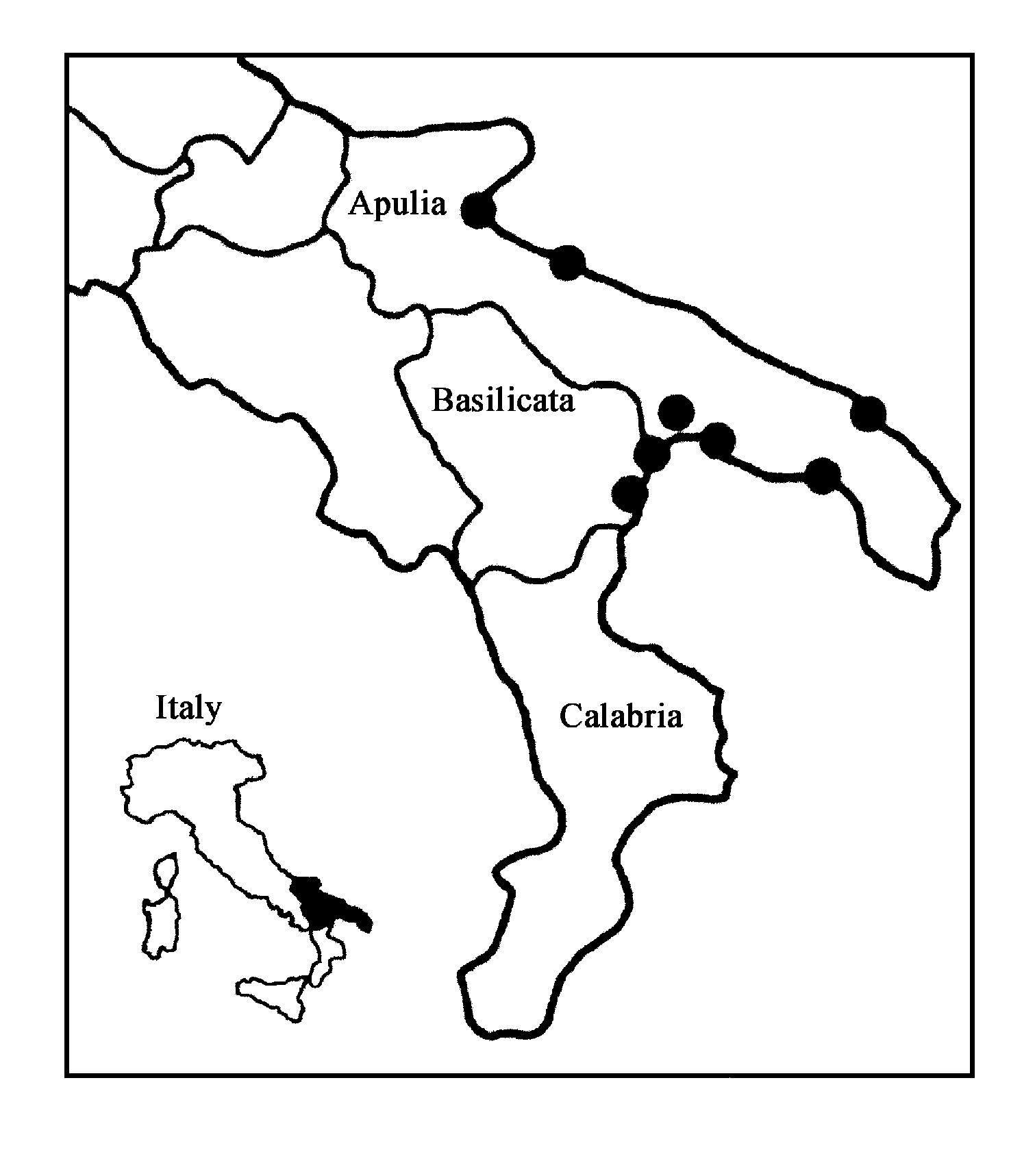

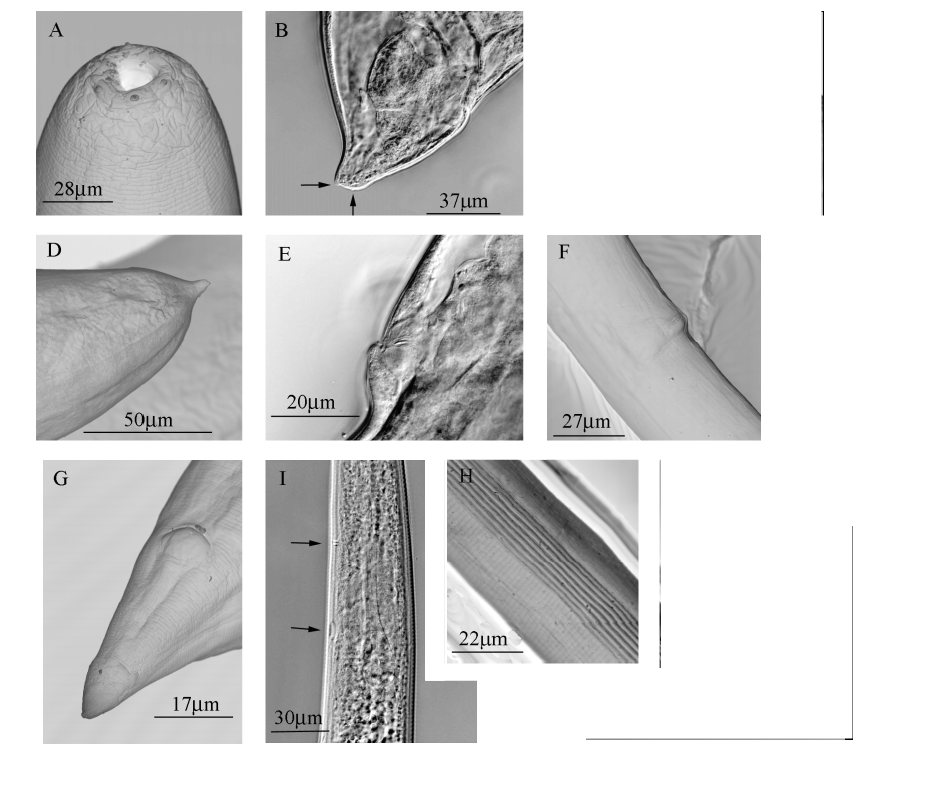
TABLE 2. Morphometrics of first and secondgeneration paratype females of Steinernema apuliae sp. n. (mean, standard deviation of the mean, range; all measurements in µm).
