
Simulium litobranchium Hamada, Pepinelli, Mattos-Gloria & Luz
|
publication ID |
https://doi.org/10.5281/zenodo.194650 |
|
DOI |
https://doi.org/10.5281/zenodo.5662712 |
|
persistent identifier |
https://treatment.plazi.org/id/B10D2F47-CC3B-FFA0-FF1E-FD8CFEA574AF |
|
treatment provided by |
Plazi |
|
scientific name |
Simulium litobranchium Hamada, Pepinelli, Mattos-Gloria & Luz |
| status |
sp. nov. |
Simulium litobranchium Hamada, Pepinelli, Mattos-Gloria & Luz View in CoL New Species
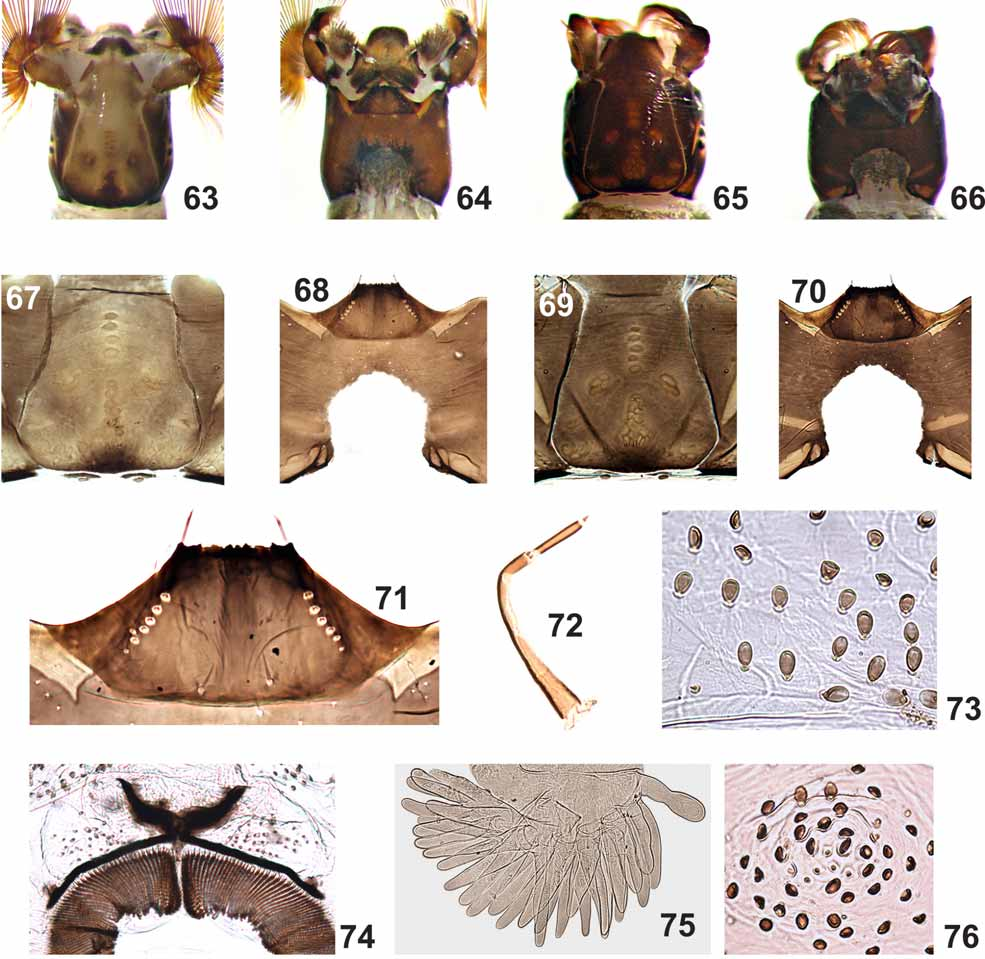
Figs 1–16 View FIGURES 1 – 11 View FIGURES 12 – 16 , 20–32 View FIGURES 17 – 25 View FIGURES 26 – 32 , 35, 38, 41, 44–52 View FIGURES 33 – 44 View FIGURES 45 – 50 View FIGURES 51 – 56 , 57–70 View FIGURES 57 – 58 View FIGURES 59 – 62 View FIGURES 63 – 76
Diagnosis. Female: with internal side of the anal lobe concave in the medial region, forming two short projections of similar width; presence of hair-like setae on the Sc. Male: ventral plate subrectangular, with median anterior projection; apical region of the projection larger than its base; anterolateral region of the ventral plate elongated. Pupa: with 12 short, thick, rigid filaments, without spicules in annular arrangement, with tips pointed and sclerotized; thorax without tubercles on the dorsal anterior region; dorsal posterior region with tubercles mostly pointed. Larva with paired dorsal abdominal tubercles, varying from weak to strongly developed on the 1st to the 5th or 6th segment; gill after histoblast dissection, without spicules in annular arrangement.
