
Girardia arenicola Hellmann & Leal-Zanchet
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4438.3.8 |
|
publication LSID |
lsid:zoobank.org:pub:6BB71AE3-6D5B-4C55-9BC0-8E853AC05928 |
|
DOI |
https://doi.org/10.5281/zenodo.5969693 |
|
persistent identifier |
https://treatment.plazi.org/id/B06687E3-D375-6C35-85B0-0FE6FB0CFABE |
|
treatment provided by |
Plazi |
|
scientific name |
Girardia arenicola Hellmann & Leal-Zanchet |
| status |
|
Girardia arenicola Hellmann & Leal-Zanchet ; sp. nov.
Etymology: The name is formed from the Latin arena (sand), referring to the name of the type-locality (Areias = arena), and the Latin verb cólere (to inhabit).
Material examined. Holotype: MZUSP PL.2137: coll. R.L. Ferreira, 19 Julho 2012, “ Areias de Cima cave”, “Areias system”, Iporanga, São Paulo, Brazil—sagittal sections on 41 slides.
Paratypes: collected by R.L. Ferreira on the same date and the same sampling site as the holotype. MZU PL. 00275: sagittal sections on 11 slides; MZU PL. 00276: sagittal sections on 15 slides; MZU PL. 00277: sagittal sections on 23 slides; MZU PL. 00278: sagittal sections on 6 slides.
Type-locality: “ Areias de Cima cave”, “Areias system”, Iporanga, São Paulo, Brazil.
Distribution: Known only from the type-locality.
Diagnosis: Troglobitic Girardia arenicola is characterized by dorsal testes, a large and branched bulbar cavity containing various diverticula, a bursal canal almost horizontally disposed, a short common ovovitelline duct and male and female atria being separated by a constriction.
Description. External features. Live and preserved specimens whitish, both dorsally and ventrally. Triangular head with large, pointed auricles in live specimens, and a pair of small eyes; posterior tip pointed ( Figs. 5–7 View FIGURES 5–7 ). Body up to 14 mm long and 2 mm wide ( Table 1). Mouth located in the median third of the body and gonopore in the posterior third of the body ( Table 1; Fig. 7 View FIGURES 5–7 ).
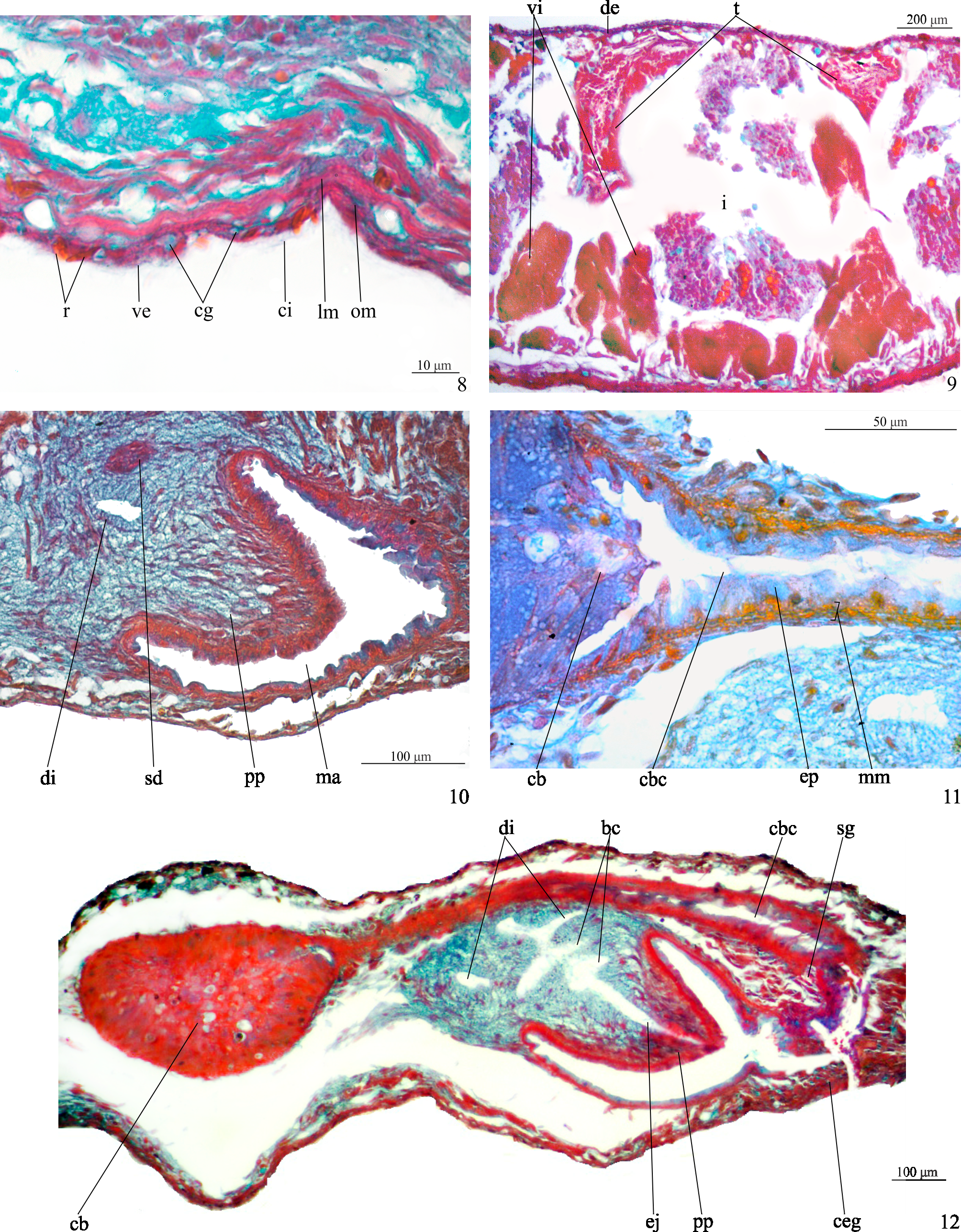