| Character | First generation (n = 15) | Second generation (n = 20) |
|---|---|---|
| Body length | 5086 ± 1700 (2828 – 9474) | 2362 ± 433 (1818 – 3192) |
| Greatest diameter | 271 ± 40 (227 – 383) | 144 ± 25 (105 – 196) |
| Stoma length | 8.1 ± 1.1 (6.4 – 11.5) | 5.5 ± 0.6 (4.7 – 6.2) |
| Stoma width | 14 ± 1.2 (11 – 16) | 9.3 ± 0.8 (7.9 – 10.6) |
| Anterior end to Excretory pore | 184 ± 15 (141 – 199) | 158 ± 18 (129 – 187) |
| Pharynx length | 229 ± 11 (207 – 246) | 225 ± 24 (196 – 266) |
| Tail length | 62 ± 8.2 (47 – 74) | 61 ± 4.4 (51 – 67) |
| Anal body width | 69 ± 6.2 (54 – 78) | 46 ± 9.7 (35 – 70) |
| A | 19 ± 5.7 (11 – 33) | 16 ± 1.6 (12 – 19) |
| B | 22 ± 6.8 (13 – 39) | 14 ± 2.1 (12 – 21) |
| C | 82 ± 29 (52 – 173) | 46 ± 5.6 (40 – 64) |
| C' | 3 ± 0.3 (2.4 – 3.6) | 1.3 ± 0.1 (1 – 1.5) |
| V | 55 ± 1.8 (50 – 58) | 57 ± 1.9 (55 – 61) |
| D % | 80 ± 4.7 (68 – 87) | 70 ± 5.1 (60 – 78) |
TABLE 3: Morphometrics of infectivestage juveniles of Steinernema apuliae sp. n. (mean, standard deviation of the mean, range; all measurements in µm).
| Character | Holotype | Paratypes (n = 19) | Laboratory populations (n = 20) |
|---|---|---|---|
| Body length | 1192 | 1064 ± 66 (945 – 1212) | 1063 ± 81 (869 – 1192) |
| Greatest diameter | 38 | 37 ± 1.9 (34 – 41) | 36 ± 1.8 (33 – 40) |
| Anterior end to Excretory pore | 100 | 95 ± 4.2 (86 – 102) | 93 ± 21 (41 – 57) |
| Anterior end to Nerve ring | 119 | 111 ± 5.3 (103 – 104) | 109 ± 6.7 (100 – 122) |
| Pharynx length | 146 | 145 ± 6.4 (137 – 159) | 147 ± 8.5 (134 – 164) |
| Tail length | 73 | 71 ± 5.4 (63 – 80) | 74 ± 6.6 (64 – 83) |
| Hyaline tail length | 33 | 30 ± 2.7 (25 – 34) | 31 ± 2.6 (27 – 35) |
| Anal body width | 25 | 23 ± 1.4 (20 – 26) | 23 ± 1.2 (21 – 25) |
| A | 31 | 29 ± 1.4 (26 – 31) | 29 ± 2 (26 – 30) |
| B | 8.2 | 7.3 ± 0.3 (6.8 – 7.8) | 7.2 ± 0.6 (6.5 – 7.3) |
| C | 16.3 | 15 ± 0.7 (14 – 17) | 14 ± 0.7 (14 – 16) |
| C' | 2.9 | 3.0 ± 0.2 (2.7 – 3.3) | 3.3 ± 0.3 (2.8 – 3.8) |
| E | 1.4 | 1.3 ± 0.1 (1.2 – 1.5) | 1.3 ± 0.1 (1.1 – 1.5) |
| H % * | 46 | 42 ± 2.6 (38 – 47) | 41 ± 2.5 (35 – 45) |
| D %** | 68 | 66 ± 2.3 (63 – 70) | 66 ± 0.05 (58 – 77) |
| CABI |
CABI Genetic Resource Collection |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Steinernema apuliae
| Triggiani, Oreste, Ek, Zdenek Mrá Č & Reid, Alex 2004 |
S. diaprepesi
| Nguyen & Duncan 2002 |
S. loci
| Phan, Nguyen & Moens 2001 |
S. karii
| Waturu, Hunt & Reid 1997 |
S. cubanum Mráček, Hernández & Boemare 1994
| Mracek, Hernandez & Boemare 1994 |
S. puertoricense
| Roman & Figueroa 1994 |
S. longicaudum
| Shen & Wang 1992 |