FEMALE ( Figs 1–16 View FIGURES 1 – 11 View FIGURES 12 – 16 ). General color black; body mean length (from anterior region of head to abdomen tip) 1.8 mm (SD = 0.08, n = 5); lateral thorax mean length (from neck to wing base) 0.6 mm (SD = 0.04, n = 5). Wing mean length 2.1 mm (SD = 0.1, n = 11); mean width 1.0 mm (SD = 0.03, n = 11).
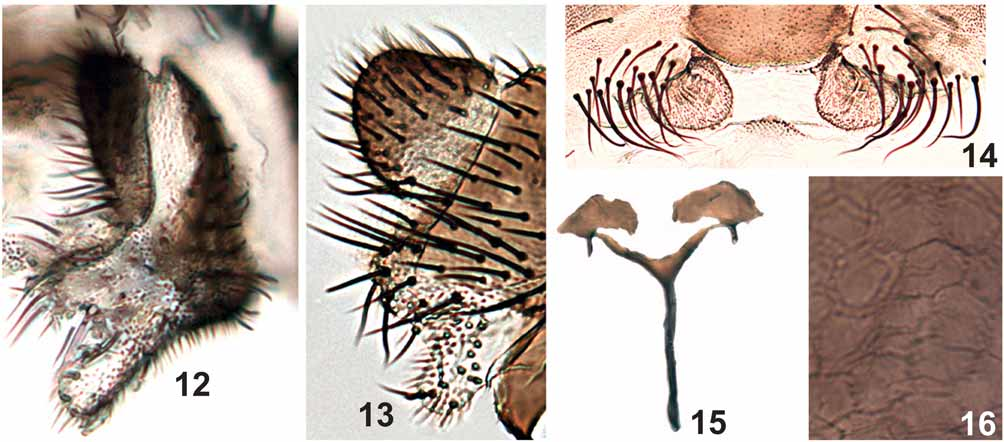
Frons brown, covered with short, golden hairs; clypeus brown with silver pruinosity, covered with short, golden hairs; frons longer than wide; fronto-ocular suture absent, fronto-ocular triangle as in Figure 4 View FIGURES 1 – 11 . Antenna mean length 0.53 mm (SD = 0.03 n = 7); scape, pedicel and first flagellomere brownish yellow, subsequent flagellomeres increasingly dark brown. Palpus dark brown; sensory vesicle length less than half the length of palpomere III ( Fig. 5 View FIGURES 1 – 11 ), palpomere V 2.6 times as long as palpomere III and 2.2 times as long as palpomere IV ( Fig. 5 View FIGURES 1 – 11 ). Mandible with 5–10 (mean = 8.1, SD = 2.0, n = 9) weak external serrations and 18–31 (mean = 29.1, SD = 4.5, n = 9) internal teeth. Lacinia with 24–29 (mean = 26.4, SD = 1.5, n = 9) retrorse teeth. Cibarium with sclerotized cornuae ( Fig. 3 View FIGURES 1 – 11 ); area between the cornuae sclerotized with small serrations. Scutum black, covered with golden hairs, distributed in small groups ( Figs 1, 2 View FIGURES 1 – 11 ). Anepisternum and katepisternum brown with silver pruinosity. Scutellum brown, with gold and brown hairs; postnotum brown, with silver pruinosity. Wing Costa with spiniform and hair-like setae, Sc and R with hair-like setae ( Fig. 6 View FIGURES 1 – 11 ). Foreleg ( Fig. 7 View FIGURES 1 – 11 ) with coxa, trochanter and femur yellowish brown; tibia with external margin whitish, except 1/5 distal area that is dark, internal margin brown; basitarsus and tarsomeres I–IV dark brown. Middle leg ( Fig. 8 View FIGURES 1 – 11 ) with coxa and trochanter yellowish brown; femur and tibia yellowish brown with distal region dark brown; basitarsus mostly yellowish brown, except a small portion in the distal end dark brown; tarsomere I–IV dark brown. Hind leg ( Fig. 9 View FIGURES 1 – 11 ) coxa, trochanter and femur brown, tibia with basal 1/3 whitish and 2/3 dark brown; and basal 3/4 of basitarsus whitish, distal 1/4 dark brown; tarsomeres I–IV dark brown; calcipala longer than wide, reaching pedisulcus ( Fig. 10 View FIGURES 1 – 11 ). Tarsal claws curved, without subbasal tooth ( Fig. 11 View FIGURES 1 – 11 ). Femora and tibiae of the middle and hind leg with narrow scale-like setae and thin setae. Basal fringe, in the abdomen with long golden-whitish and brown hairs. Tergite II with lateral silver pruinosity; tergites VI–VIII with varnish-like dorsal plate. Cercus subtriangular, anal lobe with internal area concave medially, forming two short projections, anterior projection longer than the posterior one ( Figs 12, 13 View FIGURES 12 – 16 ). Hypogynial valves ( Fig. 14 View FIGURES 12 – 16 ) not sclerotized, rounded, with microtrichias. Genital fork ( Fig. 15 View FIGURES 12 – 16 ) with stem long, lateral arms well developed forming subtriangular space; lateral plate well developed ( Fig. 15 View FIGURES 12 – 16 ) and apodemes strong. Spermatheca subspherical, with cuticular microspines distributed in nearly pentagonal pattern ( Fig. 16 View FIGURES 12 – 16 ); spermathecal duct and area of attachment unpigmented.