Epidermis, cutaneous musculature and sensory organs. The epidermis is penetrated by a xanthophil, rhabditogen secretion (rhammites), as well as by sparse secretions of three types of glands: (1) xanthophil, finely granular secretion; (2) erythrophil, finely granular secretion; (3) cyanophil amorphous secretion ( Fig. 8 View FIGURES 8–12 ). Cyanophil glands are more abundant in the anterior extremity of the body; openings from a fourth type of gland, producing a xanthophil coarsely granular secretion, concentrated at the body margins and medially at the anterior and posterior tips of the body.
Cutaneous musculature composed of three layers, viz. a thin subepithelial circular layer, followed by an oblique layer with decussate fibers and a thicker layer of longitudinal muscle. The cutaneous musculature is weakly developed; the ventral (10–12 µm thick), about twice as thick as the epidermis, is slightly thicker than the dorsal musculature (5–8 µm thick) in the pre-pharyngeal region.
Digestive system. Pharynx cylindrical, non-pigmented; about 1/3 to 1/4 of the body length. It is located in the median third of the body; mouth at the posterior end of the pharyngeal pouch, which shows a posterior diverticulum. No esophagus was observed. Anterior ramus of the intestine trunk dorsally to the brain.
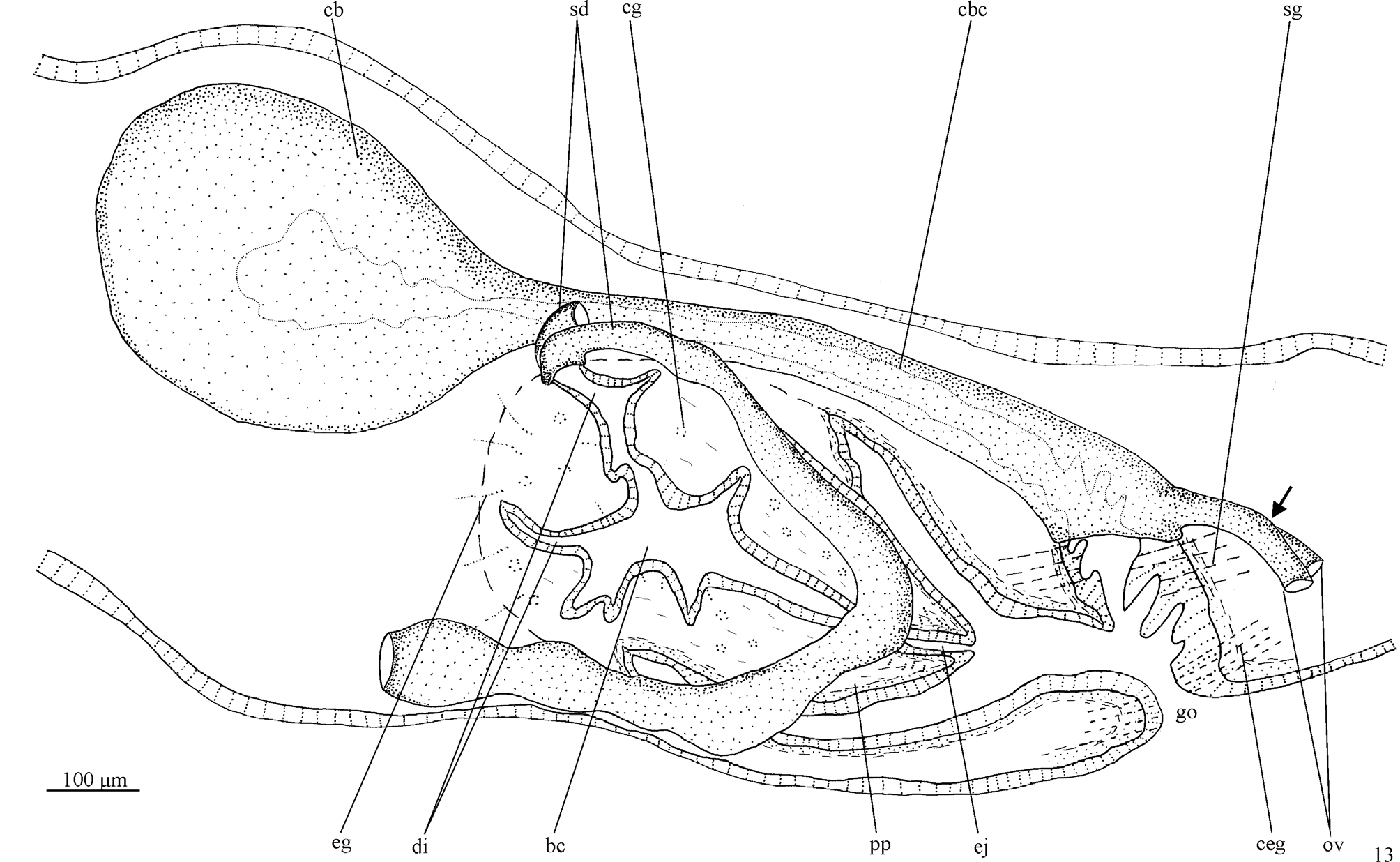
Male reproductive system ( Figs. 9–10, 12–13 View FIGURES 8–12 View FIGURE 13 ). Numerous testicular follicles, about 100–150 µm in diameter, arranged in two to three irregular lateral rows on either side of the body, being located next to the body margins, most of the follicles located in the dorsal body region ( Fig. 9 View FIGURES 8–12 ). Testes begin in the anterior 1/6th of the body, just behind the ovaries, and extend to the posterior end of the body. Sperm ducts dorsomedially to the nerve cords, expanding to form spermiducal vesicles posteriorly to the pharynx. The ducts ascend laterally to the penis, forming a loop and then recurve anteriad. While diminishing in diameter, they separately penetrate the penis bulb. The sperm ducts open symmetrically into the lateral diverticula of the bulbar cavity ( Fig.10 View