MALE ( Figs 20–32 View FIGURES 17 – 25 View FIGURES 26 – 32 , 35, 38, 41, 44 View FIGURES 33 – 44 ). General body color black, body mean length, 1.7 mm (SD = 0.14, n = 5); lateral thorax mean length (from neck to anterior region of wing insertion) 0.5 mm (SD = 0.06, n = 5). Wing mean length 2.0 mm (SD = 0.09, n = 10), mean width 1.0 mm (SD = 0.025, n = 10).
Antenna ( Fig. 27 View FIGURES 26 – 32 ) mean length 0.51 mm (SD = 0.04 mm, n = 5); scape and pedicel brownish yellow, subsequent flagellomeres increasingly dark brown. Palpus ( Fig. 28 View FIGURES 26 – 32 ) brown, palpomere V about 2.4–3.5 times as long as palpomere III and 2.2–2.8 times as long as palpomere IV; sensory vesicle small, subspherical, 1/4– 1/5 the length of palpomere III length. Scutum black ( Figs 20–25 View FIGURES 17 – 25 ), covered with golden hairs and a pair of dorso-lateral silver spots ( Figs 20–25 View FIGURES 17 – 25 ). Anepisternum and katepisternum brown with silver pruinosity.
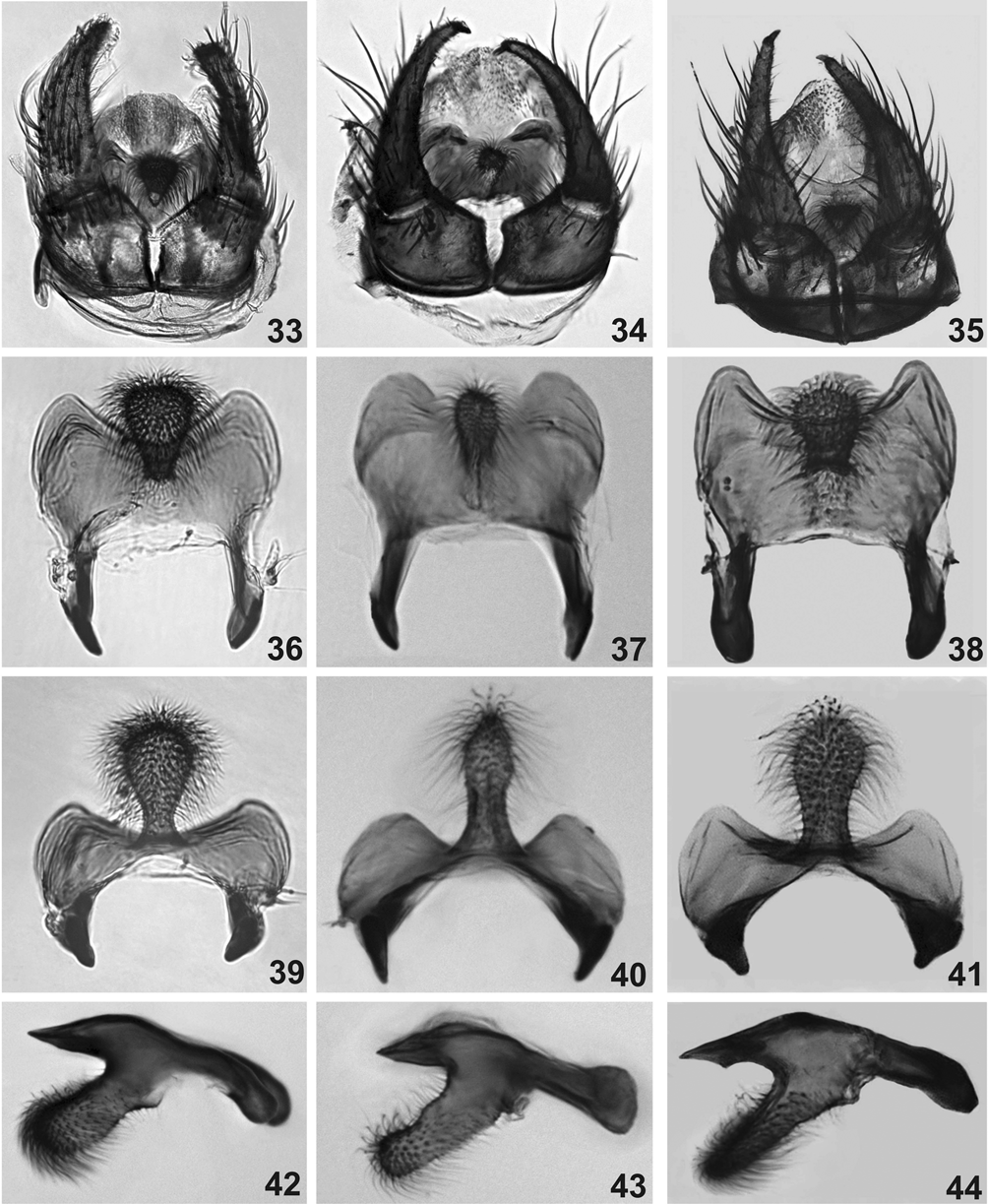
Scutellum brown with thin golden hairs. Postnotum brown with silver pruinosity. Wing similar to the female, except in Sc, which is mostly bare ( Fig. 26 View FIGURES 26 – 32 ); only one of 12 males examined had hair-like setae on Sc. Legs with same color pattern as female, except that, in general, the male legs are darker. Halter yellow. Abdominal basal fringe with thin, long brown hairs; tergites II, V–VII with lateral silver pruinosity. Gonocoxite and gonostylus ( Figs 31 View FIGURES 26 – 32 , 35 View FIGURES 33 – 44 ) brown, gonocoxite wider than long; gonostylus more than twice the gonocoxite length, longer than wide, bearing one spinule at the apex ( Figs 31, 32 View FIGURES 26 – 32 ). Ventral plate ( Figs 38, 41, 44 View FIGURES 33 – 44 ), in ventral view, subrectangular, with a median anterior projection enlarged distally and covered with setae. Median sclerite ( Fig. 29 View FIGURES 26 – 32 ) subrectangular, with mid-apical region lightly sclerotized. Paramere weakly sclerotized, poorly developed and aedeagal membrane with spicules as in Figure 30 View FIGURES 26 – 32 .
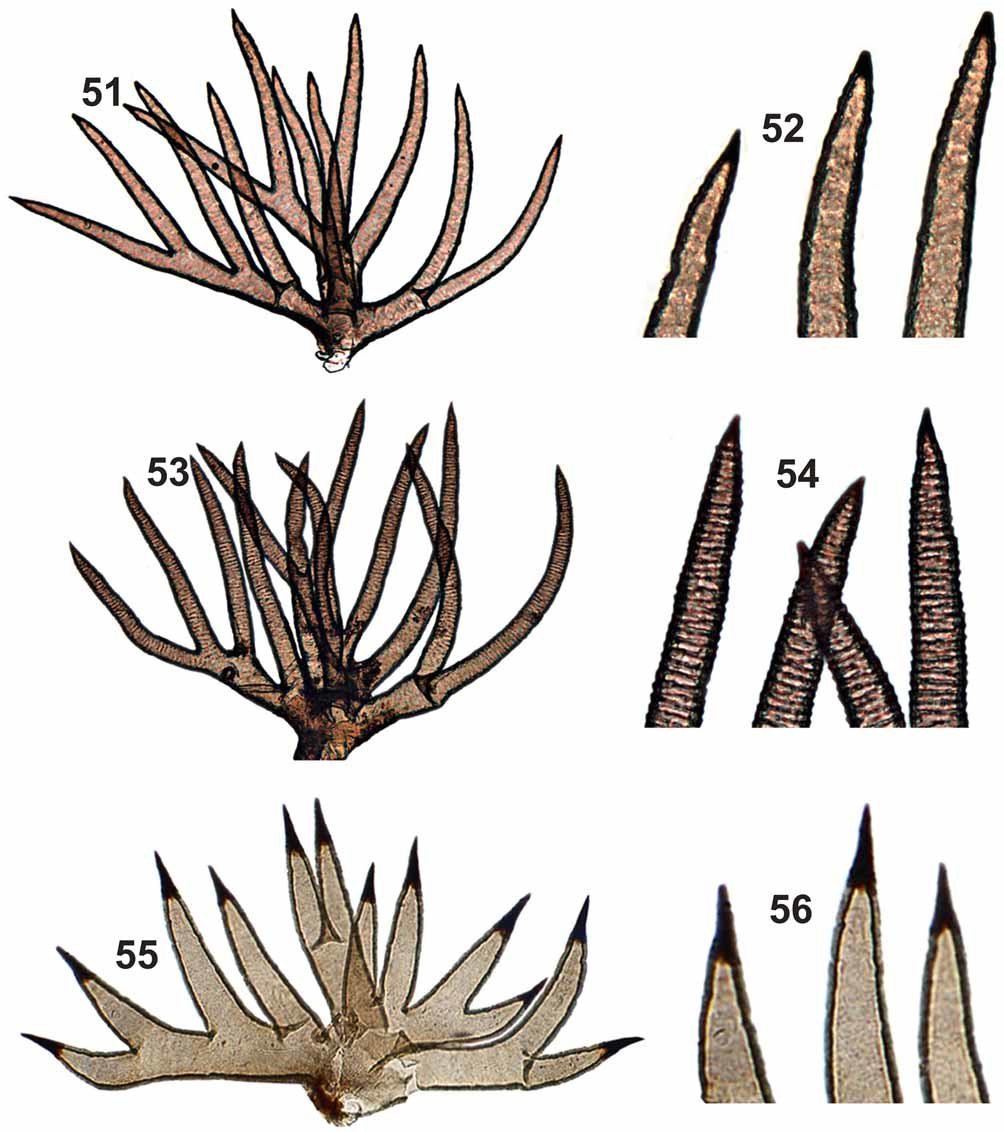
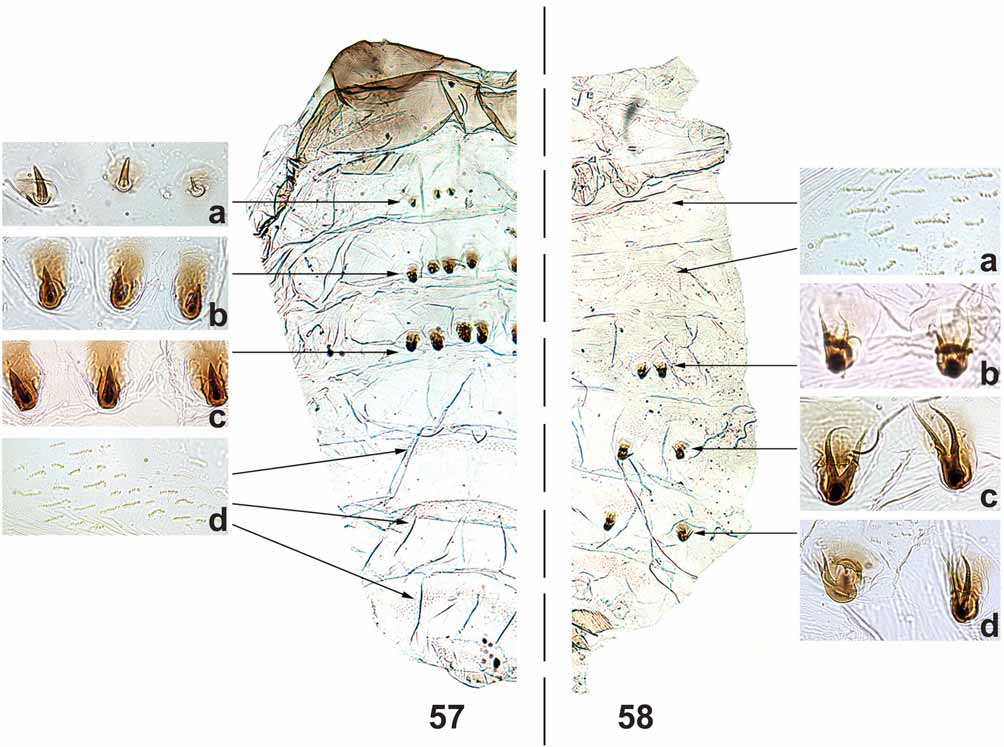
PUPA ( Figs 45–52 View FIGURES 45 – 50 View FIGURES 51 – 56 , 57, 58 View FIGURES 57 – 58 ). Mean body length 3.0 mm (SD = 0.2, n = 5). Cocoon ( Fig. 45 View FIGURES 45 – 50 ) boot-shaped, thick and hard, without central projections ( Fig. 46 View FIGURES 45 – 50 ). Mean length along cocoon dorsal surface 2.0 mm (SD = 0.2, n = 5). Head, cephalic plate with 1 + 1 short, thick, simple frontal trichome ( Fig. 48 View FIGURES 45 – 50 ) and 2 + 2 short, thick, simple dorsal trichomes ( Fig. 49 View FIGURES 45 – 50 ). Round