FIGURES 8–12 ), which is large and branched, containing various diverticula ( Figs. 12–13 View FIGURES 8–12 View FIGURE 13 ). A long and narrow ejaculatory duct traverses the conical penis papilla. The latter is about 190 µm long and 150 µm wide at its basis, being obliquely orientated in the male atrium ( Figs. 12–13 View FIGURES 8–12 View FIGURE 13 ).
Bulbar cavity and ejaculatory duct receive the openings of abundant cyanophil penial glands, as well as scattered erythrophil glands, both glands producing a granular secretion. In addition, openings from glands producing a finely erythrophil secretion are more abundant at the distal, posterior portion of the ejaculatory duct ( Fig. 12 View FIGURES 8–12 ). The lining epithelium of the penis papilla and male atrium are penetrated by the sparse openings of two types of gland, one producing a amorphous, cyanophil secretion and the other a finely granular, erythrophil secretion ( Fig. 12 View FIGURES 8–12 ).
Female reproductive system ( Figs. 9, 11–13 View FIGURES 8–12 View FIGURE 13 ). Vitellaria well developed, located dorsally and between intestinal branches ( Fig. 9 View FIGURES 8–12 ). Ovaries ovoid, about 80–90 µm in its lateral axis in the holotype. The gonads are situated dorsally to the ventral nerve cords, in close proximity to the brain, in the anterior 1/10th of the body. Ovovitelline ducts arising from the lateral surface of the ovaries and running backwards dorsally to the ventral nerve cords and ventrally to the sperm ducts. Just behind the gonoduct, the ovovitelline ducts slightly turn dorsally and fuse to form a short common section, which opens into the most distal part of the bursal canal, about halfway the distance between the dorsal and ventral epidermis ( Fig. 13 View FIGURE 13 ). Copulatory bursa ovoid with a narrow lumen, located next to the pharyngeal pocket. Bursal canal long, almost horizontally disposed, slightly bending and becoming wider just before communicating with the gonoduct ( Figs. 12–13 View FIGURES 8–12 View FIGURE 13 ). Gonoduct almost straight ( Fig. 13 View FIGURE 13 ).