tubercles present on the ventral region of cephalic plate, absent on the dorsal region. Thorax without tubercles on the dorsal anterior region; dorsal posterior region, with majority of tubercles pointed, but some rounded in the area below the tracheal trunk of gill filaments. Thorax with 5 + 5 short, thick simple trichomes ( Fig. 50 View FIGURES 45 – 50 ), rarely 1 + 1 bifid; 1 + 1, thick, longer lateral trichome. Gills with 12 short, thick, rigid filaments, distributed in a three-dimensional pattern with tips pointed and sclerotized, without spicules in annular arrangement ( Figs 51, 52 View FIGURES 51 – 56 ). Abdomen as in Figures 57 and 58 View FIGURES 57 – 58 . Tergite I with 1 + 1 sublateral setae. Tergite II with 4 + 4 stout setae ( Fig. 57 View FIGURES 57 – 58 a), 2 + 2 thin sublateral setae and many small, tubercles distributed, especially, in the anteromedian region of the tergite. Tergites III and IV each with 4 + 4 anteriorly directed hooks on posterior margin ( Figs 57 View FIGURES 57 – 58 b, 57c). Tergites VI–IX with comb-like groups of fine posteriorly directed spines on anterior margin ( Fig. 57 View FIGURES 57 – 58 d). Sternites III–VIII with anterior medial comb-like groups of microspines ( Fig. 58 View FIGURES 57 – 58 a). Sternites V–VII with 2 + 2 stout, bifid hooks ( Figs 58 View FIGURES 57 – 58 b, 58c, 58d).
LARVA (last instar) ( Figs 59–76 View FIGURES 59 – 62 View FIGURES 63 – 76 ). Body mean length 4.3 mm (SD = 0.2, n = 5); head capsule, lateral mean length 0.4 mm (SD = 0.02, n = 5).
General coloration varying from light to dark grayish green ( Figs 59–62 View FIGURES 59 – 62 ) (in Carnoy's solution). Head cuticle with small simple setae. Frontoclypeal apotome, in light colored larvae, with positive pattern as in Figure 63 View FIGURES 63 – 76 and, in dark colored larvae, with negative pattern as in Figure 65 View FIGURES 63 – 76 . Cervical sclerites small, eliptical, free in the membrane, each with a thin, elongated sclerite anteriorly ( Figs 67, 69 View FIGURES 63 – 76 ). Postgenal cleft widest at medial region and with anterior end subtriangular ( Figs 68, 70 View FIGURES 63 – 76 ), area inside cleft irregularly pigmented ( Figs 64, 66 View FIGURES 63 – 76 ). Postgenal bridge 0.3–0.6 times as long as hypostoma ( Figs 68, 70 View FIGURES 63 – 76 ). Antenna as long as labral fan stalk; distal and proximal articles smaller than medial (proportions of articles, proximal to distal, excluding cone sensillum, 1:1.4– 1.75: 0.8–0.9) ( Fig. 72 View FIGURES 63 – 76 ). Labral fan with 42–56 primary rays (Mean = 49, SD = 5, n = 7). Hypostoma with pigmented anterior margin ( Fig. 71 View FIGURES 63 – 76 ) with median tooth and sublateral teeth not well differentiated, 2 + 2 sublateral teeth with 1 – 2 small serrations; 1 + 1 strong lateral tooth and 1 – 2 paralateral teeth, with 5 – 7 sublateral setae per side. Mandibular teeth: one apical, two small external; three subapical; 6 – 7 internal teeth; one mandibular serration and one small mandibular sensillum ( sensu Craig & Craig, 1986). Gill histoblast dissected with 12 smooth filaments, with sclerotized and pointed apex. Body covered with ovoid setae ( Fig. 73 View FIGURES 63 – 76 ); ventral tubercles absent ( Figs 59–62 View FIGURES 59 – 62 ). Abdomen with 1 + 1 dorso-lateral tubercles ( Fig. 76 View FIGURES 63 – 76 ), on segments 1 – 5 ( Figs 59–61 View FIGURES 59 – 62 ), or 1 – 6 ( Fig. 62 View FIGURES 59 – 62 ), varying from well to poorly developed ( Figs 59–62 View FIGURES 59 – 62 ), especially in the population from Minas Gerais. Anterodorsal arms of anal sclerite 0.5 times the length of posteroventral arms ( Fig. 74 View FIGURES 63 – 76 ), presence of thin and few enlarged setae near the arms. Posterior circlet bearing 139–166 rows (Mean = 153, SD = 10, n = 9) with 22–26 hooks (Mean = 24, SD = 2, n = 9). Rectal papillae with three branches, each with approximately 18 – 31 digitiform lobes ( Fig. 75 View FIGURES 63 – 76 ); mean number (±SD) on each branch = 24(±3) + 24(±2) + 28 (±4), n = 9.