Ovovitelline ducts lined with ciliated, cuboidal epithelium with insunk nuclei, turning to a nucleated epithelium close to their joining to form the common ovovitelline duct. Bursal canal lined with a nucleated, columnar epithelium ( Fig. 11 View FIGURES 8–12 ), showing an irregular height close to the opening into the gonoduct. Copulatory bursa and bursal canal coated with a thin musculature (about 3–4 µm thick), which is composed of interwoven longitudinal and circular muscles. Shell glands, producing a finely granular, erythrophil secretion open into the bursal canal close to the opening of the common ovovitelline duct ( Figs. 12–13 View FIGURES 8–12 View FIGURE 13 ).. Numerous cement glands, producing a coarsely granular, xanthophil secretion, open into the gonoduct ( Figs. 12–13 View FIGURES 8–12 View FIGURE 13 ). In addition, the gonoduct receives the openings of other two types of scarcer glands, one producing an amorphous, cyanophil secretion and the other an erythrophil secretion.
Remarks. Specimens show fixation artifacts especially in the mesenchyme. Paratype MZU 0 0 278 shows a forked pharynx. Paratypes MZU PL.00275, MZU PL.00276 and MZU PL. 0 0 277 lost their pharynges during fixation.
Comparative discussion. With respect to its external features, Girardia arenicola differs from its congeners by having both a whitish body, as well as eyes. It resembles the hypogean Girardia mckenziei (Mitchell & Kawakatsu, 1973) , which has two minute eyes. However, G. mckenziei has a slightly pigmented body, whereas G. arenicola maintains a whitish colour after fixation, similarly to other troglobitic species ( Mitchell & Kawakatsu 1973a, b; Souza et al. 2015, 2016).
Regarding the reproductive system, G. arenicola shares some similarities with species that show dorsal or dorso-ventral testes, an almost horizontally disposed bursal canal and male and female atria separated by a constriction. These species are the following: G. anderlani ( Kawakatsu & Hauser, 1983) , Girardia arimana (Hyman, 1957) , G. chilla ( Marcus, 1954) , G. guatemalensis (Mitchell & Kawakatsu, 1973) , and G. paramensis (Fuhrmann, 1912) ( Mitchell & Kawakatsu 1973b; Kawakatsu & Mitchell 1981; Kawakatsu et al. 1983; Sluys 1992; Sluys et al. 2005). It mainly differs from these species by the shape of the bulbar cavity and/or the course of the spem ducts through the penis bulb. In Girardia arimana , the sperm ducts traverse the penis bulb and open into the ejaculatory duct and in G. guatemalensis , G. chilla and G. paramensis there is a tubular bulbar cavity ( Marcus 1954; Kawakatsu & Mitchell 1981; Kawakatsu & Froehlich 1992; Sluys 1992; Sluys et al. 2005), differing from the large and branched bulbar cavity with multiple diverticula of G. arenicola . In G. anderlani , the bulbar cavity is wide and with two chambers, i.e. an antero-ventral one, which receives both sperm ducts, and a blindly ending postero-dorsal chamber ( Kawakatsu et al. 1983). In contrast, G. arenicola shows a bulbar cavity with multiple diverticula, receiving the sperm duct openings on each side. In addition, it is noteworthy the occurrence of a common ovovitelline duct, similarly to G. arimana , among others. However, G. arenicola can be easily differentiated from G. arimana , since in G. arimana the common ovovitelline duct is relatively longer and the sperm ducts traverse the penis bulb, joining to open directly into the ejaculatory duct ( Sluys 1992).
The shape of the bulbar cavity of G. arenicola , as well as the openings of the sperm ducts in this cavity resemble the situation in the troglobitic G. multidiverticulata Souza et al., 2015 , which shows a large bulbar cavity, with multiple diverticula, receiving the opening of each sperm duct in one of its lateral diverticula ( Souza et al. 2015). However, G. multidiverticulata shows mainly ventral testes, contrasting to the mainly dorsal testes of G. arenicola . Other details of the copulatory apparatus distinguish G. arenicola from G. multidiverticulata , since G. arenicola has a smaller number of diverticula in the bulbar cavity and a large and conical penis papilla with a long and slender ejaculatory duct. In contrast, G. multidiverticulata shows a large number of diverticula in the bulbar cavity, a short and blunt penis papilla and a small and wide ejaculatory duct ( Souza et al. 2015).

| MZUSP |
Museu de Zoologia da Universidade de Sao Paulo |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.