TYPES. Holotype – male (M), pinned, collected in the Ponte de Pedra River, Montividiu municipality, Goiás, 17o10’S, 50o50’W, 671 masl, collectors N. Hamada, M. Pepinelli & V. Landeiro, 21/05/2006 (INPA). Paratypes - same locality, date and collectors as holotype, 8 pupae (P) and 9 larvae (L) in 80% ethanol; 5 P and 8 L mounted on slide; 3 females (F) and 5 M pinned with their pupal exuviae in glycerine; 5 M and 2F mounted on slide together with its pupal exuviae (INPA). Same locality as holotype, collectors N. Hamada & M. Pepinelli, 15/ 07/2004, 2 F and 3 M (INPA). Same locality, date and collectors as holotype, 8 P and 9 L in 80% ethanol, 3 F and 4 M pinned with their pupal exuviae in glycerine (MZUSP). Montividiu River, Montividiu municipality, Goiás, 17o26’S, 51o10’W, 779 masl., 21/05/2006, collectors N. Hamada, M. Pepinelli & V. Landeiro, 21/05/2006, 2 F, pinned with its pupal exuviae (INPA). Santo Antônio River, Delfinópolis municipality, Minas Gerais, 20o15’S, 46o51’W, 726 masl, collectors N. Hamada & M. Pepinelli, 18/09/2005 (INPA), 15 P and 10 L in 80% ethanol; 1 P and 5 L mounted on slide, 4 F and 5 M, pinned with their pupal exuviae (INPA); 15 P and 10 L in 80% ethanol, 4 F and 5 M, pinned with their pupal exuviae (MZUSP).
Etymology: The species name derived from lito (G) = smooth and branchium (G) = gill, in reference to smooth appearance of the gill filaments due to the lack of spicules in annular arrangements.
Taxonomic discussion. The new species described in this paper can be placed in the subgenus Thyrsopelma or Trichodagmia , depending on which black fly classification scheme is being followed. This situation occurs because the validity of the subgenus Thyrsopelma Enderlein is debated (e.g. Miranda-Esquivel & Coscarón 2001, Shelley et al. 1997). Thyrsopelma is considered synonymous with Trichodagmia Enderlein in the inventory of world black flies by Adler & Crosskey (2009). In this study we are following the Miranda-Esquivel & Coscarón (2001) classification scheme.
Female scutal pattern of S. litobranchium n. sp. is similar to most of the other described species in S. ( Thyrsopelma ); the exceptions are S. scutistriatum and S. perplexum , but the new species can be distinguished from these by the shape of the anal lobe ( Figs 12, 13 View FIGURES 12 – 16 ). The only species from which it cannot be distinguished based on this character is S. duodenicornium Hamada, Pepinelli & Trivinho-Strixino , which also has the internal side of the anal lobe concave in the medial region, forming two short projections of similar width. However, the female of the new species can be distinguished from that of S. duodenicronium , among other characters, by the presence of hair-like setae on the Sc, by the shape of the tarsal claws (which are shorter and more curved than those of S. duodenicornium ) and by having fronto-occular triangle longer and more pointed than that of S. duodenicornium . The female of S. jeteri (Py-Daniel, Darwich, Mardini, Strieder & Coscarón) was incompletely described ( Py-Daniel et al. 2005); therefore, morphological comparisons are not possible.
Male of S. litobranchium n. sp. is similar to most of the males in the S. ( Thyrsopelma ) subgenus, with the exception of S. duodenicornium , S. scutistriatum and S. perplexum , which do not have silver spots on the scutum. The male of the new species and those of S. itaunense , S. orbitale and S. duodenicornium can be differentiated from the male of Simulium guianense based on the fact that the latter has a more elaborated silver pattern on the scutum ( Figs 17–19 View FIGURES 17 – 25 ), while in the other species, the pattern is basically a pair of lateral silver spots ( Figs 20–25 View FIGURES 17 – 25 ). Males of S. jeteri were incompletely described ( Py-Daniel et al. 2005); therefore, morphological comparisons are not possible.
The male genitalia of S. guianense , S. duodenicornium and S. litobranchium n. sp. are very similar. However, morphological difference can be visualized in the ventral plate ( Figs 33–44 View FIGURES 33 – 44 ). Pepinelli et al. (2005) mistakenly stated that the ventral plates of S. duodenicornium and S. guianense , in Figures 27 and 28 View FIGURES 26 – 32 were shown in dorsal view; instead, the ventral plates in the above-cited figures were shown in ventral view tilted dorsally. According to Crosskey (1990), due to the diversity of shapes of this structure in Simuliidae in general and the similarity of the characteristics of this plate between related species, the ventral plate is one of the best characters for distinguishing species and groups of species. The S. litobranchium n. sp. ventral plate can be distinguished from that of S. guianense by the width of the apical region of the projection, which is larger in the former than in the latter species ( Figs 37, 38 View FIGURES 33 – 44 ). But this structure is very similar in the new species and in S. duodenicornium ( Figs 39, 41 View FIGURES 33 – 44 ); the only difference can be visualized in the anterolateral region of the ventral plate, which is more elongated in the new species than in S. duodenicornium ( Figs 39, 41 View FIGURES 33 – 44 ).
At the pupal stage, the number, shape and disposition of the gill filaments of S. litobranchium n. sp. are similar to those of five species in the S. ( Thyrsopelma ): S. guianense , S. perplexum , S. duodenicornium , S. scutistriatum and S. hirtipupa . However, except for S. duodenicornium and S. scutistriatum , all of these species have spicules or projections in an annular arrangement on the gill filament surface ( Shelley et al. 1989, Shelley et al. 1997, Pepinelli et al. 2005). Pupae of the new species can be distinguished from those of S. hirtipupa because the latter has a cocoon with irregular protuberances and a large number of spines on the cephalic and thoracic region. The pupa of new species can be distinguished from that of S. scutistriatum by the cocoon shape of the latter species, which has the anteroventral margin more elongated; additionally, the latter species has a cephalic plate with several accuminated tubercles. The pupa of the new species can be distinguished from that of S. duodenicornium by the presence, in the latter species, of small tubercles distributed evenly on tergite II and by the width of the gill filaments, which are thicker basally in the latter species ( Pepinelli et al. 2005).
The larvae of S. litobranchium n. sp. are similar to those of most species in the subgenus S. ( Thyrsopelma ), with ovoid setae covering the body cuticle (exceptions are S. scutistriatum and S. perplexum ). The new species has paired dorsal tubercles, varying from weakly to strongly developed on the 1st to the 5th or 6th abdominal segment (some larvae do not have the paired tubercles on the 6th segment). The other characteristic that can be used to easily distinguish the new species from the other in S. ( Thyrsopelma ) is the gill histoblast of last-instar larvae: after dissection, the same gill pupal characteristic can be observed.
The synonyms of Simulium pintoi d’Andretta & d’Andretta and S. ortizi Ramírez-Pérez with S. guianense and of S. brasiliense (Enderlein) with S. orbitale are well established and widely accepted ( Adler & Crosskey, 2009). Simulium pintoi and S. ortizi can be distinguished from the new species using the same characters used to distinguish it from S. guianense , especially female genitalia and male scutal pattern.
Simulium brasiliense was sucinctly described based on the female and was established as a synonym of S. orbitale ; since the anal lobe of S. litobranchium n. sp. has its internal side concave in the medial region, forming two short projections of similar width it cannot be misidentified with the species cited above because the females of these species have a tail-like projection on the membranous part of the anal lobe. Also, the hypogynial valves are rounded in the new species while in the other species cited above they are subovoidal, longer than wide.
DNA barcoding. Simuliidae is a taxonomically challenging family because these insects are small in size and structural homogeneous, and because of the presence of reproductively isolated (but morphologically indistinguishable) sibling species ( Adler et al. 2004). For this reason, knowledge about all life stages is important, and the use of a multifaceted approach incorporating both morphological and DNA-sequence data, such as DNA barcoding, is critical for species recognition.
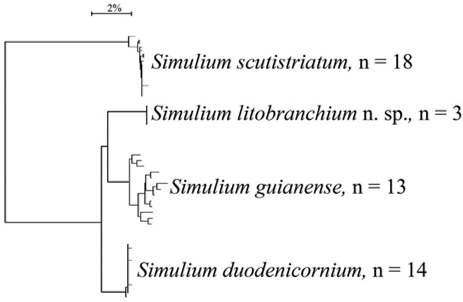
In the present study, DNA barcoding discriminated species within the subgenus S. ( Thyrsopelma ) ( Fig. 77 View FIGURE 77. A ). Mean intraspecific genetic divergence for the four species analyzed was 0.51%. Simulium guianense had a maximum intraspecific divergence of more than 2.5% ( Table 1 View TABLE 1 ). According to Rivera & Currie (2009), in the Nearctic region, the mean intraspecific divergence for 58 black fly species was 0.76%, and the maximum intraspecific divergence value was 3.84% [observed in Simulium rostratum (Lundström) ]. The high divergence (> 4%) in the sequences of the COI gene (Table 2) among the four species we analyzed suggests that they are different species; DNA barcoding, therefore, corroborates the morphological data we provided to establish that Simulium litobranchium n. sp. is not a variant of S. guianense , as indicated by Shelley et al. (2002).
Bionomics. The Paraná River, which is the main water course in this hydrographic basin, has as major tributaries, among others, the Rio Grande, Paranaíba, Tietê, Paranapanema and Iguaçu rivers. Despite the heavy anthropogenic impact caused by agriculture in the case of the two rivers in Goiás state, Simulium litobranchium n. sp. was collected in abundance. Simulium litobranchium n. sp. larvae and pupae were collected in three rivers, varying in width from 30 to 40 meters, two in Goiás (Paranaíba River basin) and one in Minas Gerais state (Rio Grande basin). Simulium guianense was also collected in these same hydrographic basins, but in larger tributaries (more than 150 m in width). The streams where the new species was collected had a streambed composed of sand, small stones and boulders. The water temperature ranged from 17 to 19o C, pH from 6.5 to 7.3, and electrical conductivities were low (below 15µ/cm). Larvae and pupae were collected on leaves of plants in the family Podostemacea, submerged tree branches and leaves of riparian vegetation and leaves of submerged grass. The female of the new species was not collected biting humans during the fieldwork.





No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |