
Results
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4752.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:A03E9412-BBE8-44DE-B6B9-325017735BFB |
|
DOI |
https://doi.org/10.5281/zenodo.4323676 |
|
persistent identifier |
https://treatment.plazi.org/id/B13C87BC-9961-E377-FF70-491E350BFD2B |
|
treatment provided by |
Felipe |
|
scientific name |
Results |
| status |
|
Results View in CoL View at ENA
Morphological description
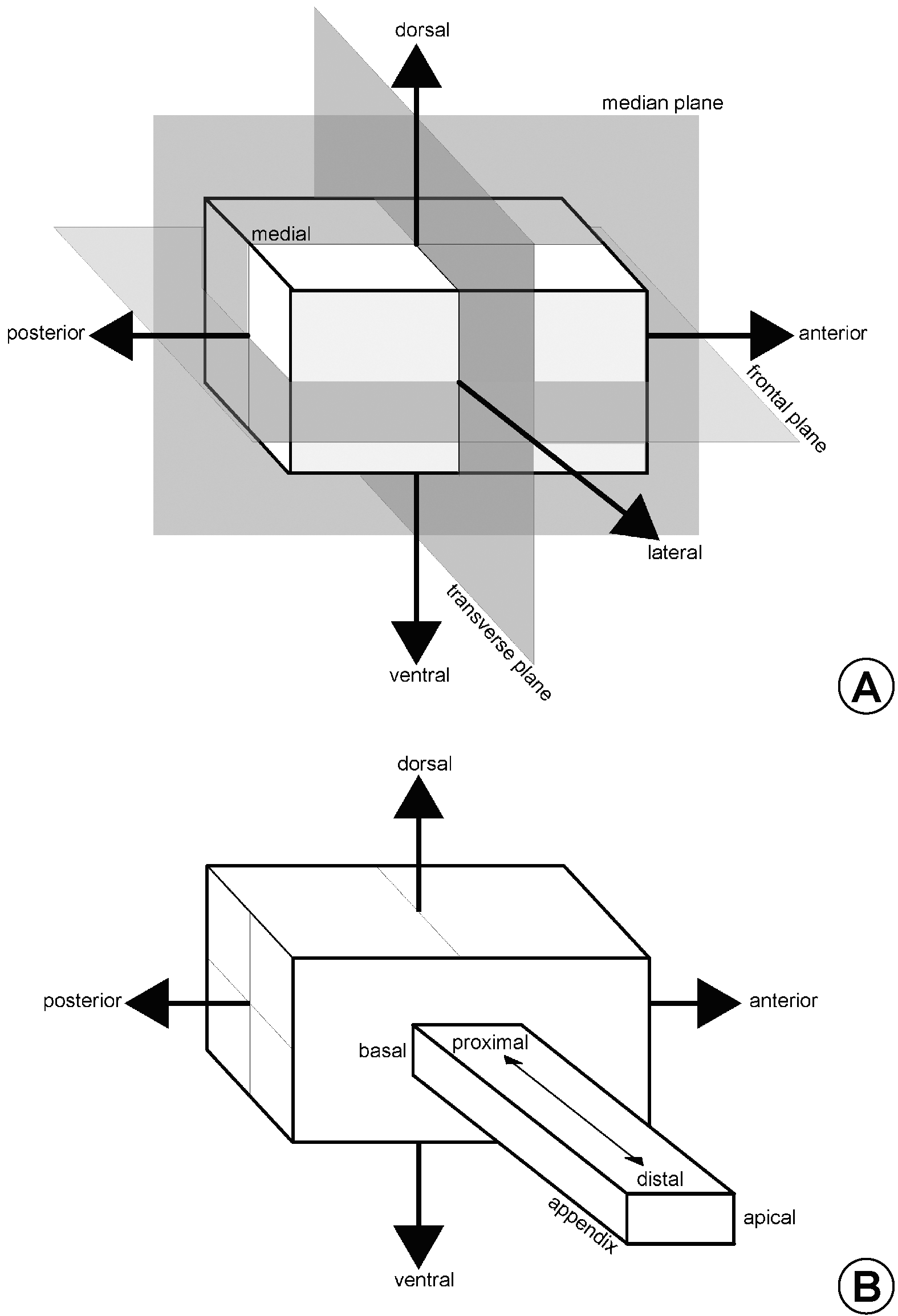
Head. The head (hd: Figs 5–8 View FIGURE 5 View FIGURE 6 View FIGURE 7 View FIGURE 8 ) of Bethylidae is usually prognathous with the mouthparts placed at the anterior extremity ( Fig. 6 View FIGURE 6 ). Thus, the anterior, posterior, dorsal and ventral surfaces correspond to the ventral, dorsal, anterior and posterior in hypognathous insects, respectively.
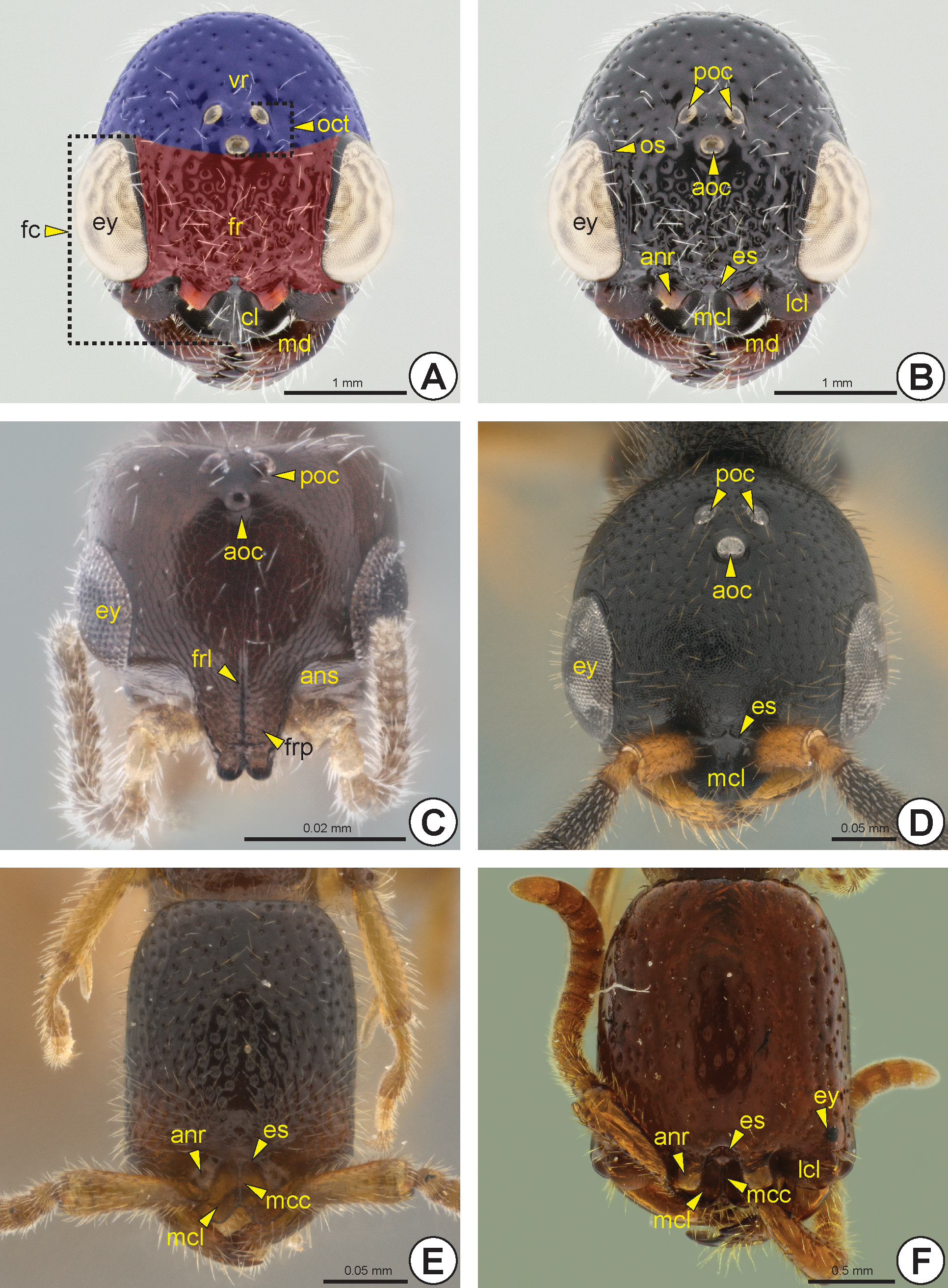
The face (fc: Fig. 5A View FIGURE 5 ) is the dorsal surface of the head between the eyes (ey: Figs 5 View FIGURE 5 A–D, 5F, 6) and anterior ocellus (aoc: Figs 5 View FIGURE 5 B–D, 6B, 6D), and including the clypeus (cl: Fig. 5A View FIGURE 5 ). The frons (fr: Fig. 5A View FIGURE 5 ) is the area of the face located between the anterior ocellus and the antennal foramina (anf: Figs 6A, 6B, 6F View FIGURE 6 , 7D View FIGURE 7 ). The antennal foramen is adjacent to the posterior margin of the clypeus, thus the frons generally occupies the whole area of the
face, except the clypeus. The definitions of the face and frons are not strictly applicable in specimens without an ocellar triangle (oct: Fig. 5A View FIGURE 5 ). A pair of anterior tentorial pits (atp: Fig. 6A View FIGURE 6 ) is visible near the anterior margin of the face; these pits are represented internally by the anterior tentorial arms (ata: Fig. 7D View FIGURE 7 ).
The frons usually has a longitudinal median line, the frontal line (frl: Figs 5C View FIGURE 5 , 8D View FIGURE 8 ), which can be represented by a furrow or a carina. When complete, the line arises medially from the posterior clypeal margin and reaches the anterior ocellus.
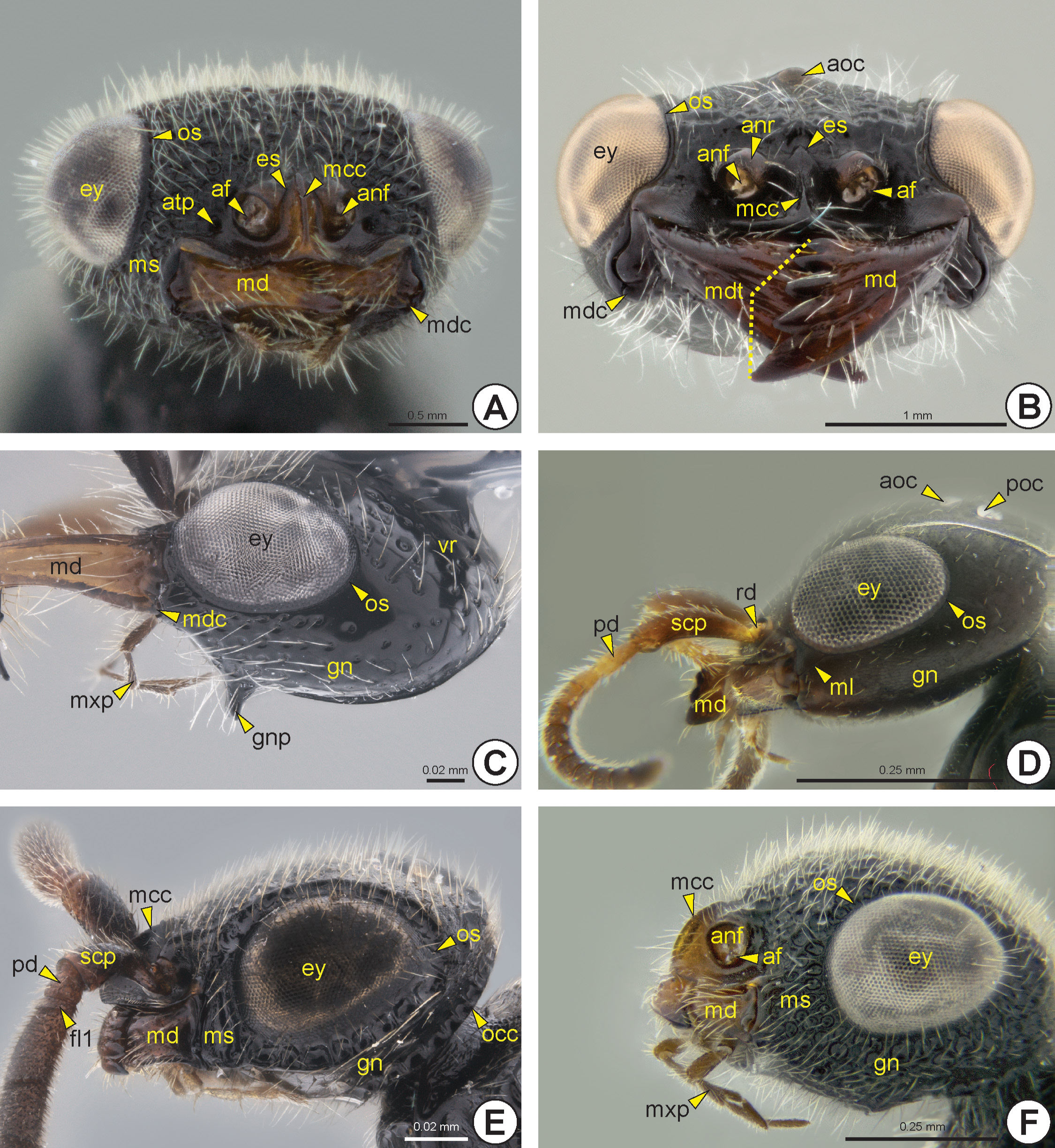
In most of the Bethylidae , the eye is adjacent to the mandible (md: Figs 5A, 5B View FIGURE 5 , 6 View FIGURE 6 , 7B View FIGURE 7 , 8D View FIGURE 8 ) and the malar space (ms: Figs 6A, 6E, 6F View FIGURE 6 ) is reduced or almost absent ( Figs 5A, 5B View FIGURE 5 , 6C View FIGURE 6 ). The malar space of some species has a longitudinal line connecting the anterior margin of the eye and the mandible, the malar line (ml: Fig. 6D View FIGURE 6 ).
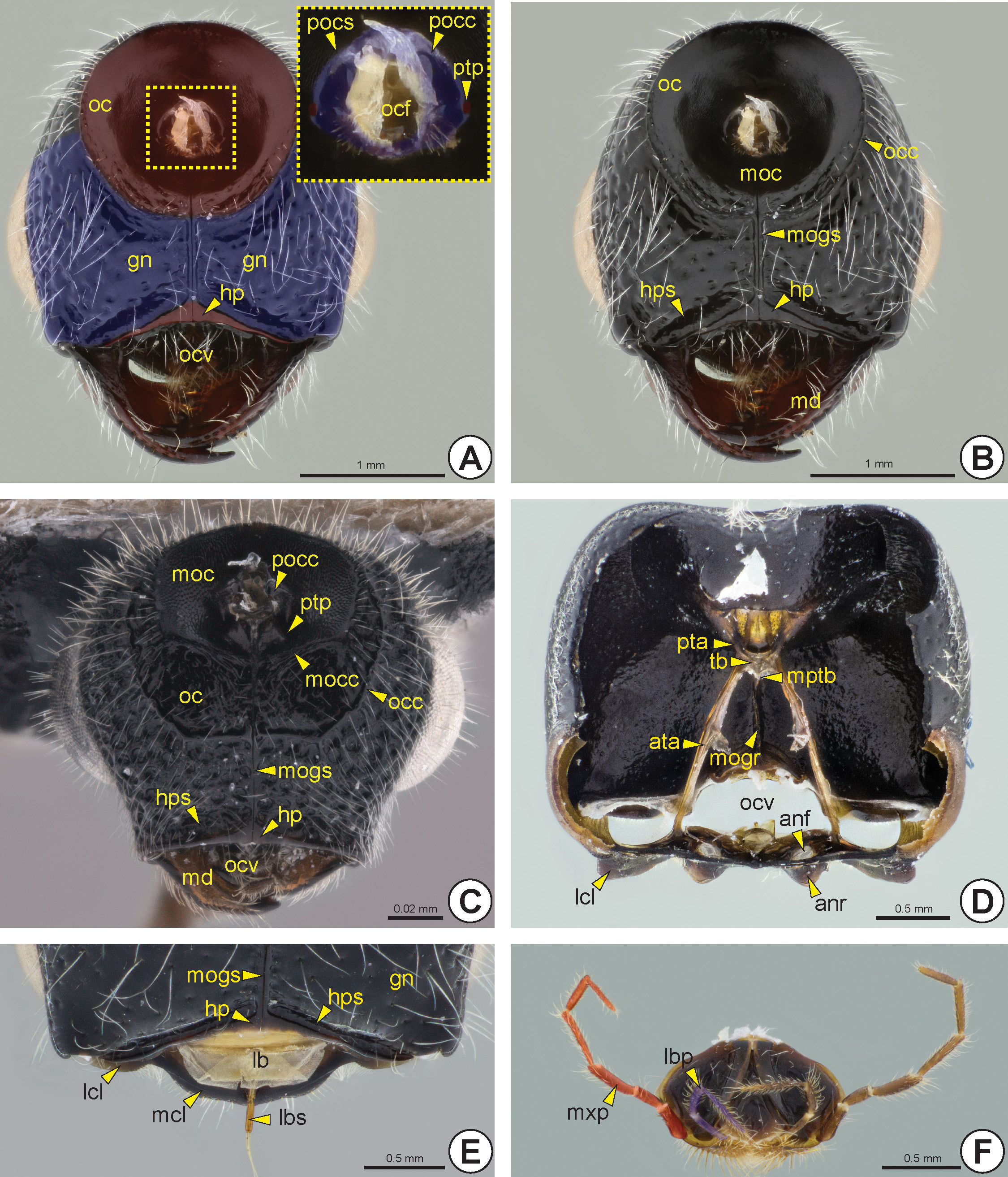
The gena (gn: Figs 6 View FIGURE 6 C–F, 7A) is the area of the head lateral to the vertex (vr: Fig. 5A View FIGURE 5 ) and between the lateral margin of the eye and the occipital carina (occ: Figs 6E View FIGURE 6 , 7B, 7C View FIGURE 7 ) and hypostoma (hp: Figs 7 View FIGURE 7 A–C, 7E). A few species have an angled projection on each gena, the genal process* (gnp: Fig. 6C View FIGURE 6 ). The occiput (oc: Figs 7 View FIGURE 7 A–C) is a circular and concave area that surrounds the occipital foramen (ocf: Fig. 7A View FIGURE 7 ). In most of the Bethylidae the occiput is delimited by the occipital carina. Mesad on the occiput, there is a circular area that is generally more concave and has a different texture from the rest of the occiput, the mesocciput * (moc: Figs 7B, 7C View FIGURE 7 ); it can be delimited by the mesoccipital carina* (mocc: Fig. 7C View FIGURE 7 ).
The vertex is the area located between the frons and the occiput, so the vertex is delimited anteriorly by the anterior margin of the anterior ocellus, laterally by the posterior margin of the eye, and posteriorly by the dorsal sector of the occipital carina. Ventrally, the gena and occiput present a median longitudinal line marking ventral
sclerotization in the head, the medioccipito-genal suture* (mogs: Figs 7B, 7C, 7E View FIGURE 7 ), which usually is carinate and continuous to the hypostoma. The medioccipito-genal suture is internally represented by a high medioccipito-genal ridge * (mogr: Fig. 7D View FIGURE 7 ). The hypostoma extends throughout the ventral margin of the oral cavity (ocv: Figs 7A, 7C, 7D View FIGURE 7 ) and is generally raised and more polished than the genal surface. Posteriorly, the hypostoma is usually delimited by the hypostomal sulcus (hps: Figs 7B, 7C, 7E View FIGURE 7 ). A pair of posterior tentorial pits (ptp: Figs 7A, 7C View FIGURE 7 ) is located anterolateral to the postocciput (pocc: Figs 7A, 7C View FIGURE 7 ) and they are represented internally by the posterior tentorial arms (pta: Fig. 7D View FIGURE 7 ). The posterior tentorial arms are very short and connected to the anterior arms by the tentorial bridge (tb: Fig. 7D View FIGURE 7 ). This bridge is complete and has a median projection, the median process of the tentorial bridge (mptb: Fig. 7D View FIGURE 7 ). The postocciput is strongly projected and the postoccipital suture (pocs: Fig. 7A View FIGURE 7 ) is poorly represented by a sulcus.
The clypeus is posteriorly delimited by the epistomal suture (es: Figs 5B, 5 View FIGURE 5 D–F, 6A, 6B), which can be represented by a furrow or carina. The clypeus varies in size and shape; it may have a continuous anterior margin or be divided into three lobes. The trilobate clypeus presents two lateral clypeal lobes (lcl: Figs 5B, 5F View FIGURE 5 , 7D, 7E View FIGURE 7 ) and one median clypeal lobe (mcl: Figs 5B, 5 View FIGURE 5 D–F, 7E). The median clypeal lobe can have a rounded, angled, serrated or truncated anterior margin. In most Bethylidae , the median clypeal lobe is posteriorly elevated, but in some genera this elevation is continuous to the anterior margin. The median linear elevation of the clypeus can form a sharp Λ shape or a blunt ∩ shape, in frontal view. Besides this elevation, many species have a median clypeal carina (mcc: Figs 5E, 5F View FIGURE 5 , 6A, 6B, 6E, 6F View FIGURE 6 ), which is often continuous with the frontal line. The labrum (lb: Fig. 7E View FIGURE 7 ) is posteroventral to the clypeus and visible only after removal of the labiomaxillary complex (lmc: Fig. 7F View FIGURE 7 ). It is weakly sclerotized and presents one or more labral setae (lbs: Fig. 7E View FIGURE 7 ) emerging from the anterior margin of the labrum.
The eye may vary in size, location and gibbosity. In most species, the eye length is between a quarter and a little more than half the length of the head, but the eyes may be absent ( Fig. 5E View FIGURE 5 ) or extremely reduced in some females ( Fig. 5F View FIGURE 5 ). The ocular suture (os: Figs 5B View FIGURE 5 , 6 View FIGURE 6 ) is usually poorly defined by a groove; only a few species have a strongly sulcate suture ( Figs 6E View FIGURE 6 ). The ocellar triangle is formed by two posterior ocelli (poc: Figs 5 View FIGURE 5 B–D, 6D) and one anterior ocellus. The position of the ocellar triangle, distance between the ocelli and the size of each ocellus are highly variable; ocelli are absent in a few apterous forms ( Figs 5E, 5F View FIGURE 5 ).
The maxillary palp (mxp: Figs 6C, 6F View FIGURE 6 , 7F View FIGURE 7 ) is composed of three to six maxillary palpomeres (mxpp1–6: Fig. 7F View FIGURE 7 ), whereas the labial palp (lbp: Fig. 7F View FIGURE 7 ) has one to three labial palpomeres (lbpp1–3: Fig. 7F View FIGURE 7 ). The mandible varies in length and width, and is generally wider distally; it generally has from one to five apical mandibular teeth (mdt: Figs 6B View FIGURE 6 ); very exceptionally there may be seven apical teeth with toothed upper and lower mandibular margins. In most Bethylidae , the lowermost apical tooth is larger than the remaining teeth. Rarely there is a laminar expansion on the lower border of the mandible, the lower mandibular expansion* (lme: Fig. 8D View FIGURE 8 ). The basal margin of the mandible is sinuous and the mandibular condyle (mdc: Figs 6 View FIGURE 6 A–C) is always well defined.
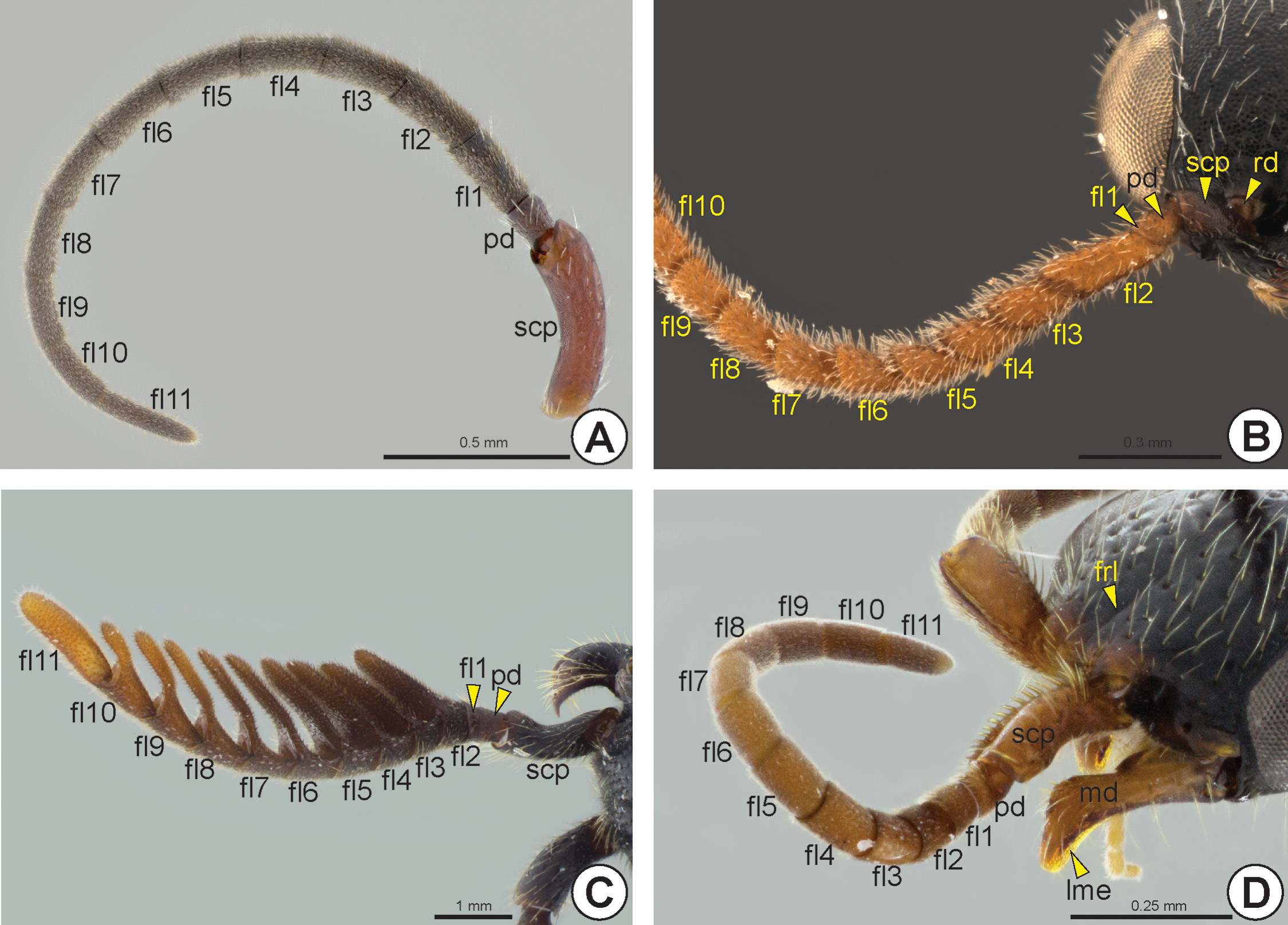
Most Bethylidae have the antenna (an: Fig. 8 View FIGURE 8 ) with 13 antennomeres (ant), but a few have 12 antennomeres. Although the antenna is usually filiform ( Figs 8A, 8D View FIGURE 8 ), some species have pectinate ( Fig. 8C View FIGURE 8 ) or serrated antennae ( Fig. 8B View FIGURE 8 ). The scape (scp: Figs 6D, 6E View FIGURE 6 , 8 View FIGURE 8 ), pedicel (pd: Figs 6D, 6E View FIGURE 6 , 8 View FIGURE 8 ) and proximal flagellomeres (fl1–11: Fig. 8 View FIGURE 8 ) may sometimes have thick and rigid setae ( Fig. 8D View FIGURE 8 ). Some species have the first flagellomere very short and often difficult to detect ( Fig. 6E View FIGURE 6 ). Proximally, the scape exhibits a constriction that delimits the radicle (rd: Figs 6D View FIGURE 6 , 8B View FIGURE 8 ), a rounded basal area of the scape that articulates with the cranial fossa of the scape (cfs) located in the antennifer (af: Figs 6A, 6B, 6F View FIGURE 6 ); it is an articular process located anterolaterally on the margin of the antennal foramen. The slightly depressed area between the antennal foramen and the eye is the antennal scrobe (ans: Fig. 5C View FIGURE 5 ); this area accommodates and facilitates the movement of the antennal scape. In some species, the antennal scrobe is posteriorly delimited by the antennal scrobal carina (ansc). The margins of the antennal foramen can be projected to form the antennal rim (anr: Figs 5B, 5E, 5F View FIGURE 5 , 6B View FIGURE 6 , 7D View FIGURE 7 ). The distance between the antennal foramina varies; in some species these foramina are very close to each other. A few species have a frontal projection* (frp: Fig. 5C View FIGURE 5 ), which overlaps the antennal foramina, extending beyond the clypeus.
Mesosoma .
The mesosoma (msm: Fig. 9) is composed of prothorax (th1: Fig. 9), mesothorax (th2: Fig. 9), metanotum (n3: Figs 9A, 22 View FIGURE 22 ) and the metapectal-propodeal complex (mpc: Fig. 9).
Pronotum: The anterior margin of the pronotum (n1: Figs 10 View FIGURE 10 , 11 View FIGURE 11 ) is projected, forming an anterior and rounded flange, the pronotal flange (n1f: Figs 10 View FIGURE 10 , 11 View FIGURE 11 ). The boundary between the pronotal flange and the rest of the pronotum is usually marked by a transverse pronotal sulcus (n1ts: Figs 10A View FIGURE 10 , 11 View FIGURE 11 A–11C), which is internally represented by the anterolateral pronotal ridge (n1ar). The pronotal flange usually extends laterally on the pronotum. In many species, the dorsal surface forms a distinct area, sometimes delimited by carinae, the dorsal pronotal area (n1da: Figs 10 View FIGURE 10 A–E, 11), which can be trapezoidal, semicircular or bell-shaped, among other forms. The anterior and lateral margins of the dorsal pronotal area can be delimited by the transverse pronotal carina (n1tc: Figs 10A, 10B View FIGURE 10 , 11A, 11C View FIGURE 11 ), posterior to which the pronotal suprahumeral sulcus (n1ss: Figs 10B, 10C View FIGURE 10 , 11A, 11C View FIGURE 11 ) generally extends. The area anterior to the transverse pronotal carina is usually referred to as the pronotal neck (n1n: Fig. 10A View FIGURE 10 ), in which the pronotal flange is included. The pronotal neck rarely presents a distinct epomia (ep: Figs 11A, 11C View FIGURE 11 ), which, when present, is represented by a vertical carina that crosses the pronotal neck, subdividing it into lateral pronotal area (n1la: Fig. 11 View FIGURE 11 ) and cervical pronotal area (n1ca: Figs 10 View FIGURE 10 A–D, 11).
The posterior pronotal margin can have a sulcus or a sequence of foveae, the posterior pronotal sulcus (n1ps: Figs 10A, 10B, 10D View FIGURE 10 ), internally this sulcus corresponds to the posterior pronotal inflection (n1pi: Fig. 10F View FIGURE 10 ) and dorsal pronotal inflection (n1di). Some Bethylidae have a median pronotal line* (n1ml: Figs 10D, 10E View FIGURE 10 ) that can be represented by a carina or a sulcus, this latter is internally represented by the median pronotal ridge* (n1mr: Fig. 10F View FIGURE 10 ).
In some bethylids, the anterolateral corner of the pronotum is very projected, forming the humeral angle (ha: Fig. 10E View FIGURE 10 ). The anterior thoracic spiracle (sp 1t) is located at the posterolateral corner of the pronotum on the bor- der between the pronotum and the mesopleuron (pl2: Fig. 19 View FIGURE 19 ); it is entirely surrounded by sclerotized cuticle and is not visible externally where it is covered by the posterodorsal corner of the pronotum, the pronotal lobe (n1l: Figs 11 View FIGURE 11 B–D).
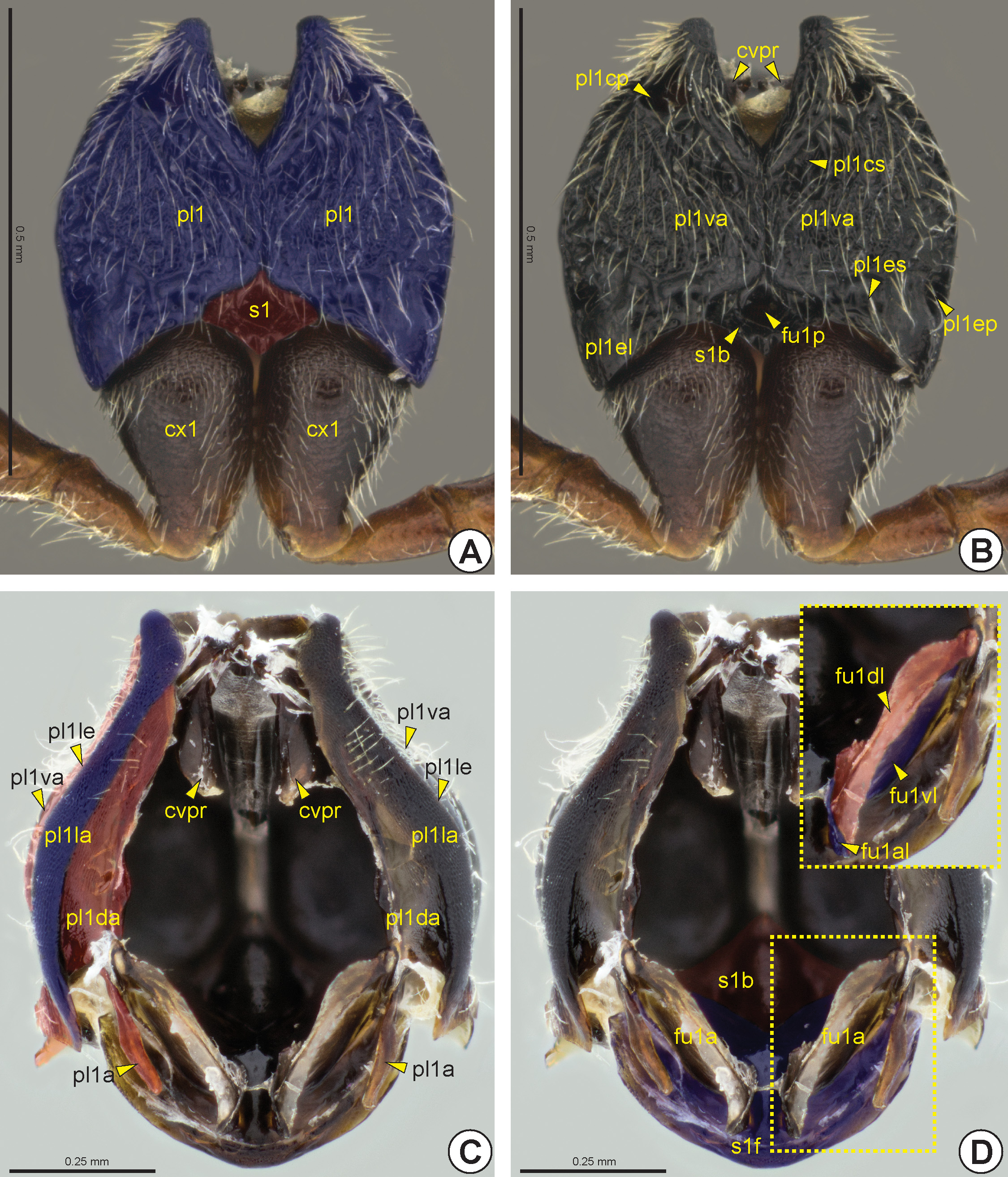
Propleuron: The propleuron (pl1: Figs 12 View FIGURE 12 A–C, 13) is not fused with the prosternum (s1: Figs 12D View FIGURE 12 , 13B View FIGURE 13 ). The propleuron is articulated anteriorly with the head and posteriorly with the pronotum and mesopleuron. In Apocrita,
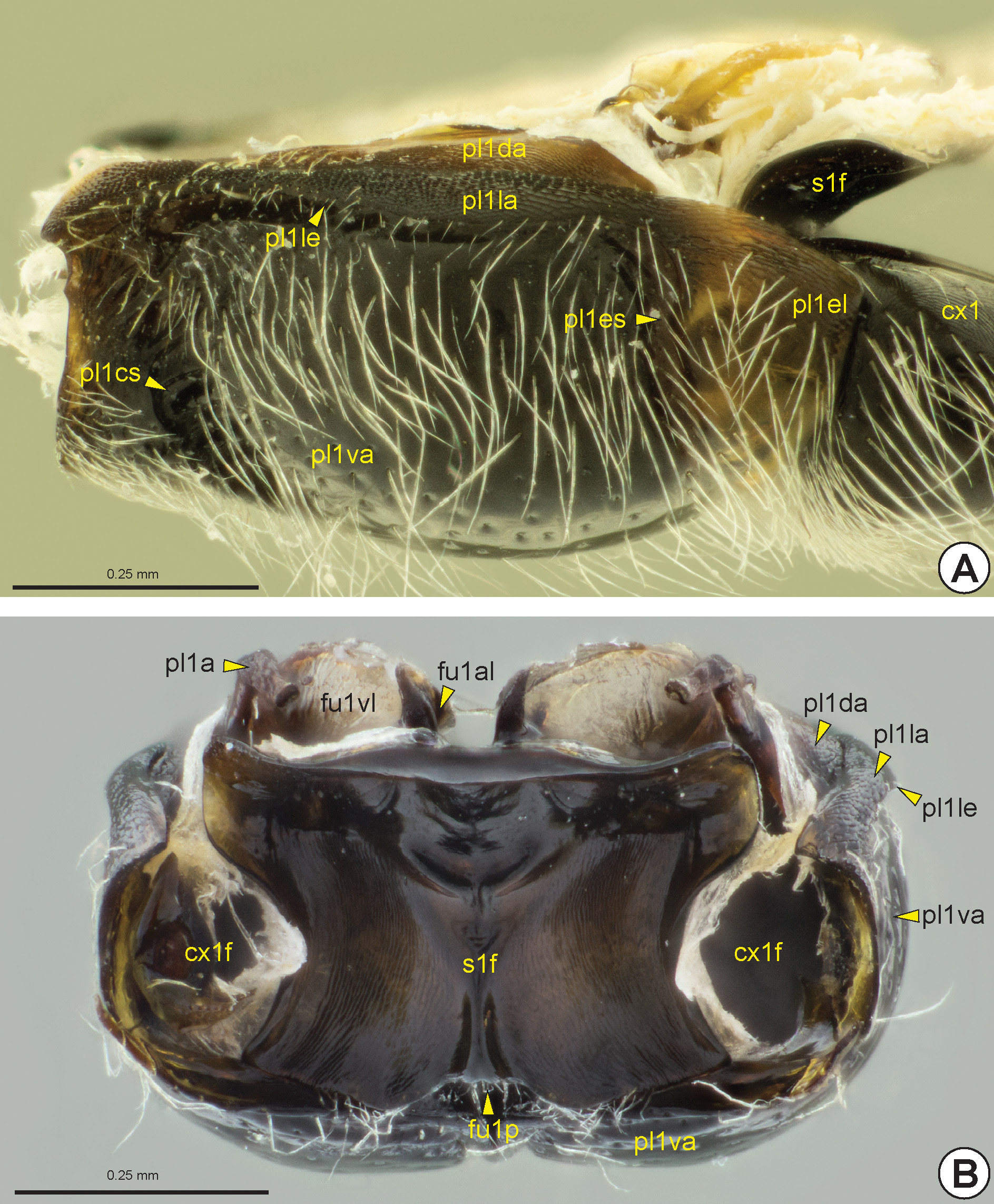
the propleuron is not divided into proepimeron and proepisternum because the propleural sulcus is absent or completely marginal. On the other hand, the propleuron generally is divided into three areas, ventral (pl1va: Figs 12B, 12C View FIGURE 12 , 13 View FIGURE 13 ), lateral (pl1la: Figs 12C View FIGURE 12 , 13 View FIGURE 13 ) and dorsal propleural areas (pl1da: Figs 12C View FIGURE 12 , 13 View FIGURE 13 ). The lateral propleural area is located dorsolaterally on the propleuron and it is concealed by the pronotum and is generally higher and more polished than the ventral propleural area. The boundary between the lateral and ventral propleural areas is defined by the longitudinal propleural edge* (pl1le: Figs 12C View FIGURE 12 , 13 View FIGURE 13 ). In most Bethylidae , the medial margins of the lateral propleural areas are distant from each other ( Figs 12C, 12D View FIGURE 12 ), but in some the anterior corners are very close to each other and in a few the medial margins of the lateral propleural areas are extensively connected; in some of the last group the propleura are anteriorly elongated, forming an externally visible neck between the head and the pronotal flange. The weakly sclerotized dorsal propleural area is located posterodorsally to the lateral propleural area. The propleural arm (pl1a: Figs 12C View FIGURE 12 , 13B View FIGURE 13 ) is usually long and protrudes from the posterior corner of the medial margin of the dorsal propleural area.
The propleural cervical sulcus (pl1cs: Figs 12B View FIGURE 12 , 13A View FIGURE 13 ) extends along the anterior margin of the ventral propleural area. This sulcus is generally deeper at its medial end, forming one or two depressions ( Fig. 12B View FIGURE 12 ). The lateral end of the propleural cervical sulcus includes the propleural cervical pit* (pl1cp: Fig. 12B View FIGURE 12 ), which is internally represented by the cervical prominence (cvpr: Figs 12B, 12C View FIGURE 12 ). The propleural cervical sulcus generally extends through the medial margin of the propleural ventral area and can connect to the propleural epicoxal sulcus (pl1es: Figs 12B View FIGURE 12 , 13A View FIGURE 13 ). The propleural epicoxal sulcus delimits the posterolateral corner of the ventral propleural area, which overlaps the base of the procoxa (cx1: Figs 12A View FIGURE 12 , 13A View FIGURE 13 , 32A View FIGURE 32 ) and forms a distinctive triangular area, the propleural epicoxal lobe* (pl1el: Figs 12B View FIGURE 12 , 13A View FIGURE 13 ). The lateral end of the propleural epicoxal sulcus includes the propleural epicoxal pit* (pl1ep: Fig. 12B View FIGURE 12 ), which internally is represented by the lateral coxal articular process of the propleuron (pl1lcap), which articulates with the basal portion of the procoxa. The procoxal foramen (cx1f: Fig. 13B View FIGURE 13 ) is open dorsally and is formed by the propleuron and prosternum, where the prosternum forms the dorsal and medial margins, while the propleuron forms the ventral and lateral margins.
Prosternum: The prosternum is usually divided into two areas, the external probasisternum (s1b: Figs 12B, 12D View FIGURE 12 ) and the internal profurcasternum (s1f: Figs 12D View FIGURE 12 , 13 View FIGURE 13 ). In most Bethylidae the probasisternum is greatly reduced, being restricted to a diamond-shaped region between the propleura posteriorly, but a few species present a widely visible probasisternum. The prodiscrimen (dc1) is generally represented only by a narrow midline. When present, the profurcal pit (fu1p: Figs 12B View FIGURE 12 , 13B View FIGURE 13 ) is medial and located posteriorly in the probasisternum. Internally, the invagination of the prodiscrimen produces the prodiscriminal lamella (dc1l), which arises anteriorly from the base of the profurca (fu1: Figs 12D View FIGURE 12 , 13B View FIGURE 13 ). The profurcasternum is obliquely vertical and not as heavily sclerotized as the probasistemum. From the internal invagination of the profurcal pit arises the base of the profurca, which extends to the dorsal margin of the profurcasternum, from where the profurcal arms (fu1a: Fig. 12D View FIGURE 12 ) protrude. The profurcal arms are broad and form an oval structure. In most, the dorsal profurcal lamella (fu1dl: Fig. 12D View FIGURE 12 ) is narrow and appears as an expansion of the dorsal surface of the profurcal arm. The ventral profurcal lamella (fu1vl: Figs 12D View FIGURE 12 , 13B View FIGURE 13 ) is larger than the dorsal lamella and comprises the entire ventral surface of the profurcal arm. The anterior profurcal lamella (fu1al: Figs 12D View FIGURE 12 , 13B View FIGURE 13 ) varies in length and is located at the dorsomedial corner of the dorsal profurcal lamella. The proacetabulum (cx1a) accommodates the procoxa and is formed by the external surface of the profurcasternum ( Fig. 13B View FIGURE 13 ).
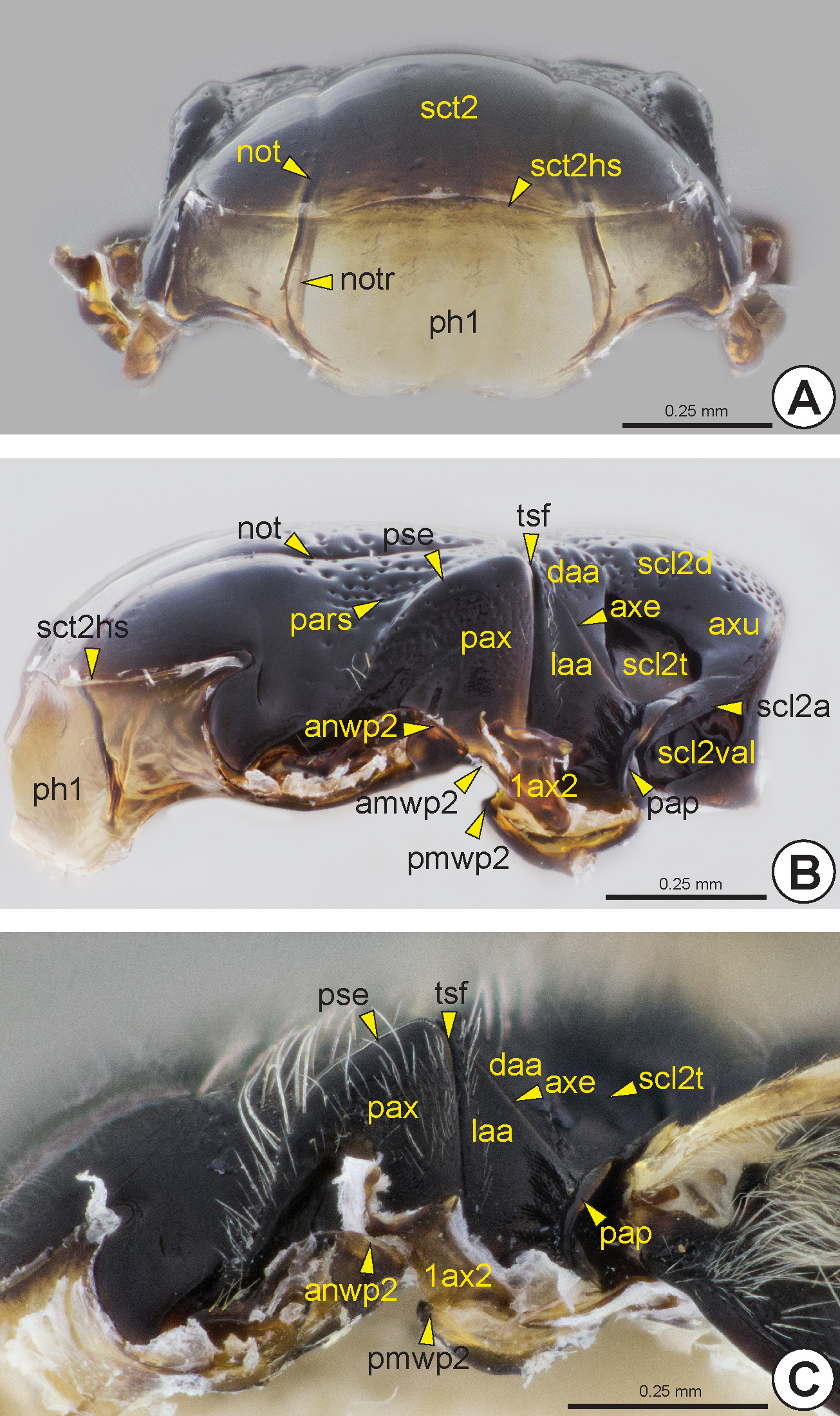
Mesonotum and phragmata: The mesonotum (n2: Figs 14–18 View FIGURE 14 View FIGURE 15 View FIGURE 16 View FIGURE 17 View FIGURE 18 ) is anteriorly articulated with the posterodorsal margin of the pronotum and widely continuous with the internal and transverse prophragma (ph1: Figs 17A, 17B View FIGURE 17 ). The prophragma is large, oriented vertically and not as heavily sclerotized as the mesonotum. The ventral margin of the prophragma can be medially excavated and its dorsal margin is limited by a weak mesoscutal suprahumeral sulcus (sct2hs: Figs 17A, 17B View FIGURE 17 ). The mesonotum laterally adjoins the anterodorsal portion of the mesopleuron and is posterolaterally in contact with the mesopostnotum (pn2).
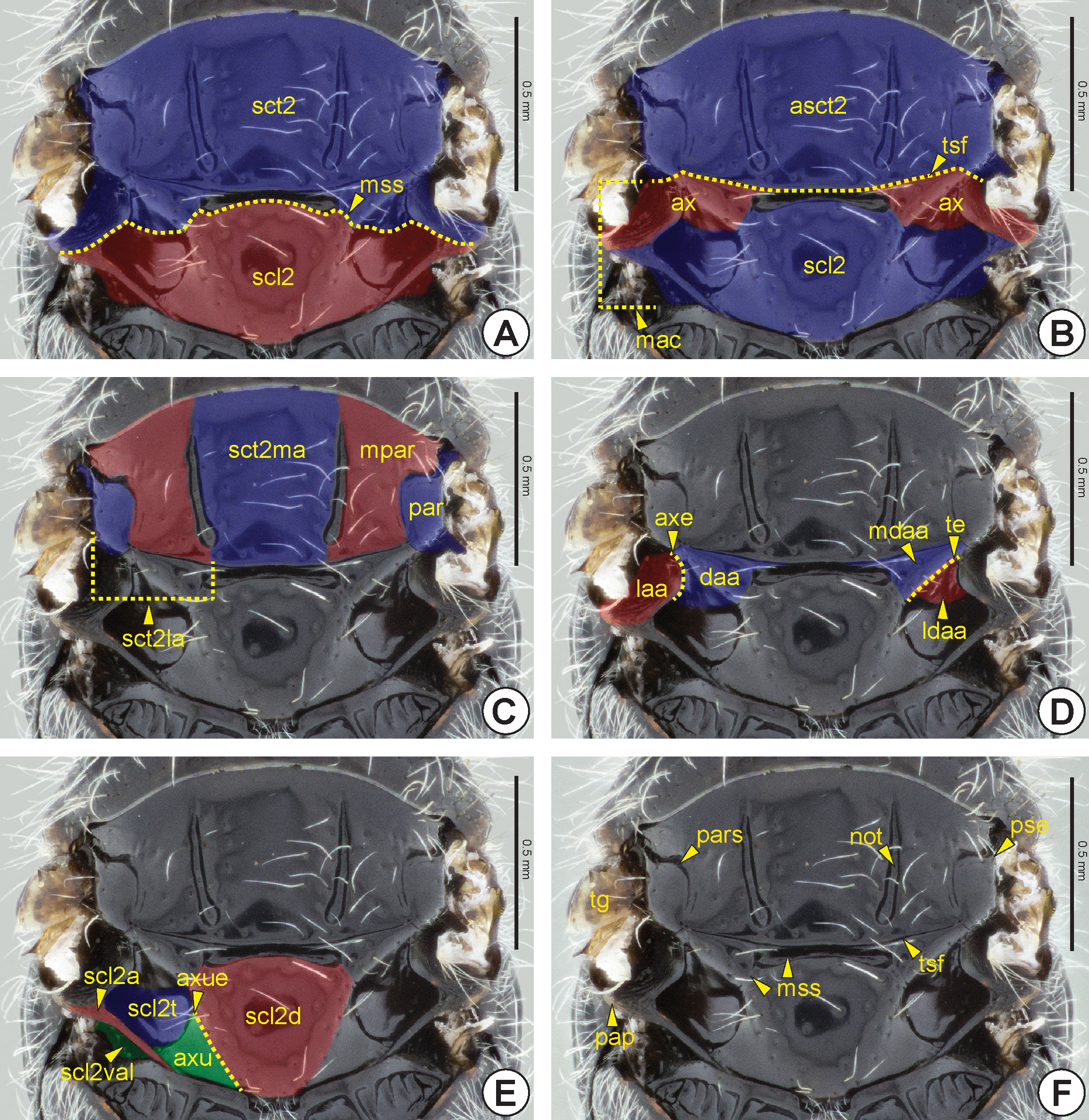
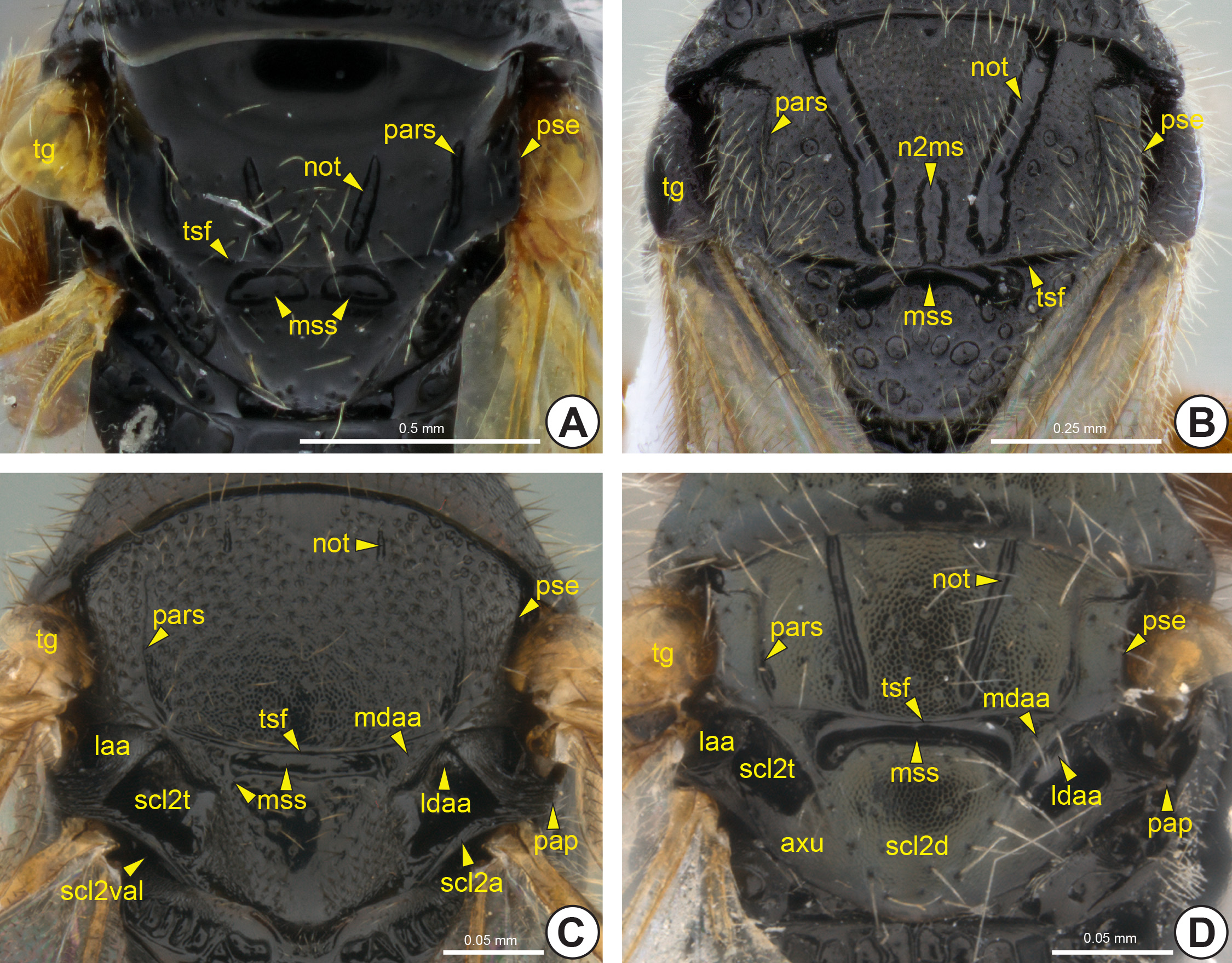
Most Bethylidae have the mesonotum divided into mesoscutum (sct2: Fig. 14A View FIGURE 14 ) and mesoscutellum (scl2: Figs 14A View FIGURE 14 , 16C, 16D View FIGURE 16 , 18A View FIGURE 18 ), by the mesoscuto-scutellar suture (mss: Figs 14A, 14F View FIGURE 14 , 15 View FIGURE 15 , 16 View FIGURE 16 ). Apterous females have the mesoscutum very reduced and often totally covered by the pronotum ( Figs 16C, 16D View FIGURE 16 ). The mesoscutum of Hymenoptera generally has three pairs of linear marks, the anteroadmedian signum (aas), the parapsidal signum (pars: Figs 14F View FIGURE 14 , 15 View FIGURE 15 , 16B View FIGURE 16 , 17B View FIGURE 17 ) and the notaulus (not: Figs 14F View FIGURE 14 , 15 View FIGURE 15 , 16A, 16B View FIGURE 16 , 17A, 17B View FIGURE 17 ). The anteroadmedian signum is a short line that appears submedially from the anterior margin of the mesoscutum and marks the origin of the longitudinal indirect flight muscle; in bethylids it is weakly visible and generally covered by the pronotum. The parapsidal signum, when present, varies in length and is generally represented only by a thin line that marks the origin of the dorsoventral indirect flight muscle. The notaulus appears on the anterior margin of the prophragma and extends along the mesoscutum, between the anteroadmedian and parapsidal signa. In most species of Bethylidae the notaulus is a deep sulcus, but it is rarely represented by a carina. The length, width and direction (parallel or convergent) of the notauli vary between species. Internally, the notaulus is represented by the notaular ridge (notr: Fig. 17A View FIGURE 17 ), which delimits the areas of insertion of the dorsoventral and the longitudinal indirect flight muscles.
The notauli separate the mesoscutum into two areas, the median area of the mesoscutum (sct2ma: Fig. 14C View FIGURE 14 ) and lateral area of the mesoscutum (sct2la: Fig. 14C View FIGURE 14 ). The median area of the mesoscutum is located between the notauli, whereas the lateral area of the mesoscutum is located between the notaulus and the lateral margin of the mesoscutum, the parascutal edge* (pse: Figs 14F View FIGURE 14 , 15 View FIGURE 15 , 16A, 16B View FIGURE 16 , 17B, 17C View FIGURE 17 ). The lateral area is usually subdivided by the parapsidal signum into another two areas, the mediparapsis* (mpar: Fig. 14C View FIGURE 14 ) and parapsis (par: Fig. 14C View FIGURE 14 ). The mediparapsis is located between the notaulus and the parapsidal signum, whereas the parapsis is located between the parapsidal signum and the parascutal edge.
In most Bethylidae , the parascutal edge is smooth and gradually sloping, but in some genera it can be strongly angled, similar to a carina. The surface lateral to the parascutal edge, the preaxilla (pax: Figs 17B, 17C View FIGURE 17 ), is obliquely vertical and accommodates the tegula (tg: Figs 14F View FIGURE 14 , 15 View FIGURE 15 , 16A, 16B View FIGURE 16 ). The lateral margin of the preaxilla is expanded, forming a pair of lobes, the anterior mesonotal wing process (anwp2: Figs 17B, 17C View FIGURE 17 ) and antemedian mesonotal wing process (amwp2: Fig. 17B View FIGURE 17 ). These wing processes articulate with the anterior end and medial portion of the first axillary sclerite of the fore wing (1ax2: Figs 17B, 17C View FIGURE 17 ), respectively.
A few genera of Bethylidae have a longitudinal median mesonotal sulcus* (n2ms: Fig. 15B View FIGURE 15 ), which may be incomplete in a few species, occupying up to the posterior third of the mesoscutum. Internally, this sulcus is represented by the median mesonotal ridge* (n2mr). A few genera have a transverse mesoscutal sulcus* (sct2s: Fig. 16A View FIGURE 16 ), which is deeper and better delimited laterally.
The mesoscutum is separated into two distinct areas, the axilla (ax: Fig. 14B View FIGURE 14 ) and anteromesoscutum (asct2: Fig. 14B View FIGURE 14 ). The axilla is a posterolateral triangular area and is intimately connected to the mesoscutellum, forming the mesoscutellar-axillar complex (mac: Fig. 14B View FIGURE 14 ). The anteromesoscutum comprises most of the mesoscutum and is separated from the mesoscutellar-axillar complex by the transscutal fissure (tsf: Figs 14B, 14F View FIGURE 14 , 15 View FIGURE 15 , 16A, 16B View FIGURE 16 , 17B, 17C View FIGURE 17 ). Internally, the transscutal ridge (tsr) is a weak ridge posteriorly adjacent to the transscutal fissure. The axilla is divided into the dorsal axillar area (daa: Figs 14D View FIGURE 14 , 17B, 17C View FIGURE 17 ) and the lateral axillar area (laa: Figs 14D View FIGURE 14 , 15C, 15D View FIGURE 15 , 17B, 17C View FIGURE 17 ) by the axillar edge* (axe: Figs 14D View FIGURE 14 , 17B, 17C View FIGURE 17 ). In most Bethylidae , the dorsal axillar area is subdivided into the medial surface of the dorsal axillar area* (mdaa: Figs 14D View FIGURE 14 , 15C, 15D View FIGURE 15 ) and lateral surface of the dorsal axillar area* (ldaa: Figs 14D View FIGURE 14 , 15C, 15D View FIGURE 15 ) by a weak transaxillar edge* (te: Fig. 14D View FIGURE 14 ). The lateral margin of the lateral axillar area is expanded, forming a pair of lobes, the postmedian mesonotal wing process (pmwp2: Figs 17B, 17C View FIGURE 17 ) and the postalar process (pap: Figs 14F View FIGURE 14 , 15C, 15D View FIGURE 15 , 17B, 17C View FIGURE 17 ). The postmedian mesonotal wing process is located immediately posterior to the transscutal fissure and articulates with the posterior end of the first axillary sclerite of the fore wing. The postalar process consists of a strong posterodorsal projection of the axilla and separates the site of origin of the fore wing (fwg: Figs 27–30 View FIGURE 27 View FIGURE 28 View FIGURE 29 View FIGURE 30 ) from the site of origin of the hind wing (hwg: Fig. 31 View FIGURE 31 ).
In most Bethylidae , the mesoscuto-scutellar suture (mss) is medially sulcate ( Figs 14 View FIGURE 14 , 15 View FIGURE 15 B–D, 16A, 16B).
Generally, the lateral extremities of this sulcus are dilated ( Fig. 15A View FIGURE 15 ), which are commonly described as foveae. In some genera, these dilations are strongly distinctive because of the weakness or absence of the sulcus between them. In most Bethylidae the mesoscuto-scutellar suture is laterally inconspicuous, but when present it is represented by a thin sulcus or line. The mesoscuto-scutellar suture is represented internally by the mesoscuto-scutellar ridge* (mssr: Fig. 18B View FIGURE 18 ), which has a shape and thickness corresponding to its external condition; in species with the mesoscuto-scutellar suture represented only by lateral dilations, the mesoscuto-scutellar ridge is moderately narrowed. The mesoscuto-scutellar ridge is projected posteriorly, forming a narrow plate that covers the anterior region of the scutellum, and is continuous posterolaterally with the mesoscutellar bridge * (scl2b: Fig. 18B View FIGURE 18 ), which merges with the posterior part of the mesoscutellum.
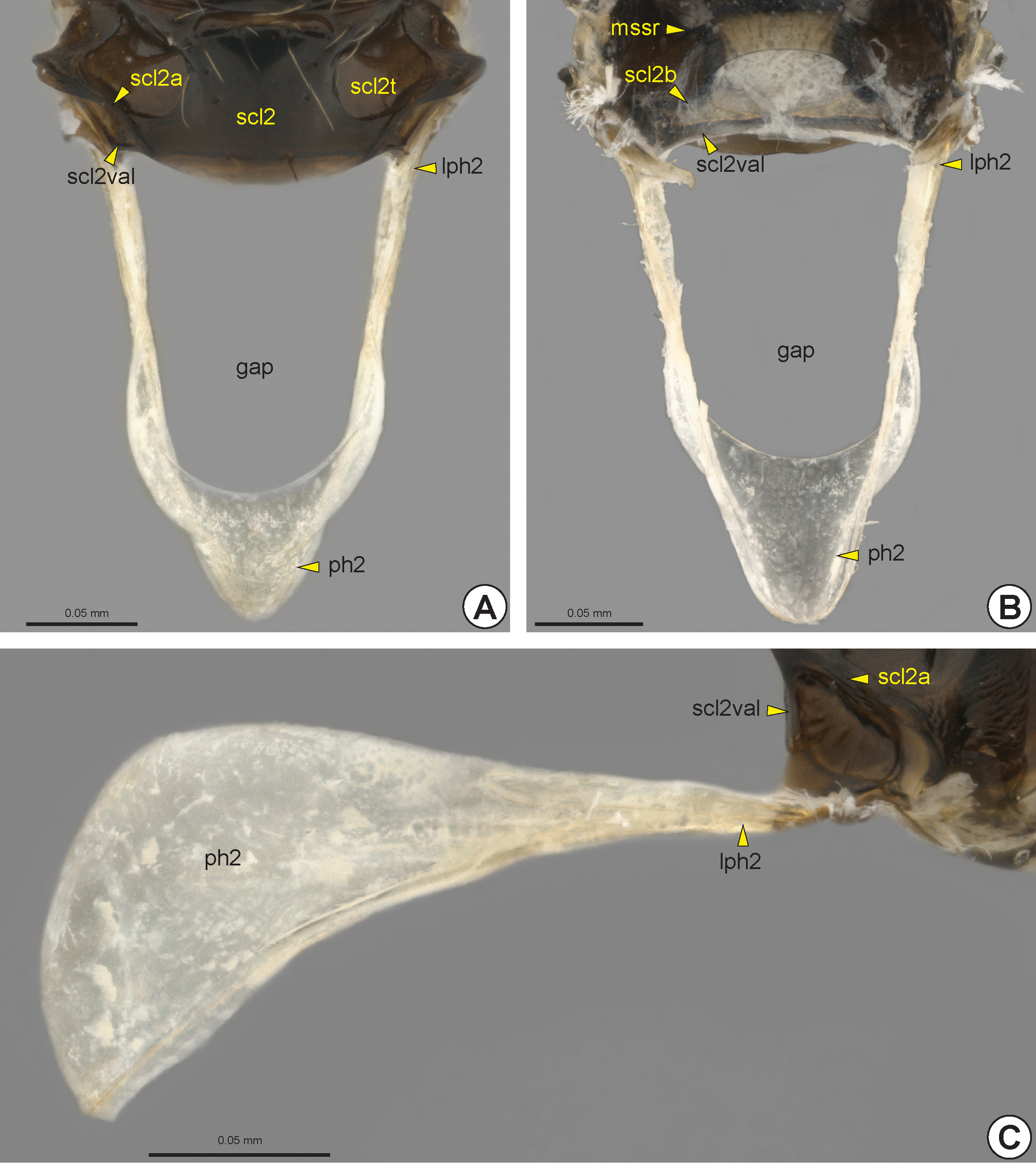
The median area of the mesoscutellum is usually separated into two areas, the mesoscutellar disc (scl2d: Figs 14E View FIGURE 14 , 15D View FIGURE 15 , 16A, 16B View FIGURE 16 , 17B View FIGURE 17 ) and axillula (axu: Figs 14E View FIGURE 14 , 15D View FIGURE 15 , 16A View FIGURE 16 , 17B View FIGURE 17 ), which are separated by the axillular edge* (axue: Fig. 14E View FIGURE 14 ). The axillula is a triangular area located lateral to the mesoscutellar disc and is delimited posterolaterally by the mesoscutellar arm (scl2a: Figs 14E View FIGURE 14 , 15C View FIGURE 15 , 16A, 16B View FIGURE 16 , 17B View FIGURE 17 , 18A, 18C View FIGURE 18 ) and anterolaterally by the mesoscutellar trough (scl 2t: Figs 14E View FIGURE 14 , 15C, 15D View FIGURE 15 , 16A, 16B View FIGURE 16 , 17B, 17C View FIGURE 17 , 18A View FIGURE 18 ). The mesoscutellar trough is usually depressed, forming a large fovea. The mesoscutellar arm extends laterally along the posterior margin of the mesoscutellum. The vertical apodemal lobe of the mesoscutellum (scl2val: Figs 14E View FIGURE 14 , 15C View FIGURE 15 , 17B View FIGURE 17 , 18 View FIGURE 18 ) is the internal inflection that extends along the posterolateral margin of the mesoscutellum and is laterally continuous with the mesoscutellar bridge.
Internally, the mesopostnotum is composed of the lateral and well sclerotized mesolaterophragma (lph2: Fig. 18 View FIGURE 18 ) and the posterior and weakly sclerotized mesophragma (ph2: Fig. 18 View FIGURE 18 ); the latter serves as a surface for insertion of the indirect longitudinal flight muscle. The mesopostnotum protrudes posteriorly, forming a large median gap ( Figs 18A, 18B View FIGURE 18 ); it extends to the end of the metapectal-propodeal complex and occupies the area externally represented by the metapostnotum (pn3: Fig. 23A View FIGURE 23 ).
Mesopectus: The mesopleurosternal suture is absent and there is, therefore, no clear delimitation between mesopleuron and mesosternum. Thus, the mesopectus (pec2) is the complex formed by the mesopleuron (pl2: Fig. 19 View FIGURE 19 ), mesosternum (s2: Fig. 20 View FIGURE 20 ), mesofurca (fu2: Fig. 21 View FIGURE 21 ) and some intersegmental sclerites [mesobasalare (ba2) and mesosubalare (sa2)]. The mesopectus is anteriorly articulated with the pronotum and propleuron, and posteriorly with the metapectal-propodeal complex.
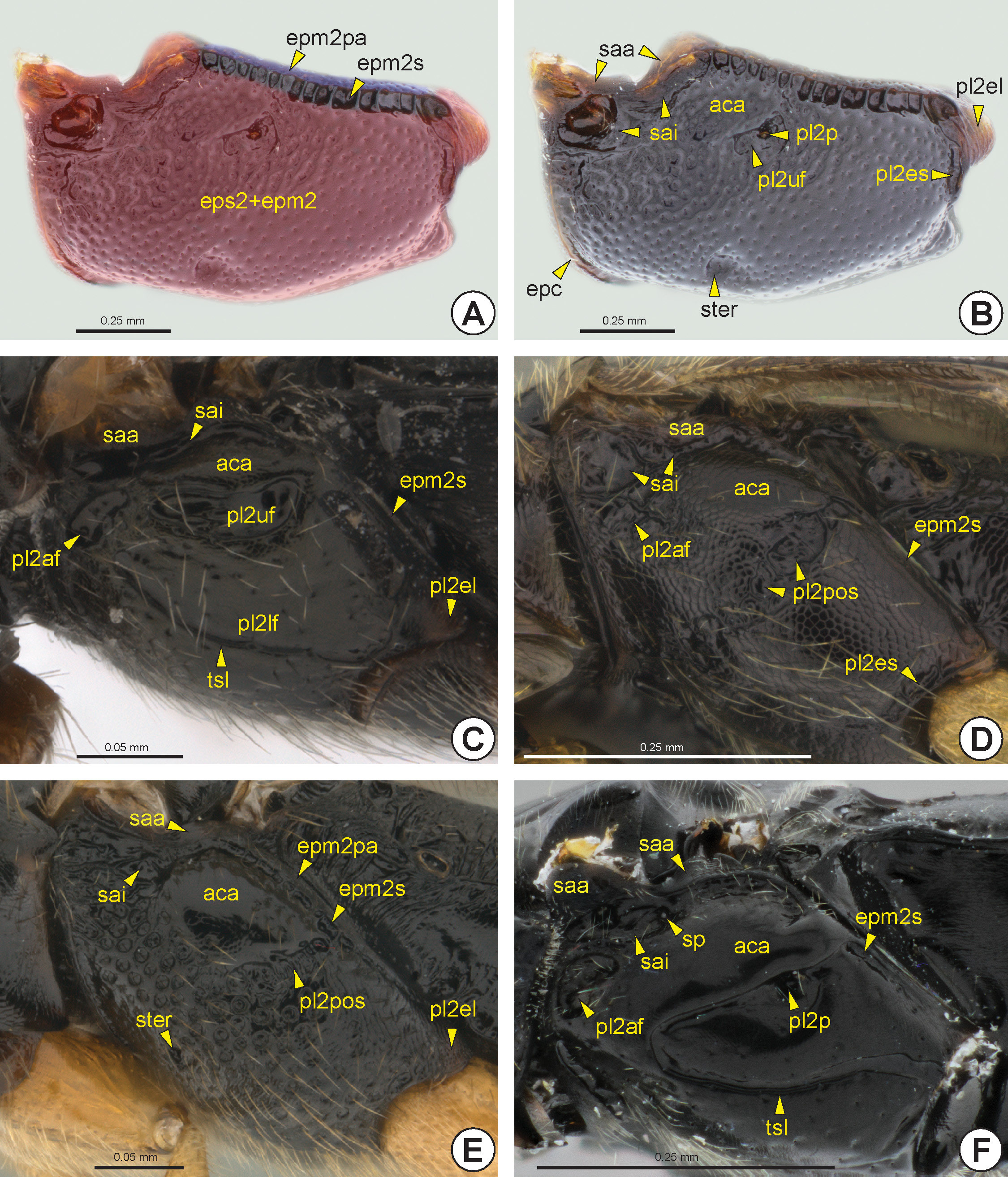
The mesopleuron of Bethylidae is not clearly divided into mesepimeron (epm2) and mesepisternum (eps2) ( Fig. 19A View FIGURE 19 ), because the mesopleural suture (pl2s) is obliterated. Dorsally, the mesopleuron of the most Bethylidae has the mesepimeral sulcus (epm2s: Figs 19A, 19 View FIGURE 19 C–F), which separates the very narrow posterior mesepimeral area (epm2pa: Figs 19A, 19E View FIGURE 19 ) from the other areas of the mesopleuron. The mesepimeral sulcus can be anteriorly
connected to the subalar impression (sai: Figs 19 View FIGURE 19 B–F). Internally, the mesepimeral sulcus is represented by the mesepimeral ridge (epm2r: Fig. 21B, 21C View FIGURE 21 ). In some species the subalar impression is anteriorly deeper, which is represented internally by the acropleural apodeme (acra). In some genera the anterodorsal region of the mesopleuron can be bulging and polished, the acropleural area* (aca: Figs 19 View FIGURE 19 B–F). Dorsal to the subalar impression is the subalar area (saa: Figs 19 View FIGURE 19 B–F), which bears the mesopleural wing process* (pl2wp). This process includes the anterodorsal incision of the mesopleuron (pl2ai), where the mesobasalare fits. Posteriorly, the mesopleural wing process connects to the second axillary sclerite of the fore wing (2ax2).
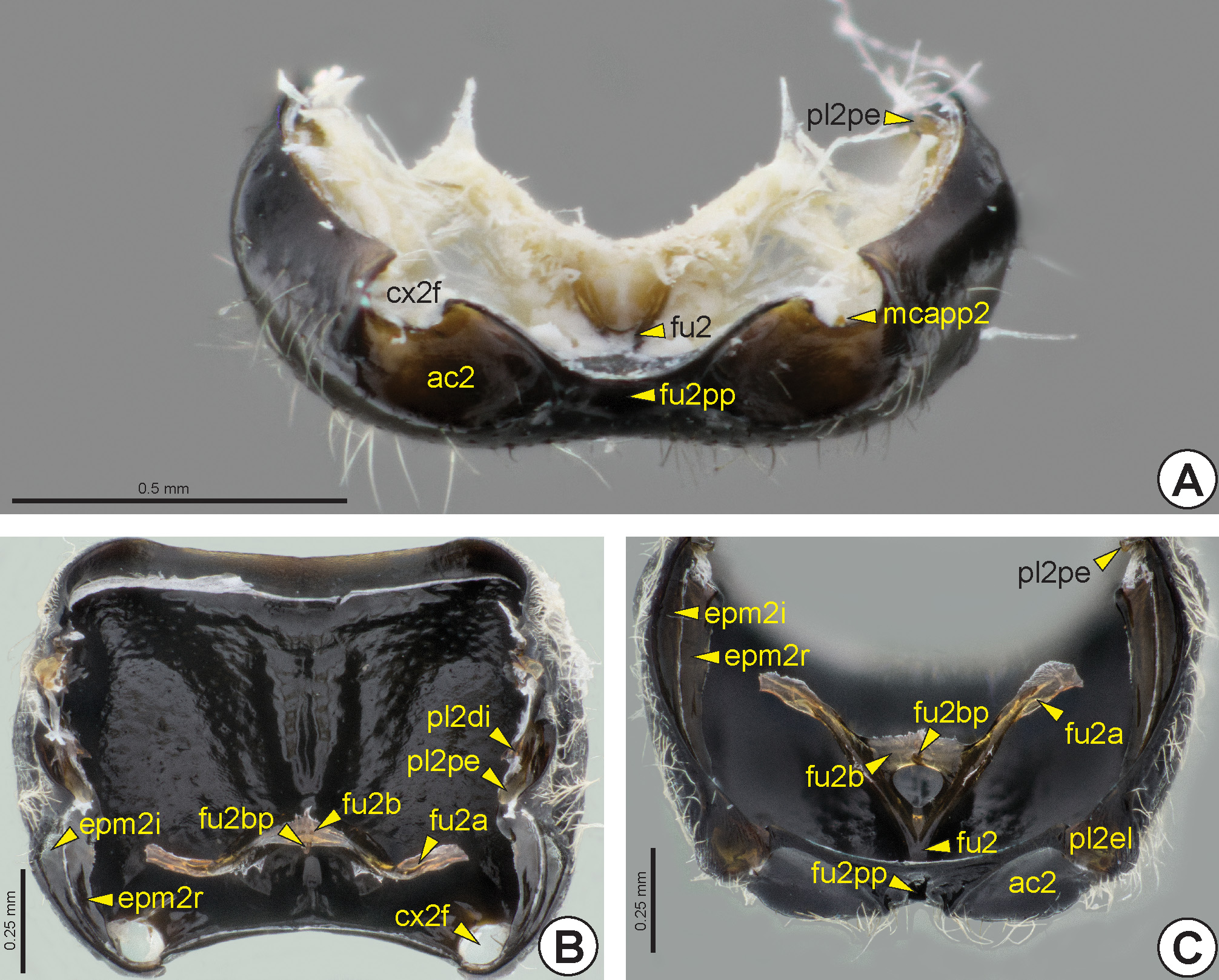
The posterior end of the mesepimeral sulcus is usually continuous with the mesopleural epicoxal sulcus (pl2es: Figs 19B, 19D View FIGURE 19 ). This is located anterior to the mesocoxa (cx2: Figs 32B View FIGURE 32 ), and separates the mesopleural epicoxal lobe* (pl2el: Figs 19B, 19C, 19E View FIGURE 19 , 21C View FIGURE 21 ) from the rest of the mesopleuron. The mesopleural epicoxal sulcus is represented internally by the mesopleural epicoxal ridge * (pl2er). Internally, at the posterior end of the mesepimeral ridge, along with the mesopleural epicoxal ridge, is the lateral coxal articular process of the mesopleuron (lcapp2). The medial coxal articular process of the mesopleuron * (mcapp2: Figs 20B View FIGURE 20 , 21A View FIGURE 21 ) is well developed and located on the medial margin of the mesocoxal foramen (cx2f: Figs 20 View FIGURE 20 A–C, 21A, 21B).
The mesopleural pit (pl2p: Figs 19B, 19F View FIGURE 19 ) is located centrally in the mesopleuron and is represented internally by the mesopleural apodeme (pl2a). Some species have a posterior oblique sulcus of the mesopleuron (pl2pos: Figs 19D, 19E View FIGURE 19 , 20D View FIGURE 20 ), which extends anteroventrally from the mesopleural pit to the anterior margin of the ventral mesopleuron. This sulcus is internally represented by the posterior oblique ridge of the mesopleuron * (pl2por), which is continuous with the mesopleural apodeme. In many Bethylidae there is a small fovea located anteroventrally on the mesopleuron, the sternaulus (ster: Figs 19B, 19E View FIGURE 19 ). When the posterior oblique sulcus is complete, this fovea is located at the anteroventral end of this sulcus.
In several genera the mesopleural pit appears within the upper mesopleural fovea (pl2uf: Figs 19B, 19C View FIGURE 19 ). Most Bethylidae have an anterior mesopleural fovea (pl2af: Figs 19C, 19D, 19F View FIGURE 19 ) represented by a small depression located near the anterior margin of the mesopleuron and below the anterior end of the subalar impression; it is usually irregular and open posteriorly. The lower mesopleural fovea (pl2lf: Fig. 19C View FIGURE 19 ) is a broad depression below the mesopleural pit in some species. This depression can be completely enclosed, divided into two foveae, fused with the upper mesopleural fovea or restricted to its ventral margin, the last condition denoted as the transepisternal line (tsl: Figs 19C, 19F View FIGURE 19 ). There is rarely a mesopleural dentate process* (pl2dp: see Figure 11 View FIGURE 11 in Alencar and Azevedo, 2011), a sharp elevation located in the median region of the mesopleuron posterior to the mesopleural pit.
Ventrally on the mesopectus, the subpleural signum (sps: Figs 20B, 20D View FIGURE 20 ) is a short longitudinal and weak line near the posterior margin, anterior to the mesacetabulum (ac2: Figs 20 View FIGURE 20 A–C, 21A, 21C). Internally, the subpleural signum is defined by a weak mark or linear elevation at the origin of the dorsoventral indirect flight muscle.
The prepectus (pre: Fig. 20 View FIGURE 20 ) is an independent and broad intersegmental sclerite that occupies the full extent of the anterior margin of the mesopectus. In most Bethylidae the prepectus is formed by a pair of narrow ventral plates. The anterior and posterior margins of the prepectus each have a flange, the anterior prepectal flange* (preaf: Figs 20 View FIGURE 20 A–C) and the posterior prepectal flange * (prepf: Fig. 20A View FIGURE 20 ). In some genera, the prepectus is formed by a single large plate with a narrow anterior flange.
The epicnemium (epc: Figs 19B View FIGURE 19 , 20 View FIGURE 20 ), when present, is represented by a narrow strip, often similar to a carina. This structure extends along the anterior margin of the mesopectus and generally precedes the postepicnemial sulcus* (peps: Figs 20A, 20B View FIGURE 20 ). When the prepectus is divided into two plates, the epicnemium usually has an anterior epicnemial medial projection* (epcmp: Fig. 20D View FIGURE 20 ).
The mesodiscrimen (dc2: Fig. 20 View FIGURE 20 ) can be represented by a line or a longitudinal sulcus on the mesosternum (s2). The mesodiscrimen marks the location of the invagination of the mesosternum and corresponds internally to the mesodiscrimenal lamella (dc2l) which arises from the mesofurcal base. The mesofurcal arm (fu2a: Figs 21B, 21C View FIGURE 21 ) is connected laterally to the posterodorsal edge of the mesopleuron (pl2pe: Fig. 21 View FIGURE 21 ), which is rarely represented by an evident subalar pit (sp: Fig. 19F View FIGURE 19 ). Medially the mesofurcal arms are connected by the mesofurcal bridge (fu2b: Figs 21B, 21C View FIGURE 21 ), which carries the anterior process of the mesofurcal bridge (fu2bp: Figs 21B, 21C View FIGURE 21 ). The posterodorsal edge of the mesopleuron arises anterodorsally between the dorsal mesopleural inflection (pl2di: Fig. 21B View FIGURE 21 ) and the mesepimeral inflection (epm2i: Figs 21B, 21C View FIGURE 21 ). Externally, the mesofurcal bases are represented by the anterior mesofurcal pit* (fu2ap: Figs 20A, 20C, 20D View FIGURE 20 ) and the posterior mesofurcal pit* (fu2pp: Figs 20 View FIGURE 20 A–C, 21A, 21C).
In Bethylidae the mesacetabulum is generally bordered by the mesacetabular carina* (ac2c). The mesocoxal foramen is located along the lateral margin of the mesacetabulum and is ventrally open in many genera ( Fig. 21A View FIGURE 21 ). The mesacetabulum is continuous with the metepisternal depression (mtd: Figs 25 View FIGURE 25 , 26A, 26C, 26D View FIGURE 26 ) to accommodate the mesocoxa.
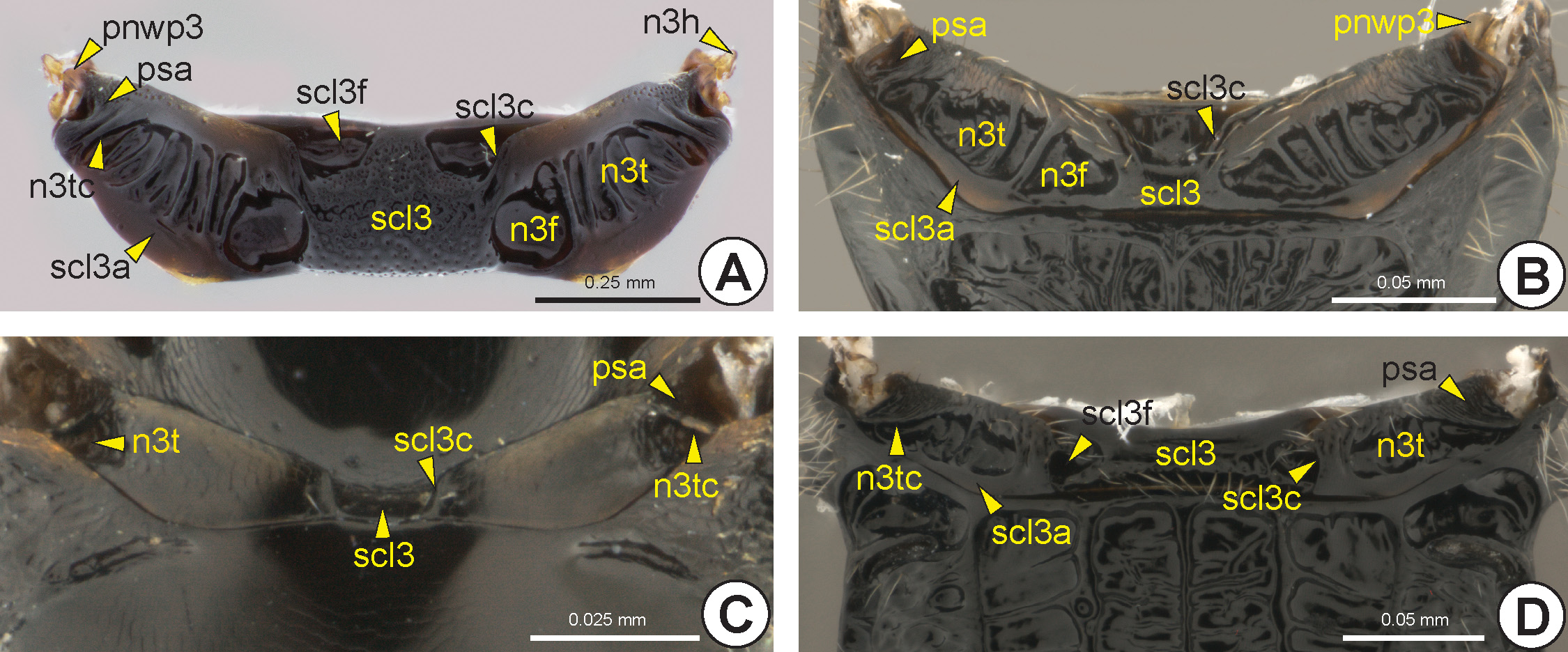
Metanotum: In most Bethylidae the metanotum is distinct, although in some species the metanotum is absent or fused to the metapectal-propodeal complex. The metascutellum (scl3: Fig. 22 View FIGURE 22 ) is usually delimited laterally by the metascutellar carina (scl3c: Fig. 22 View FIGURE 22 ). In most Bethylidae the metascutellar carina originates from the posterior margin of the metanotum ( Figs 22C, 22D View FIGURE 22 ), although in a few it emerges from the dorsal surface of the metanotum ( Figs 22A, 22B View FIGURE 22 ). The metascutellum sometimes has one pair of foveae, the metascutellar fovea* (scl3f: Figs 22A, 22D View FIGURE 22 ). Internally, the metascutellum is represented by the chamber of the metanotum (n3c), which is represented only by a thin transverse depression. Lateral to the metascutellum, the metanotal trough (n 3t: Fig. 22 View FIGURE 22 ) generally forms a foveolate depression, internally represented by the weakly differentiated metanotal ridge (n3r). The lateral margin of the metanotal trough may be delimited by an acute elevation, the transmetanotal carina (n3tc: Figs 22A, 22C, 22D View FIGURE 22 ). Lateral to the transmetanotal carina or to the external margin of the metanotal trough is the posterior supraalar area (psa: Fig. 22 View FIGURE 22 ), which can be flat or depressed. In some there is a metanotal fovea (n3f: Figs 22A, 22B View FIGURE 22 ) between the metanotal trough and the metascutellum. The metascutellar arm (scl3a: Figs 22A, 22B, 22D View FIGURE 22 ) is a narrow and elevated area that extends along the posterior margin of the metanotum, posterior to the metanotal trough. The anterior metanotal wing process (anwp3) is located at the apical end of the humeral sclerite of the metanotum (n3h: Fig. 22A View FIGURE 22 ), which is elongated and transversally connected to the ventral surface of the metanotum. The postmedian metanotal wing process (pnwp3: Figs 22A, 22B View FIGURE 22 ) is located anteroventrally on the posterior supraalar area.
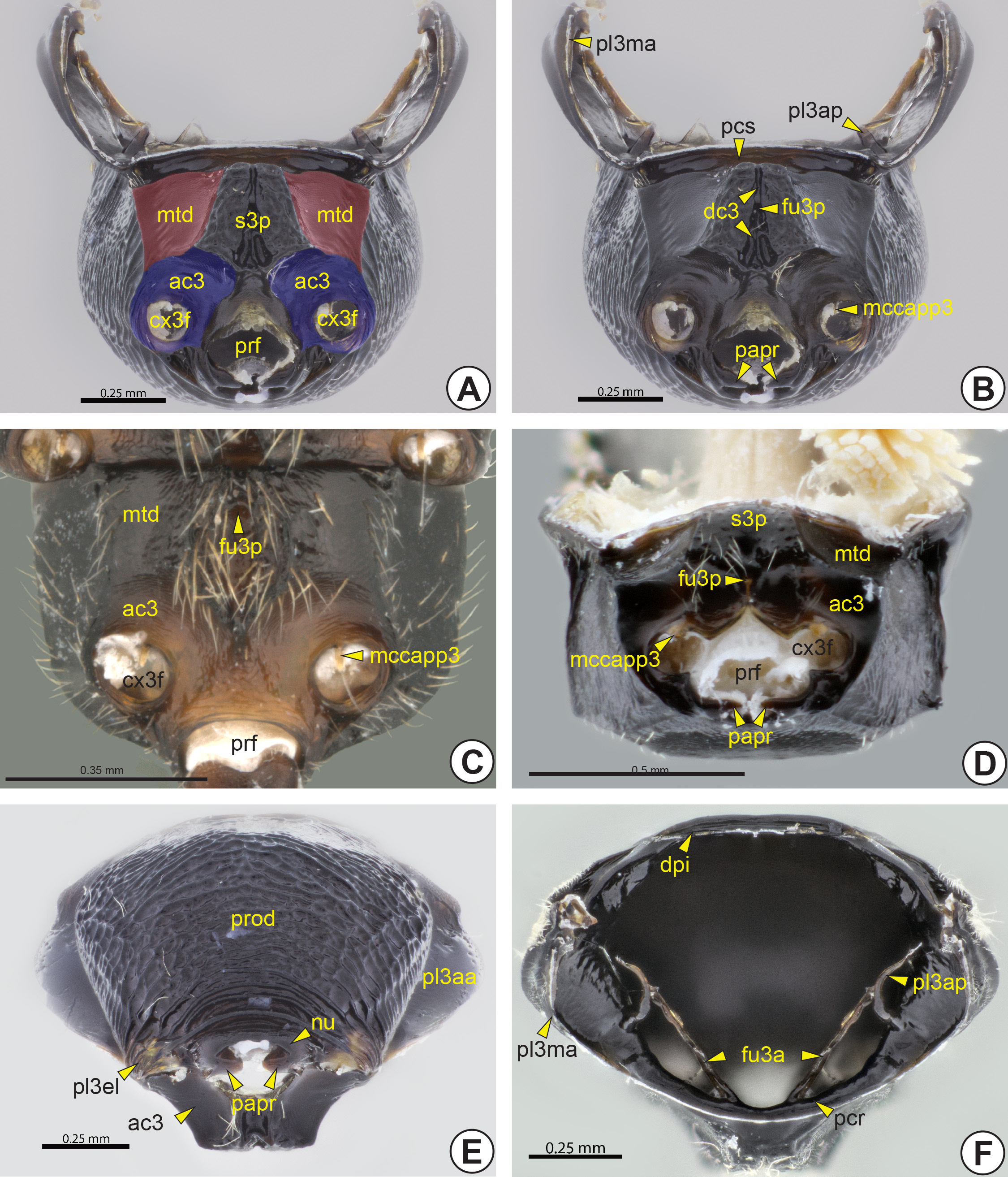
Metapectal-propodeal complex: The complex formed by the metapectus (pec3), the metapostnotum and the propodeum (pro: Figs 23A View FIGURE 23 , 33C View FIGURE 33 ) (= first abdominal tergum). The metapectus is formed by the metapleuron (pl3), metasternum (s3), metafurca (fu3: Fig. 26F View FIGURE 26 ) and some pleural sclerites [metabasalare (ba3) and metasubalare (sa3)]. As in the mesothorax, there is no distinction between the pleuron and sternum in the metathorax (th3), because the metapleurosternal suture is obliterated.
Dorsally, the anterolateral corners of the metapectal-propodeal complex extend anteriorly to form the prespiracular propodeal area (ppa: Figs 23B, 23F View FIGURE 23 , 24 View FIGURE 24 C–G), which is located anterior to the propodeal spiracle (psp: Figs 23B, 23 View FIGURE 23 D–F, 24A–F, 25A, 25B). In many bethylids, the prespiracular propodeal area may be totally flat or present a deep depression, the prespiracular propodeal depression* (ppd: Figs 23B, 23 View FIGURE 23 D–F, 24A, 24B, 25C). The metapleural arm (pl3a: Fig. 25B View FIGURE 25 ) is the anterodorsal extension of the metapleuron and is located anterolateral to the prespiracular propodeal area. The metapleural arm bears the metapleural wing process (pl3wp: Fig. 25B View FIGURE 25 ), to which the metabasalare is connected.
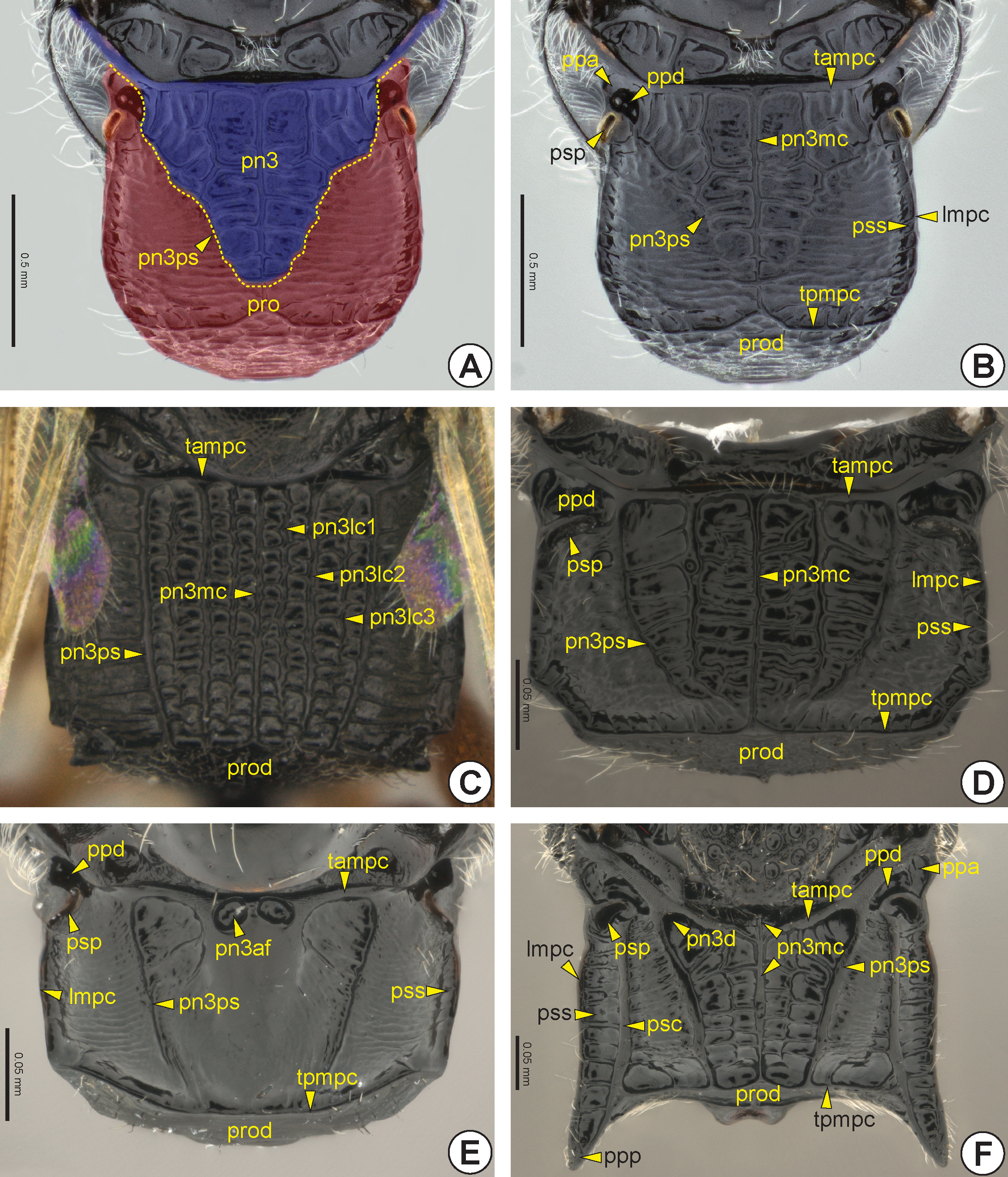
The dorsal surface of the metapectal-propodeal complex comprises the metapostnotum, the median area delimited by the metapostnotal-propodeal suture* (pn3ps: Figs 23 View FIGURE 23 , 24A, 24B, 24D View FIGURE 24 ), and part of the propodeum. The metapostnotal-propodeal suture is often represented by a carina ( Figs 23 View FIGURE 23 B–D, 23F, 24A, 24B), but in many species it is obliterated and the limits of the metapostnotum can be differentiated from the propodeum by the sculpturing, flatness ( Figs 23E View FIGURE 23 , 24D View FIGURE 24 ) and/or pigmentation of the integument. The propodeal spiracle can be located on the dorsal or lateral surface of the propodeum. When dorsal, this spiracle usually is sickle-shaped, and when lateral, it is generally circular. In several, the posterolateral corner of the metapectal-propodeal complex is posteriorly projected, forming the posterior propodeal projection (ppp: Figs 23F View FIGURE 23 , 25C View FIGURE 25 ), but in others the posterolateral corner may be concave ( Figs 23C, 23D View FIGURE 23 , 25D View FIGURE 25 ). Internally, the metapectal-propodeal complex shows the dorsal propodeal inflection (dpi: Fig. 26F View FIGURE 26 ), an inflection that extends along its anterior margin and accommodates the metanotum. The metaphragma (ph3) is poorly delimited, but, judging from the muscular insertions, it seems to be located along the margin of the metapostnotum.
The dorsal surface of the metapectal-propodeal complex is usually limited laterally by the lateral marginal carina of the metapectal-propodeal complex* (lmpc: Figs 23B, 23 View FIGURE 23 D–F, 24B–G, 25A, 25C, 25D), anteriorly by the transverse anterior carina of the metapectal-propodeal complex* (tampc: Figs 23 View FIGURE 23 , 24 View FIGURE 24 A–C, 24E, 25B) and posteriorly by the transverse posterior carina of the metapectal-propodeal complex* (tpmpc: Figs 23B, 23 View FIGURE 23 D–F, 24B, 25A). The transverse anterior carina may sometimes be displaced slightly posteriorly ( Fig. 23F View FIGURE 23 ). From the transverse anterior carina there may arise the metapostnotal median carina (pn3mc; Figures 23 View FIGURE 23 B–D, 23F, 24A, 24D, 24E) that can be posteriorly incomplete or complete, the latter when it reaches the transverse posterior carina. In some species the propodeal declivity (prod: Figs 23 View FIGURE 23 B–F, 24A–C, 24E–G, 25A, 26E) have an extension of the metapostnotal median carina. The lateral marginal carina of the metapectal-propodeal complex extends from the prespiracular propodeal area to the metacoxal articulation. Along the medial margin of the lateral marginal carina of the metapectal-propodeal complex of some species, there is the paraspiracular sulcus (pss: Figs 23B, 23 View FIGURE 23 D–F, 24B–G, 25C) extending longitudinally from the propodeal spiracle to the transverse posterior carina. Usually, a paraspiracular carina (psc: Figs 23F View FIGURE 23 , 24E View FIGURE 24 , 25C View FIGURE 25 ) extends along the medial margin of the paraspiracular sulcus. Bethylidae have up to three carinae between the metapostnotal median carina and metapostnotal-propodeal suture, the first metapostnotal lateral carina* (pn3lc1: Fig. 23C View FIGURE 23 ), second metapostnotal lateral carina* (pn3lc2: Fig. 23C View FIGURE 23 ), and third metapostnotal lateral carina* (pn3lc3: Fig. 23C View FIGURE 23 ). The metapostnotum shows much variation in sculpture, including a pair of anteromedial metapostnotal foveae* (pn3af: Figs 23E View FIGURE 23 , 24B View FIGURE 24 ), or a metapostnotal depression (pn3d: Figs 23F View FIGURE 23 , 24A View FIGURE 24 ). The metapostnotal depression is located in the anterolateral corner of the metapostnotum and communicates with the prespiracular propodeal depression. In some species, the prespiracular propodeal depression communicates with the dorsal metapleural pit* (pl3dp: Fig. 25C View FIGURE 25 ) of the lateral surface of the metapectal-propodeal complex.
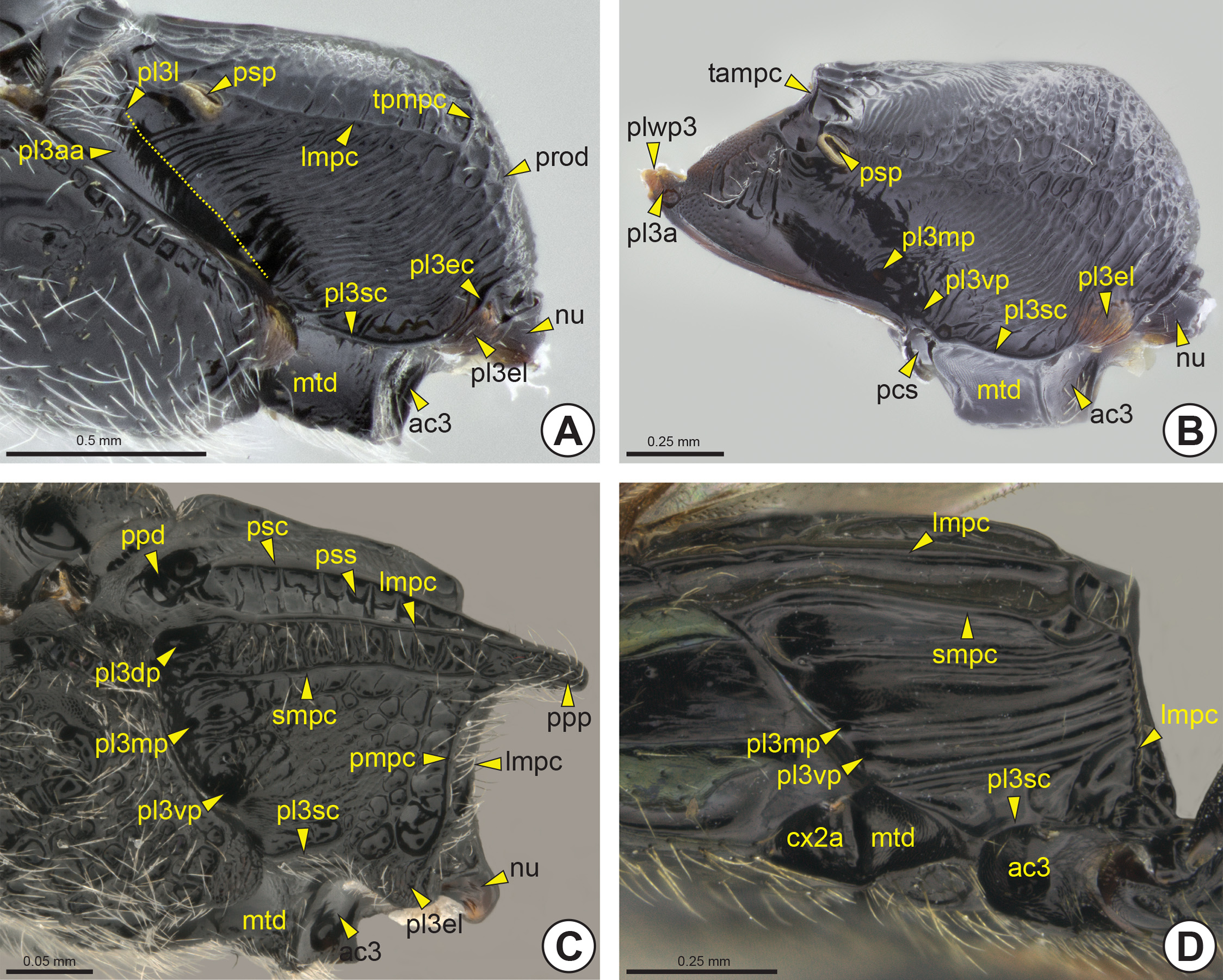
The metapleural-propodeal suture is totally obliterated and therefore the exact boundary between the propodeum and the metapectus is not evident. The anterior margin of the lateral surface of the metapectal-propodeal complex is inflected, forming the marginal metapleural apodeme (pl3ma: Fig. 26F View FIGURE 26 ) internally. In most Bethylidae , the lateral surface of the metapectal-propodeal complex has an anterior triangular area, the anterior metapleural area (pl3aa: Figs 25A View FIGURE 25 , 26E View FIGURE 26 ), which is posteriorly delimited by the metapleural line (pl3l: Fig. 25A View FIGURE 25 ) which is usually represented only by a difference in texture and profile. Along the metapleural line there are the dorsal metapleural pit, median metapleural pit* (pl3mp: Figs 25 View FIGURE 25 B–D) and ventral metapleural pit* (pl3vp: Figs 25 View FIGURE 25 B–D). In some Bethylidae all three metapleural pits are present, but most genera have only the median metapleural pit. Internally,
the median metapleural pit is represented by the thin and sinuous metapleural apodeme (pl3ap: Figs 26B, 26F View FIGURE 26 ). In other families this apodeme occurs on the metapleural ridge, but in Bethylidae this structure is absent. In some, the lateral surface of the metapectal-propodeal complex has two additional carinae, the submarginal carina of the metapectal-propodeal complex* (smpc: Figs 25C, 25D View FIGURE 25 ) and posterior carina of the metapectal-propodeal complex * (pmpc: Fig. 25C View FIGURE 25 ).
The submetapleural carina (pl3sc: Fig. 25 View FIGURE 25 ) is located along the ventral margin of the lateral surface of the metapectal-propodeal complex, between the mesocoxal and the metacoxal foramina (cx3f: Figs 26A, 26 View FIGURE 26 C–D). The posteroventral corner of the lateral surface may include a differentiated triangular area which involves the articulation of the metacoxa and forms the metapleural epicoxal lobe* (pl3el: Figs 25 View FIGURE 25 A–C, 26E). The distinction between the metapleural epicoxal lobe and the rest of the metapectal-propodeal complex is usually inconspicuous, but in some species there is the metapleural epicoxal carina (pl3ec: Fig. 25A View FIGURE 25 ) between these areas; it is anteriorly continuous with the submetapleural carina. Anteriorly along the metapleural epicoxal carina a shallow sulcus may extend, the metapleural epicoxal sulcus (pl3es). The metacetabulum (ac3: Figs 25 View FIGURE 25 , 26A, 26C, 26D, 26E View FIGURE 26 ) is often flanked by the metacetabular carina* (ac3c). The medial coxal articular process of the metapleuron* (mccapp3: Figs 26B, 26C, 26D View FIGURE 26 ) is well developed, arising ventrally from the anterior margin of the metacoxal foramen (cx3f). In most Bethylidae the metacoxal foramina are closed, but in some they are continuous with the propodeal foramen (prf: Figs 26A, 26C, 26D View FIGURE 26 ), forming a single opening ( Fig. 26D View FIGURE 26 ). The propodeal foramen is surrounded by a raised area, the nucha (nu: Figs 25 View FIGURE 25 A–C, 26E). The propodeal articular process* (papr: Figs 26B, 26D, 26E View FIGURE 26 ) is located on the lateral margin of the propodeal foramen and fits into the metasomal articular process* (map: Figs 34A and B View FIGURE 34 ).
The anterior margin of the ventral surface of the metapectus has the paracoxal sulcus (pcs: Figs 25B View FIGURE 25 , 26B View FIGURE 26 ) that fits the posterior margin of the ventral surface of the mesopectus. Internally, the paracoxal sulcus is represented by the paracoxal ridge (pcr: Fig. 26F View FIGURE 26 ). Ventrally, the metapectus forms a large plate between the mesocoxae, the metasternal plate* (s3p: Figs 26A, 26D View FIGURE 26 ). The metadiscrimen (dc3: Fig. 26B View FIGURE 26 ), when present, is represented by a longitudinal sulcus located ventrally on the metasternum. The central or posterior area of the metasternum has a single metafurcal pit (fu3p: Figs 26 View FIGURE 26 B–D). Internally, the metadiscrimen and metafurcal pit represent the internal metadiscriminal lamella (dc3l) and the metafurca (fu3), respectively. The base of the metafurca arises close to the posterior surface of the paracoxal ridge and is divided into two metafurcal arms (fu3a: Fig. 26F View FIGURE 26 ) that extend laterally to fuse with the metapleural apodeme. The metafurcal arm is medially enlarged, forming the dorsal metafurcal lamella (fu3dl).
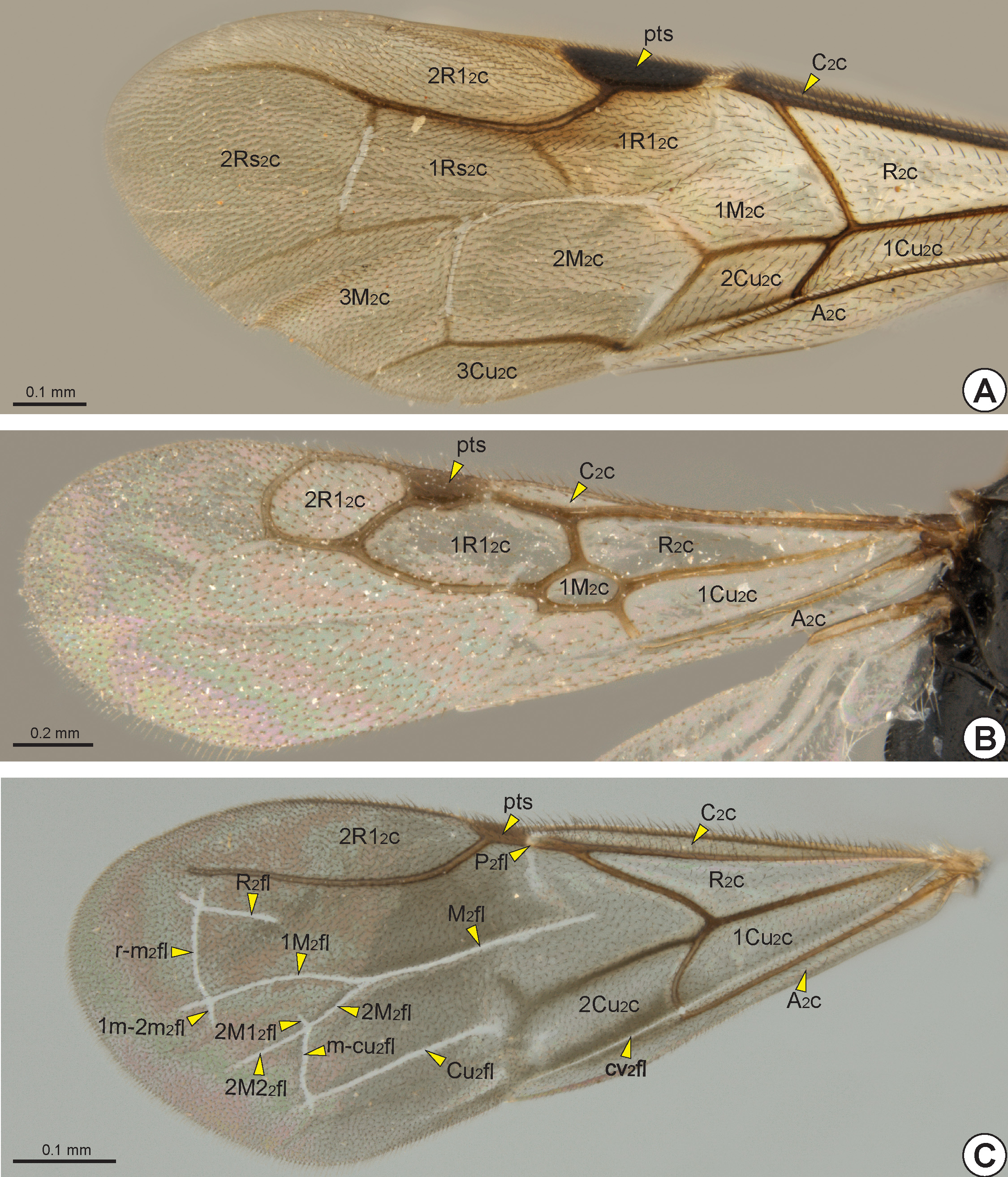
Wings: Most Bethylidae have well-developed wings (macropterous), but some species may have the wings moderately reduced (brachypterous), strongly reduced (micropterous), or even absent (apterous). Compared with most other aculeates, the wing veins are reduced in extent; they may be tubular, nebulous or spectral (see Mason 1986 for definitions).
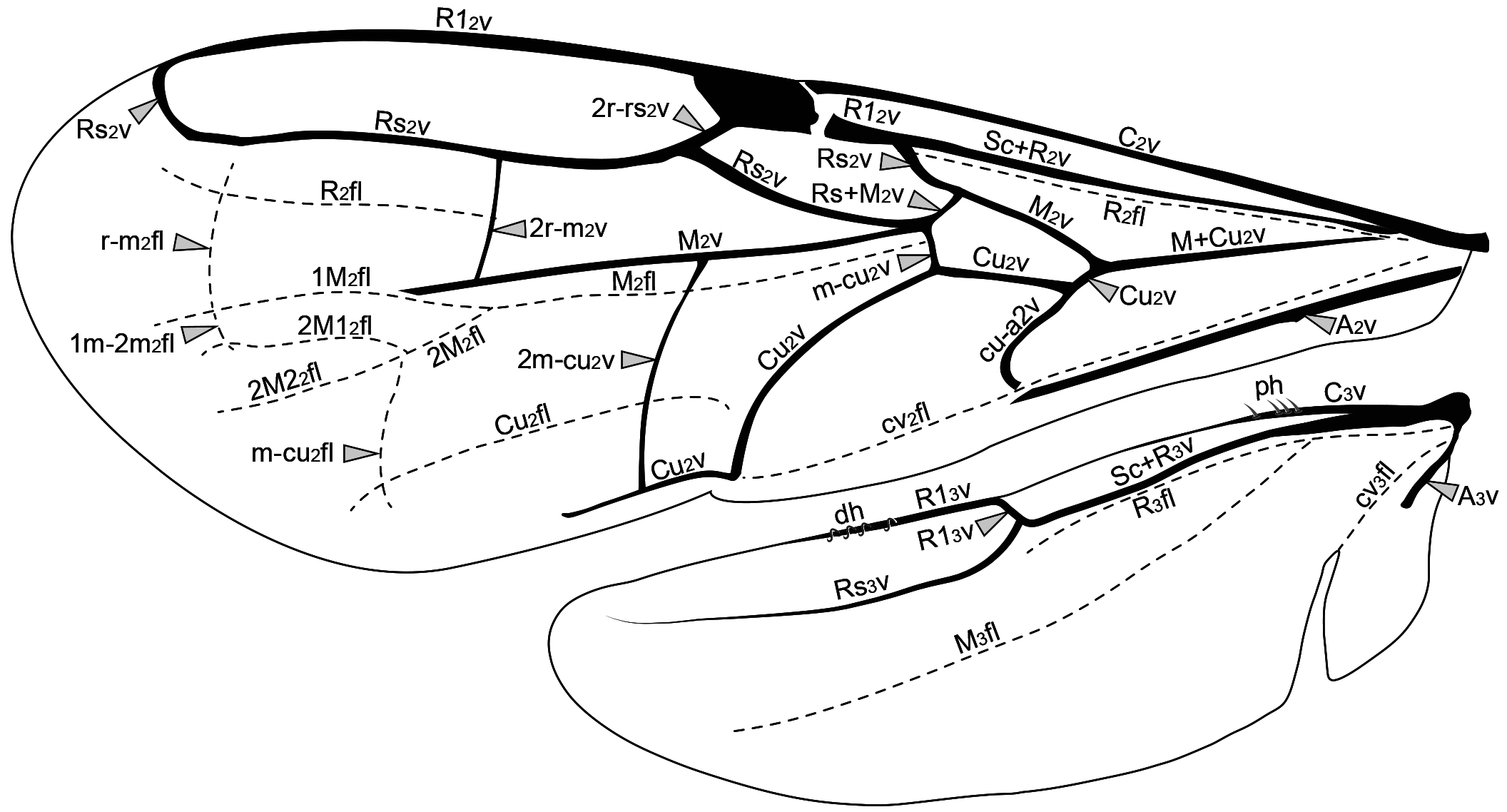
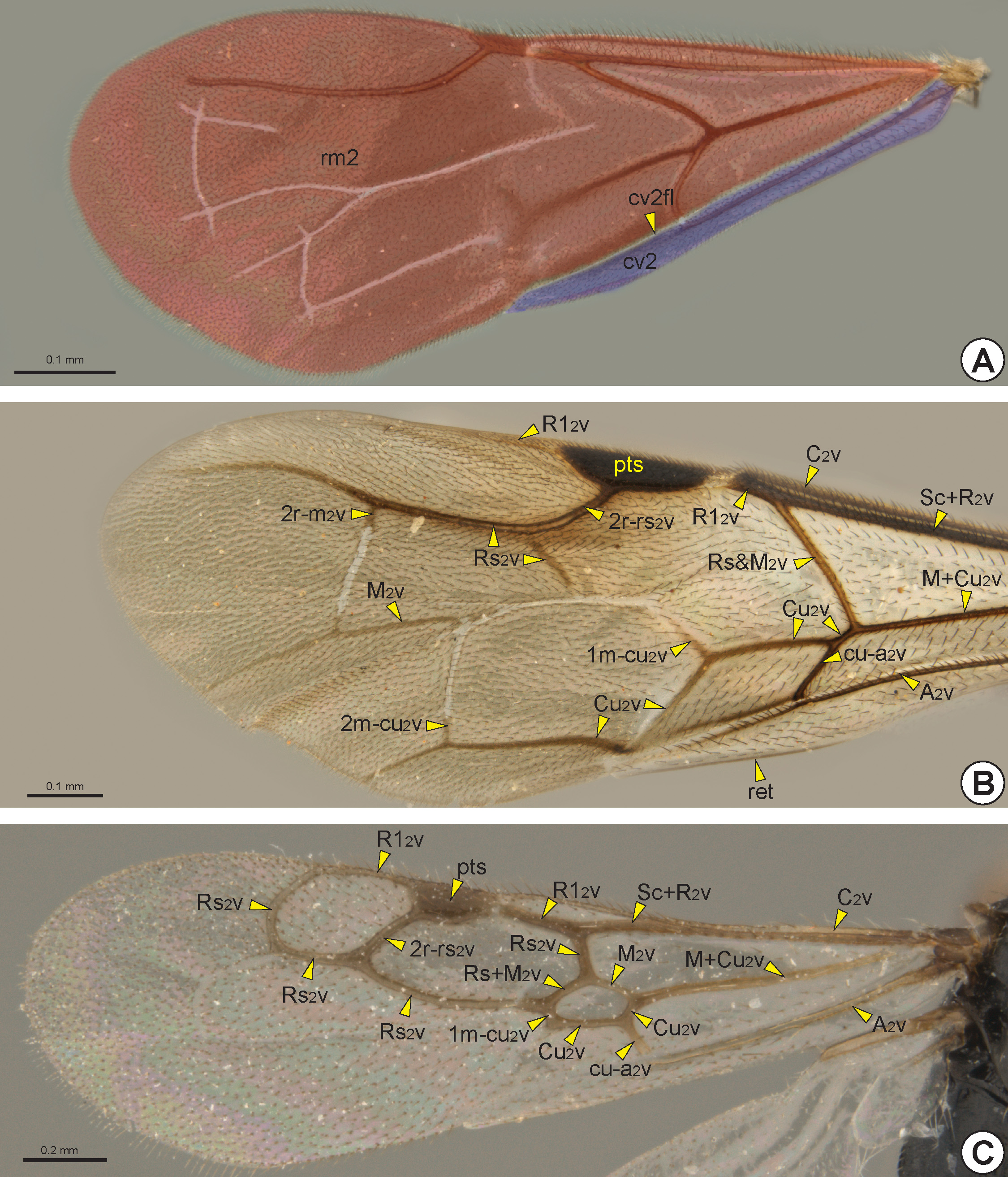
The fore wing of macropterous specimens is divided into remigium of the fore wing (rm2: Fig. 28A View FIGURE 28 ), clavus
of the fore wing (cv2: Fig. 28A View FIGURE 28 ), and jugum of the fore wing (ju2). The remigium comprises the largest region, from the anterior margin to the claval flexion line of the fore wing* (cv2fl: Figs 27 View FIGURE 27 , 28A View FIGURE 28 , 30C View FIGURE 30 ). The clavus comprises the area posterior to the claval flexion line, and includes the anal vein of the fore wing* (A 2 v: Figs 27 View FIGURE 27 , 28B, 28C View FIGURE 28 , 29 View FIGURE 29 ) which articulates with the third axillary sclerite of the fore wing (3ax2). The jugum is a small and membranous lobe linked to the third axillary sclerite, and only visible after dissection of the wing.
The fore wing is usually hyaline, but some species have wings with dark transverse bands. In most Bethylidae the fore wing gradually widens from the base to the apex, but in a few genera it is narrow from the base to the pterostigma (pts: Figs 28B, 28C View FIGURE 28 , 29B View FIGURE 29 , 30 View FIGURE 30 ) and then widens abruptly toward the apex.
The sclerotized area at the base of the fore wing, the humeral complex of the fore wing (hc2), is composed of the humeral plate of the fore wing (hp2) and the bases of the two anterior veins. The humeral plate is an irregular and sinuous sclerite connected basally to the mesobasalare, apically to the costal vein of the fore wing (C 2 v: Figs 27 View FIGURE 27 , 28B, 28C View FIGURE 28 , 29C View FIGURE 29 ) and posteriorly to the median plate of the fore wing (mp2), which is distinguishable from the C 2 v by a constriction. The first axillary sclerite articulates medially with the second axillary sclerite, which is posteriorly articulated with the third axillary sclerite. The third axillary sclerite is apically connected with A 2 v.
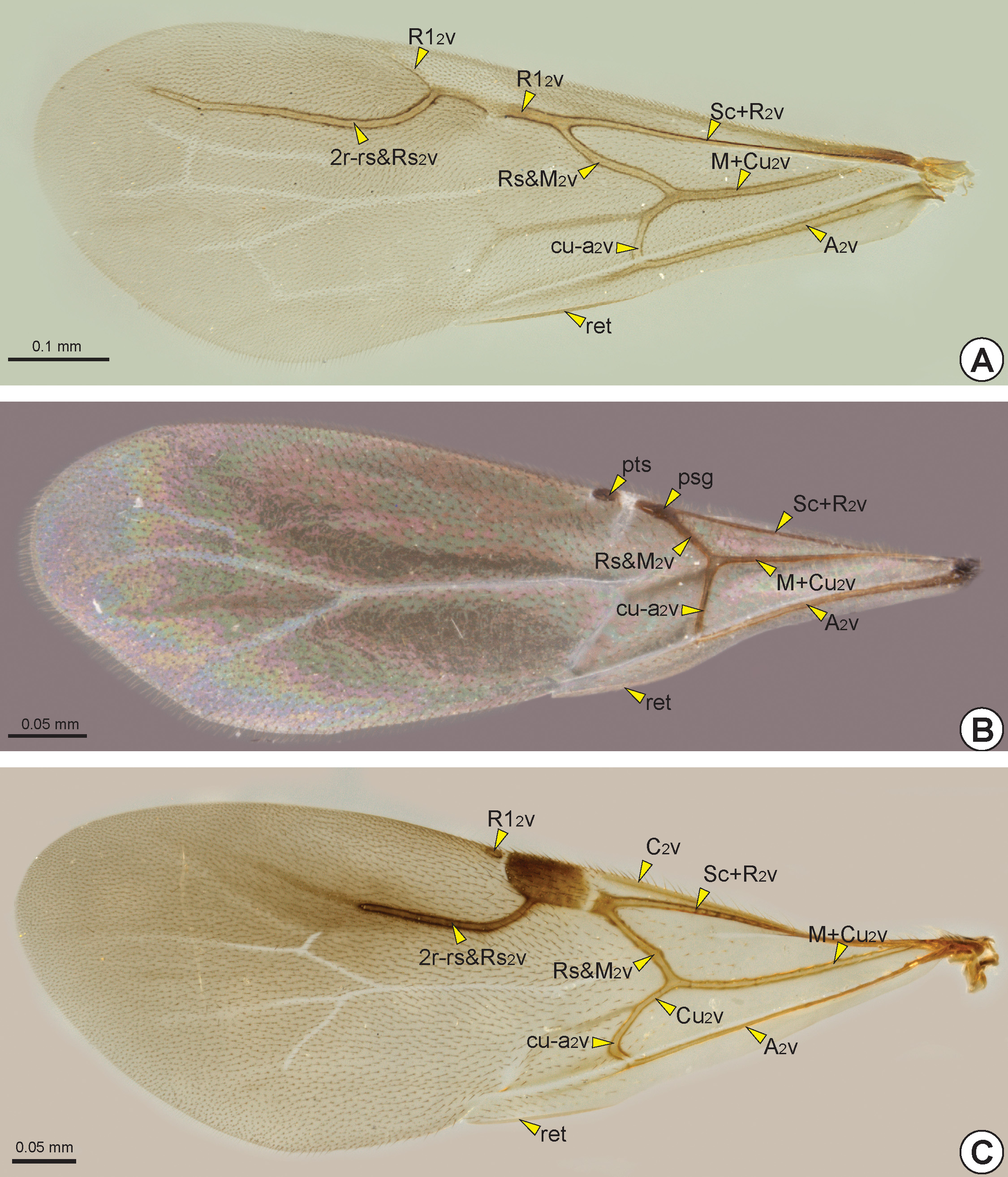
The C 2 v is tubular and is located at the anterior margin of the fore wing. Sometimes the C 2 v is very short, restricted to its base. The radial vein of the fore wing* (R 2 v) and the subcostal vein of the fore wing (Sc 2 v) are fused, forming the tubular subcostal+radial vein of the fore wing* (Sc+R 2 v: Figs 27 View FIGURE 27 , 28B, 28C View FIGURE 28 , 29 View FIGURE 29 ). The area bounded by the C 2 v and Sc+R 2 v forms the costal cell of the fore wing* (C 2 c: Fig. 30 View FIGURE 30 ). When the C 2 v and Sc+R 2 v are confluent, the C 2 c is absent or very narrow.
At the apical end of the Sc+R 2 v, the Sc 2 v is absent and the R 2 v branches into the radial 1 vein of the fore wing* (R1 2 v: Figs 27 View FIGURE 27 , 28B, 28C View FIGURE 28 , 29A, 29C View FIGURE 29 ) and radial sector vein of the fore wing* (Rs 2 v: Figs 27 View FIGURE 27 , 28B, 28C View FIGURE 28 , 29 View FIGURE 29 ). In some genera the node of the R 2 v, where it branches into R1 2 v and Rs 2 v, is dilated and commonly called the prestigma (psg: Fig. 29B View FIGURE 29 ). The R1 2 v follows toward the pterostigma, and its first abscissa is usually much reduced ( Fig. 28B View FIGURE 28 ); in only a few genera is the first abscissa of R1 2 v considerably longer ( Figs 28C View FIGURE 28 , 29A View FIGURE 29 ). The R1 2 v bounds the posterior margin of the pterostigma and, in some taxa, it follows along the anterior margin of the wing ( Figs 28B, 28C View FIGURE 28 , 29C View FIGURE 29 ). The pterostigma is wide in most Bethylidae , but it is reduced ( Fig. 29B View FIGURE 29 ) in a few and rarely absent ( Fig. 29A View FIGURE 29 ).
The median vein of the fore wing (M 2 v) and cubital vein of the fore wing (Cu 2 v) are fused to form the median+cubital vein of the fore wing* (M+Cu 2 v: Figs 27 View FIGURE 27 , 28B, 28C View FIGURE 28 , 29 View FIGURE 29 ), which is generally tubular, but it may be nebulous at the base. At the apical end of the M+Cu 2 v, the M 2 v is separated from the Cu 2 v and deflected toward the anterior margin of the wing, joining the end of the Rs 2 v. The structure formed by the apical union of the M 2 v and Rs 2 v is generally considered as a single vein, the radial sector & median vein of the fore wing* (Rs&M 2 v: Figs 27 View FIGURE 27 , 28B View FIGURE 28 , 29 View FIGURE 29 ), but sometimes the two components are separately identifiable ( Fig. 28C View FIGURE 28 ). The Sc+R 2 v, M+Cu 2 v and Rs&M 2 v delimit the radial cell of the fore wing* (R 2 c: Fig. 30 View FIGURE 30 ). At about the middle of the Rs&M 2 v, both M 2 v and Rs 2 v deflect toward the apical margin, fuse with each other and form the radial sector + median vein of the fore wing (Rs+M 2 v: Figs 27 View FIGURE 27 , 28C View FIGURE 28 ). In most Bethylidae the Rs+M 2 v is absent ( Figs 28B View FIGURE 28 , 29 View FIGURE 29 ), but in some it may be partially or totally tubular.
At the apical end of the Rs+M 2 v, the Rs 2 v and M 2 v are separated from each other, forming the third abscissa of the Rs 2 v and the fourth abscissa of the M 2 v. The third abscissa of the Rs 2 v is usually spectral or absent, although it is sometimes tubular and extends towards the anteroapical margin of the fore wing ( Figs 28B, 28C View FIGURE 28 ). The Rs 2 v is connected to the R1 2 v by the second radial cross vein of the fore wing* (2r-rs 2 v: Figs 28B, 28C View FIGURE 28 ). This cross vein is present in most Bethylidae , even in species where the third abscissa of Rs 2 v is absent ( Figs 29A, 29C View FIGURE 29 ). In a few, R1 2 v, Rs 2 v, Rs+M 2 v and 2r-rs 2 v, form the first radial 1 cell of the fore wing* (1R1 2 c: Figs 30A, 30B View FIGURE 30 ). In most Bethylidae the 1R1 2 c is absent ( Fig. 30C View FIGURE 30 ) or open ( Fig. 30A View FIGURE 30 ). Posterior to the 2r-rs 2 v, the fourth abscissa of the Rs 2 v
usually goes towards the apical margin. Where the third abscissa of Rs 2 v is absent, the segment formed by the union between the 2r-rs 2 v and the fourth abscissa of the Rs 2 v appears as a single tubular vein, the second radial cross vein & radial sector vein of the fore wing* (2r-rs&Rs 2 v: Figs 29A, 29C View FIGURE 29 ). In a few the 2r-rs 2 v is present, but the fourth abscissa of the Rs 2 v is absent; in others the fifth abscissa of Rs 2 v can deflect toward the anterior margin, and then sometimes it is long and connects to the R1 2 v, forming the closed second radial 1 cell of the fore wing* (2R1 2 c: Fig. 30B View FIGURE 30 ). In most Bethylidae the 2R1 2 c is absent or open ( Figs 30A, 30C View FIGURE 30 ). Although weakly defined, some species may have two cells delimited anteriorly by the r-rs&Rs 2 v and posteriorly by the M 2 v, the first radial sector cell of the fore wing* (1Rs 2 c: Fig. 30A View FIGURE 30 ) and second radial sector cell of the fore wing* (2Rs 2 c: Fig. 30A View FIGURE 30 ).
Distal to the Rs+M 2 v, the fourth abscissa of the M 2 v is usually spectral or nebulous and goes toward the apical margin ( Fig. 28B View FIGURE 28 ). In a few species, a transverse vein connects Rs 2 v and M 2 2v distally, the second radio-median cross vein of the fore wing * (2r-m 2 v: 27, Fig. 28B View FIGURE 28 ). Distal to the M+Cu 2 v, the second abscissa of Cu 2 v is short and tubular ( Fig. 29C View FIGURE 29 ), but its third abscissa becomes spectral or nebulous and can deflect toward the posterior margin
of the wing and deflect again toward the apical margin ( Fig. 28B View FIGURE 28 ). In some genera, the Cu 2 v is connected to the M 2 v by the first medio-cubital cross vein of the fore wing* (1m-cu 2 v: Figs 28B, 28C View FIGURE 28 ). In only a few species, a second transverse vein connects M 2 v and Cu 2 v distally, the second medio-cubital cross vein of the fore wing* (2m-cu 2 v: Fig. 28B View FIGURE 28 ). Rarely, the M 2 v, Cu 2 v, Rs+M 2 v and m-cu 2 v delimit the first median cell of the fore wing* (1M 2 c: Figs 30A, 30B View FIGURE 30 ). A few species have the second median cell of the fore wing* (2M 2 c: Fig. 30A View FIGURE 30 ) and third median cell of the fore wing* (3M 2 c: Fig. 30A View FIGURE 30 ), which are open or poorly defined.
The A 2 v is long, almost reaching the posteroapical margin of the fore wing. The basal half of A 2 v is tubular, whereas the apical half is usually nebulous, this second half is often confluent with the claval flexion line and therefore difficult to identify. The tubular abscissa of the A 2 v connects to the Cu 2 v by the cubito-anal cross vein of the fore wing* (cu-a 2 v: Figs 27 View FIGURE 27 , 28B, 28C View FIGURE 28 , 29 View FIGURE 29 ). The first cubital cell of the fore wing* (1Cu 2 c: Fig. 30 View FIGURE 30 ) is delimited by the M+Cu 2 v, the second abscissa of Cu 2 v, A 2 v and cu-a 2 v. In some species, the A 2 v, Cu 2 v and cu-a 2 v delimit the second cubital cell of the fore wing* (2Cu 2 c: Figs 30A, 30C View FIGURE 30 ), that is located distal to the 1Cu 2 c. When the Cu 2 v is long and reaches the posteroapical margin of the fore wing, it anteriorly delimits the third cubital cell of the fore wing* (3Cu 2 c: Fig. 30A View FIGURE 30 ). Posterior to the A 2 v, the anal cell of the fore wing* (A 2 c: Fig. 30 View FIGURE 30 ) is always open, and is posteriorly limited by the posterior margin of the fore-wing.
Rarely, due to the absence of the M+Cu 2 v, the R 2 c and 1Cu 2 c are fused, forming a single cell, or the M+Cu 2 v is basally absent and the R 2 c and 1Cu 2 c are basally fused. In some, the distinction between the Cu 2 v and cu-a 2 v is obscure and the segment formed by the union of these two veins can be straight ( Fig. 29B View FIGURE 29 ), curved towards the posteroapical margin ( Fig. 28B View FIGURE 28 ) or obliquely facing towards the posterobasal margin.
In Bethylidae , the radial flexion line of the fore wing* (R 2 fl: Figs 27 View FIGURE 27 , 30C View FIGURE 30 ) runs parallel and immediately posterior to Sc+R 2 v. The R 2 fl is convex, so that the membranous area anterior to this line is relatively ventrally oriented, making it difficult to see in dorsal view. The R 2 fl is usually interrupted medially, separating into two distinct abscissae. The apical end of the first abscissa of the R 2 fl curves anteriorly towards the base of the pterostigma, where it is contiguous with the hinge-like mechanism, a weakened area at the pterostigmal base, the prestigmal flexion line of the fore wing* (P 2 fl: Fig. 30C View FIGURE 30 ). When present, the second abscissa of the R 2 fl is located posterior to the r-rs&Rs 2 v ( Fig. 30C View FIGURE 30 ).
The median flexion line of the fore wing* (M 2 fl: Figs 27 View FIGURE 27 , 30C View FIGURE 30 ) is a longitudinal flexion line that arises distal to the Rs+M 2 v or where this vein would be located. In some, the M 2 fl is continuous and curved to the anterior margin of the wing. In many genera, the M 2 fl branches into the anterior median flexion line of the fore wing* (1 M 2 fl: Figs 27 View FIGURE 27 , 30C View FIGURE 30 ) and posterior median flexion line of the fore wing* (2M 2 fl: Figs 27 View FIGURE 27 , 30C View FIGURE 30 ), which extend towards the anteroapical and posteroapical margins, respectively. The 2M 2 fl is usually subdivided into the first posterior median flexion line of the fore wing* (2M1 2 fl: Figs 27 View FIGURE 27 , 30C View FIGURE 30 ) and second posterior median flexion line of the fore wing* (2M2 2 fl: Figs 27 View FIGURE 27 , 30C View FIGURE 30 ).
The cubital flexion line of the fore wing* (Cu 2 fl: Figs 27 View FIGURE 27 , 30C View FIGURE 30 ) is absent in many Bethylidae , but, when present, it arises distal to the third abscissa of the Cu 2 v and extends towards the posteroapical margin of the wing.
Several genera have the transverse radial-median cross flexion line of the fore wing* (r-m 2 fl: Figs 27 View FIGURE 27 , 30C View FIGURE 30 ) connecting the R 2 fl to the M 2 fl. In specimens with the M 2 fl branched, the r-m 2 fl connects the R 2 fl to the 1M 2 fl. In some species, the 1M 2 fl and 2M 2 fl are connected by the transverse anterior-posterior median cross flexion line of the fore wing* (1m- 2m 2 fl: Figs 27 View FIGURE 27 , 30C View FIGURE 30 ). The medio-cubital cross flexion line of the fore wing* (m-cu 2 fl: Figs 27 View FIGURE 27 , 30C View FIGURE 30 ) which connects 2M 2 fl and Cu 2 fl is usually present, even in species that do not have the Cu 2 fl.
The claval flexion line of the fore wing is concave, so that the membranous area posterior to this line is relatively dorsally oriented. The retinaculum (ret: Figs 28B View FIGURE 28 , 29 View FIGURE 29 ) is the area of the posterior margin that is ventrally curved and accommodates the distal hamuli (dh: Figs 27 View FIGURE 27 , 31B View FIGURE 31 ) of the hind wing.
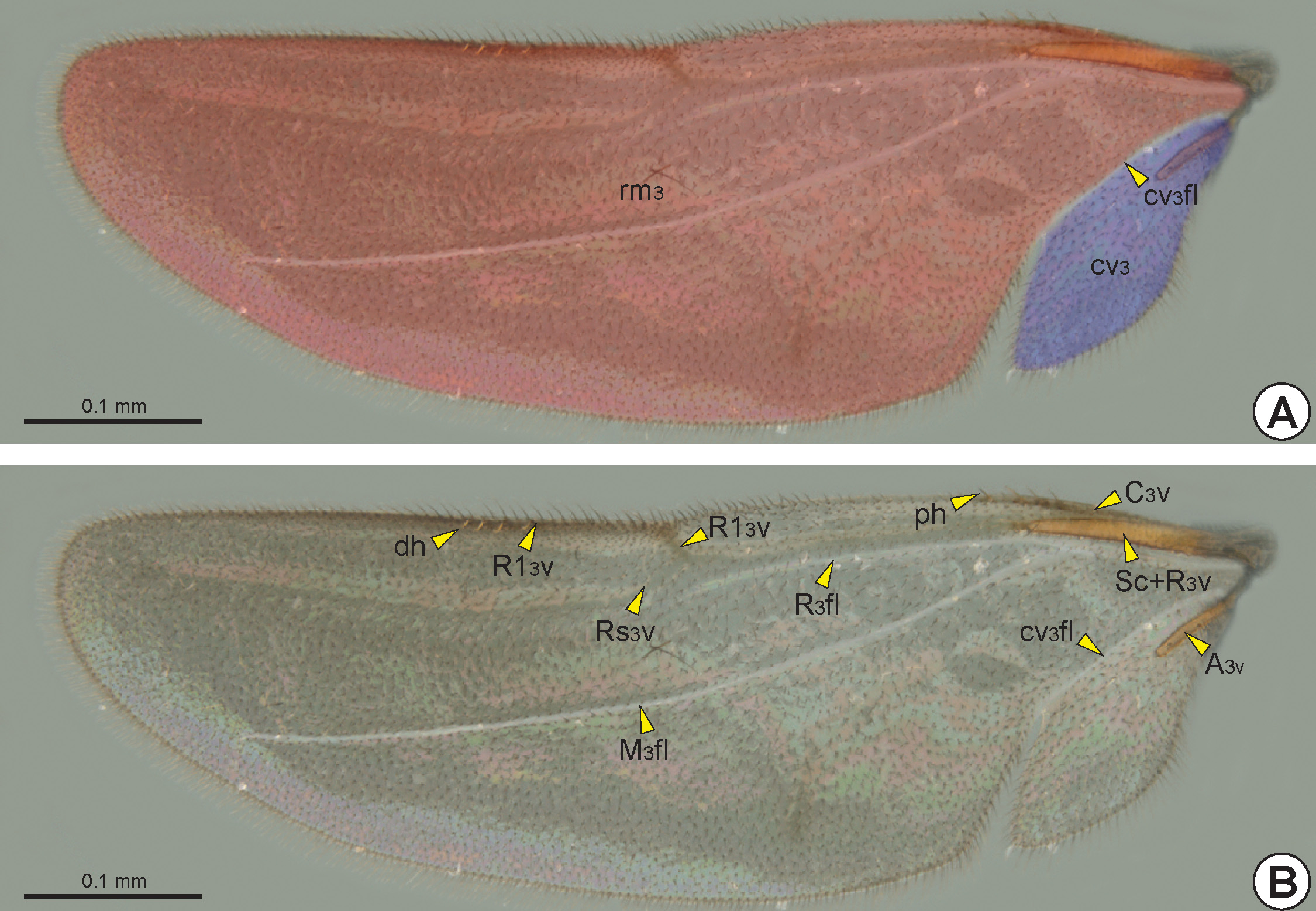
The hind wing of macropterous specimens is divided into remigium of the hind wing (rm3: Fig. 31A View FIGURE 31 ), clavus of the hind wing (cv3: Fig. 31A View FIGURE 31 ), and jugum of the hind wing (ju3). The remigium comprises the largest region of the hind wing, from the anterior margin to the claval flexion line of the hind wing* (cv 3 fl: Figs 27 View FIGURE 27 , 31 View FIGURE 31 ). The clavus comprises the area posterior to the claval flexion line, and includes the short and tubular anal vein of the hind wing* (A 3 v: Figs 27 View FIGURE 27 , 31B View FIGURE 31 ) which articulates with the third axillary sclerite of the hind wing (3ax3). The jugum is a small and membranous lobe linked to the third axillary sclerite, and is visible only after dissection of the hind wing.
Similar to the fore wing, the humeral complex of the hind wing (hc3) is composed of the humeral plate of the hind wing (hp3) and the base of the two anterior veins. As in the fore wing, the humeral plate of the hind wing is irregular and sinuous, and it can be interpreted as two distinct sclerites. The humeral plate is connected basally to the metabasalare and apically to the subcostal+radial vein of the hind wing* (Sc+R 3 v: Figs 27 View FIGURE 27 , 31B View FIGURE 31 ). The median plate of the hind wing* (mp3) articulates anteromedially with the humeral plate and posteriorly with the A 3 v. The first axillary sclerite of the hind wing (1ax3) is F-shaped. The anterior arm of the first axillary sclerite is connected to the humeral plate, and the posterior arm is connected to the second axillary sclerite of the hind wing* (2ax3). The mesobasal margin and the posterobasal end of the first axillary sclerite are connected to the postmedian metanotal wing process. The second axillary sclerite articulates apically with the humeral plate and posteriorly with the third axillary sclerite. The third axillary sclerite is basally articulated with the metapleural wing process and apically with the A 3 v.
The costal vein of the hind wing (C 3 v: Figs 27 View FIGURE 27 , 31B View FIGURE 31 ) is short and less sclerotized than the other veins of the hind wing. The radial vein of the hind wing* (R 3 v) and the subcostal vein of the hind wing (Sc 3 v) are fused, forming the subcostal+radial vein. The Sc+R 3 v may extend to approximately half the hind-wing length. The first half of Sc+R 3 v is tubular, but the second half is spectral or nebulous. At the apical end of the Sc+R3v, the Sc 3 v is absent and the R 3 v branches into the radial 1 vein of the hind wing* (R1 3 v: Fig. 27 View FIGURE 27 , 31B View FIGURE 31 ) and the radial sector vein of the hind wing* (Rs 3 v: Figs 27 View FIGURE 27 , 31B View FIGURE 31 ). The R1 3 v is initially nebulous and follows along the anterior margin of the hind wing, while the Rs 3 v is spectral and is usually only visible apically.
When present, the median+cubital vein of the hind wing* (M+Cu 3 v) is nebulous or spectral. At the apical end of the M+Cu 3 v, the cubital vein of the hind wing (Cu 3 v) is separated from the median vein of the hind wing (M 3 v). The Cu 3 v and M 3 v are spectral or nebulous, with Cu 3 v following diagonally towards the posteroapical margin and the M 3 v generally visible only apically.
One to three straight proximal hamuli (ph: Figs 27 View FIGURE 27 , 31B View FIGURE 31 ) arise on the anterior margin of the hind wing apical to the tubular segment of the Sc+R 3 v. They are often difficult to identify in wings with the surface covered with erect setae. Medio-apically, the anterior margin of the hind wing has from three to eleven curved distal hamuli (dh: Figs 27 View FIGURE 27 , 31 View FIGURE 31 ) which engage with the retinaculum of the fore wing.
The radial flexion line of the hind wing* (R 3 fl: Figs 27 View FIGURE 27 , 31 View FIGURE 31 ) runs parallel and immediately posterior to Sc+R 3 v. The median flexion line of the hind wing* (M 3 fl: Figs 27 View FIGURE 27 , 31 View FIGURE 31 ) originates near the R 3 fl and runs diagonally and immediately anterior to the M+Cu 3 v and M 3 v. The claval flexion line of the hind wing is anterior to the A 3 v and extends to the posteroapical margin, where there is usually an incision that separates the clavus from the remigium.
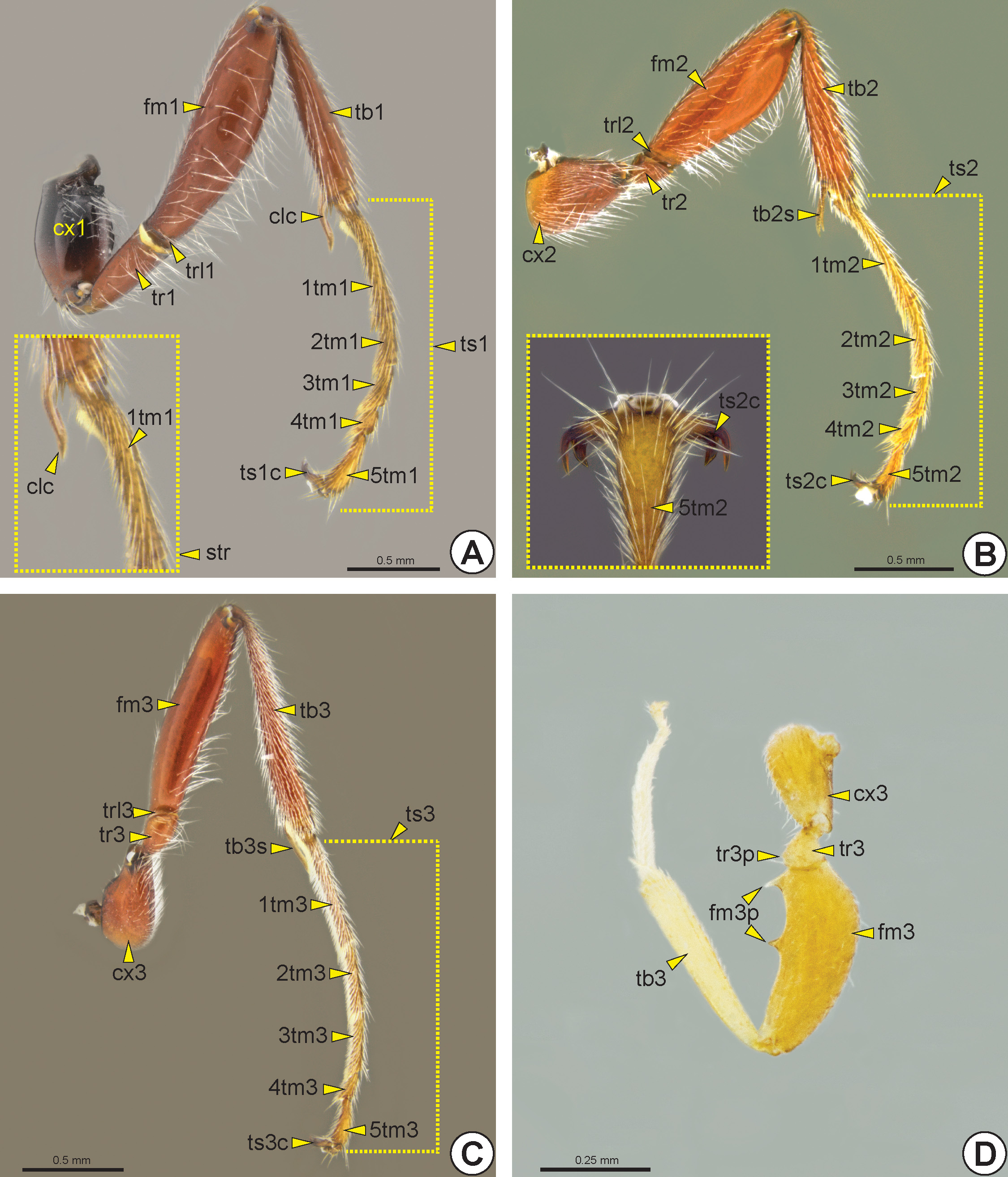
Proleg (lg1: Fig. 32A View FIGURE 32 ): The procoxa is slightly longer than wide. The protrochanter (tr1: Fig. 32A View FIGURE 32 ) varies in length. The protrochantellus (trl1: Fig. 32A View FIGURE 32 ) is weakly evident. The profemur (fm1: Fig. 32A View FIGURE 32 ) varies in width, length and thickness, and can be conspicuously dilated in some genera. The protibia (tb1: Fig. 32A View FIGURE 32 ) may have spines at the apex. The protarsus (ts1: Fig. 32A View FIGURE 32 ) is divided into five protarsomeres (1-5tm1: Fig. 32A View FIGURE 32 ). The first protarsomere, the probasitarsus (1tm1: Fig. 32A View FIGURE 32 ), is slightly curved along the calcar (clc: Fig. 32A View FIGURE 32 ). The probasitarsus and calcar together constitute the antennal-cleaning apparatus, the strigil (str: Fig. 32A View FIGURE 32 ). Most protarsomeres have spines that vary in size and shape, usually conic, but rarely flattened. The protarsal claw (ts1c: Fig. 32A View FIGURE 32 ), which can be simple, bifid or trifid, arises laterally on the propretarsus (prt1).
Mesoleg (lg2: Fig. 32B View FIGURE 32 ): The mesocoxa (cx2) is usually longer than wide. The mesotrochanter (tr2: Fig. 32B View FIGURE 32 ) varies in length. The mesotrochantellus (trl2: Fig. 32B View FIGURE 32 ) is weakly evident. The mesofemur (fm2: Fig. 32B View FIGURE 32 ) varies in width, length and thickness. The mesotibia (tb2: Fig. 32B View FIGURE 32 ) usually has a pair of mesotibial spurs (tb2s: Fig. 32B View FIGURE 32 ), which may differ in length from each other. The mesotibia may have spines apically. The mesotarsus (ts2: Fig. 32B View FIGURE 32 ) is divided into five mesotarsomeres (1-5tm2: Fig. 32B View FIGURE 32 ). The mesobasitarsus (1tm2: Fig. 32B View FIGURE 32 ) and other mesotarsomeres have spines that vary in size and shape. The mesotarsal claw (ts2c: Fig. 32B View FIGURE 32 ), which can be simple, bifid or trifid, arises laterally on the mesopretarsus (prt2).
Metaleg (lg3: Figs 32C, 32D View FIGURE 32 ): The metacoxa (cx3: Figs 32C, 32D View FIGURE 32 ) is usually longer than wide. A few species have a metatrochanteral process* (tr3p: Fig. 32D View FIGURE 32 ), in the form of a tubercle or spine, on the ventral margin of the metatrochanter (tr3: Figs 32C, 32D View FIGURE 32 ). The metatrochantellus (trl3: Fig. 32C View FIGURE 32 ) is weakly evident. The metafemur (fm3: Figs 32C, 32D View FIGURE 32 ) varies in width, length and thickness, rarely with one or two metafemoral process es * (fm3p: Fig. 32D View FIGURE 32 ) in the form of tubercles or spines on the ventral margin. The metatibia (tb3: Figs 32C, 32D View FIGURE 32 ) has a pair of metatibial spurs (tb3s: Fig. 32C View FIGURE 32 ) that generally differ in length from each other. The metatibia can have spines apically. The metatarsus (ts3: Fig. 32C View FIGURE 32 ) is divided into five metatarsomeres (1-5tm3: Fig. 32C View FIGURE 32 ). The metabasitarsus (1tm3: Fig. 32C View FIGURE 32 ) varies in length, and in some species it may be as long as the metatibia. The metatarsomeres may have spines that vary in size and shape. The metatarsal claw (ts3c: Fig. 32C View FIGURE 32 ), which can be simple, bifid or trifid, arises laterally on the metapretarsus (prt3).
Metasoma. The metasoma (mtm: Fig. 33 View FIGURE 33 ) has seven or eight abdominal segments externally visible, from the second (A2) to the eighth abdominal segment (A8) in females ( Fig. 33B View FIGURE 33 ), and from the second to ninth abdominal segment (A9) in males ( Fig. 33A View FIGURE 33 ).
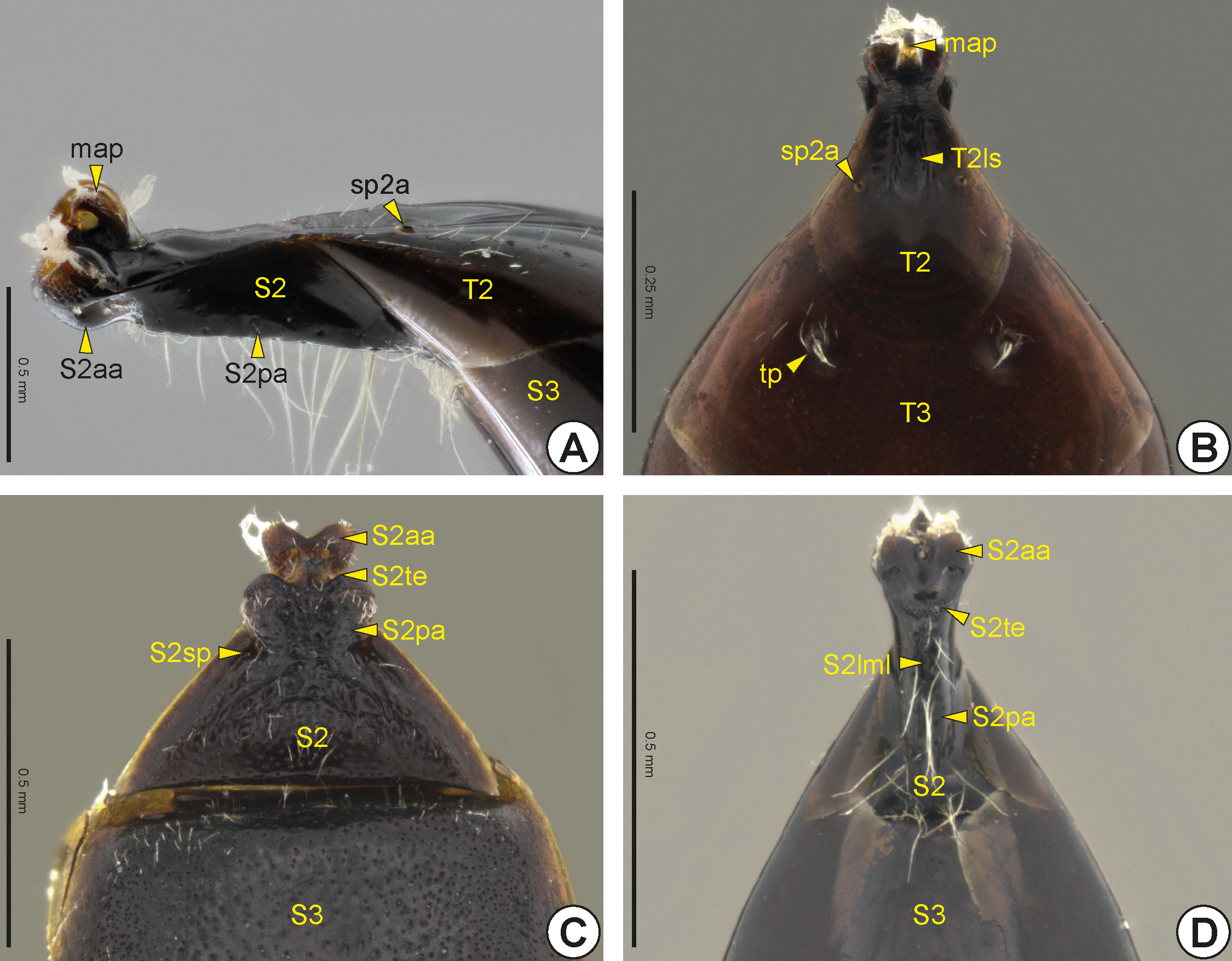
In both sexes, the second abdominal segment is anteriorly slender and cylindrical, and connects to the propodeal foramen. In some genera this anterior narrowing of the second abdominal segment is short and almost indistinguishable ( Figs 34B, 34C View FIGURE 34 ), whereas in others this narrow area is easily identifiable ( Figs 34A, 34D View FIGURE 34 ); its components are very closely associated. The second abdominal tergum (T2: Figs 33C View FIGURE 33 , 34A, 34B View FIGURE 34 ) and second abdominal sternum (S2: Figs 33D View FIGURE 33 , 34A, 34C, 34D View FIGURE 34 ) are narrower anteriorly but widen posteriorly to connect to the anterior margin of the third abdominal segment, T2 overlapping S2 laterally. On the anterior portion of the second abdominal tergum is the metasomal articular process (map: Figs 34A, 34B View FIGURE 34 ), which has an arcuate shape and articulates with the metasomal condyles of the propodeum. The dorsal surface of the second abdominal tergum usually has a longitudinal sulcus of the T2* (T2ls: Fig. 34B View FIGURE 34 ) that corresponds internally to a longitudinal ridge of the T2 * (T2lr). The second abdominal spiracle (sp2a: Figs 34A, 34B View FIGURE 34 ) is usually located medio-laterally on the second abdominal tergum. In many Bethylidae the second abdominal sternum is divided into the anterior area of the S2* (S2aa: Figs 34A, 34C, 34D View FIGURE 34 ) and the posterior area of the S2* (S2pa: Figs 34A, 34C, 34D View FIGURE 34 ), which may be partially or totally separated by the transverse edge of the S2* (S2te: Figs 34C, 34D View FIGURE 34 ). The anterior area of S2 is generally gibbous and has the anterior margin rounded or medially invaginated, whereas the posterior area is usually flat and widens posteriorly. In some species, the anterior area of S2 has a longitudinal median line of the S2* (S2lml: Fig. 34D View FIGURE 34 ), which can be represented by a groove or carina. In several, the S2 is not divided into anterior and posterior areas, but the posterior portion of this sternum bends toward the tergum, being difficult to distinguish externally. Sublaterally, the S2 has the sublateral pit of the S2* (S2sp: Fig. 34C View FIGURE 34 ).
Some females have a pair of longitudinal fissures from the posterior margin of the fourth to sixth abdominal sterna (S4, S5, S6), the longitudinal fissure of the S4* (S4lf: Fig. 35A View FIGURE 35 ), longitudinal fissure of the S5* (S5lf: Fig. 35A View FIGURE 35 ) and longitudinal fissure of the S6* (S6lf: Fig. 35A View FIGURE 35 ).In females of only one species (Galadoxa torquata), the S5 has a pair of projections directed backwards, the expansions of the S5* (S5e). A very few have a pair of acute tubercles on the fifth abdominal tergum (T5: Figs 33C View FIGURE 33 , 35B View FIGURE 35 ) and sixth abdominal tergum (T6: Figs 33C View FIGURE 33 , 35B View FIGURE 35 ), the process of the T5* (T5p: Fig. 35B View FIGURE 35 ) and process of the T6* (T6p: Fig. 35B View FIGURE 35 ). In females the eighth and ninth abdominal terga are closely associated with the genitalia and therefore are visible only after dissection of the T7.
A few males have a pair of tergal depressions, pits, bristle tufts or tubercles, which are commonly referred to as the tergal process (tp: Fig. 34B View FIGURE 34 ), on the third abdominal tergum (T3: Figs 33C View FIGURE 33 , 34B View FIGURE 34 ) or the fourth abdominal tergum (T4: Fig. 33C View FIGURE 33 ). In males, the eighth abdominal tergum (T8: Figs 33C View FIGURE 33 , 35C View FIGURE 35 ) is the last prominent tergum in the metasoma. The ninth and tenth abdominal terga are fused, forming the syntergum 9+10 (T9+10: Figs 35C, 35D View FIGURE 35 ); it is retracted beneath T8 and is almost entirely membranous, the only sclerotized parts being the anterior ends, which function as apodemes. The syntergum 9+10 is closely linked to the T8 by a membrane. The cerci (ce: Figs 35C, 35D View FIGURE 35 ) arise from the posterior margin of syntergum 9+10 and are often not visible externally. In males the ninth abdominal sternum (S9: Figs 33D View FIGURE 33 , 36 View FIGURE 36 ), usually called the hypopygium, is concave dorsally and involves the genitalia. The S9 is usually visible only after dissection of the genitalia, although S9 is rarely visible externally and then the eighth abdominal sternum (S8) is short and generally covered by the seventh abdominal sternum (S7: Fig. 33D View FIGURE 33 ). In most the anterior margin of the ninth abdominal sternum has a median stalk, the spiculum (spc: Figs 36A, 36B, 36D, 36E View FIGURE 36 ), which varies in length and width. In many species they present the anterolateral apodeme of the ninth abdominal sternum (S9ala: Fig. 36D View FIGURE 36 ), which is located in the anterolareal corner of S9 and serves for muscle insertion. In some species the S9 has a pair of sublateral stalks, the anteromedial apodeme of the ninth abdominal sternum (S9ama: Fig. 36E View FIGURE 36 ). In a few genera, the lateral margins of S9 converge, narrowing the anterior margin ( Fig. 36C View FIGURE 36 ). The posterior margin of the S9 can vary between genera; it is usually angulated, rounded ( Fig. 36E View FIGURE 36 ) or medially invaginated ( Figs 36 View FIGURE 36 A–D), sometimes with two small teeth close to each other, or bilobate with the median notch short or long ( Figs 36C, 36D View FIGURE 36 ), or even deeply invaginated and almost divided into two plates ( Figs 36A, 36B View FIGURE 36 ) with the dorsal surface of the S9 often exhibiting a fold that forms a kind of dome, the chamber of the S9* (S9c: Fig. 36B View FIGURE 36 ).
Male genitalia: Anteriorly in the genitalia, the cupula (cu: Figs 38D View FIGURE 38 , 39B, 39C, 39E, 39F View FIGURE 39 ) has an open ring shape and is more sclerotized ventrally; its dorsal area is less sclerotized and surrounds the base of the genitalia. The cupula is connected to the anterior margin of the paramere (pm: Figs 37A, 37B View FIGURE 37 , 39A View FIGURE 39 ) and to the S9 by connective tissue and muscles. In most species the cupula is present, but is vestigial in some groups and absent in others.
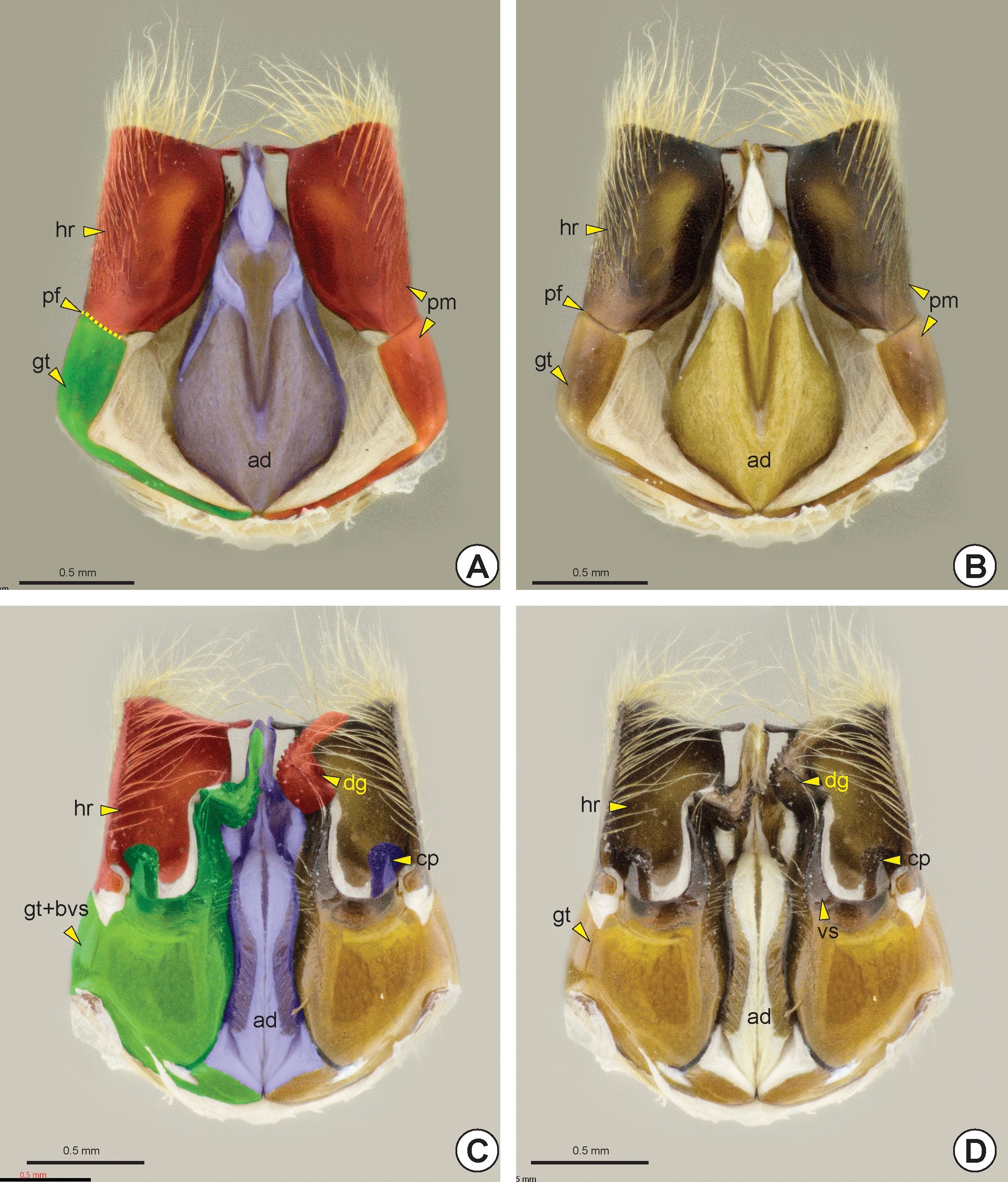
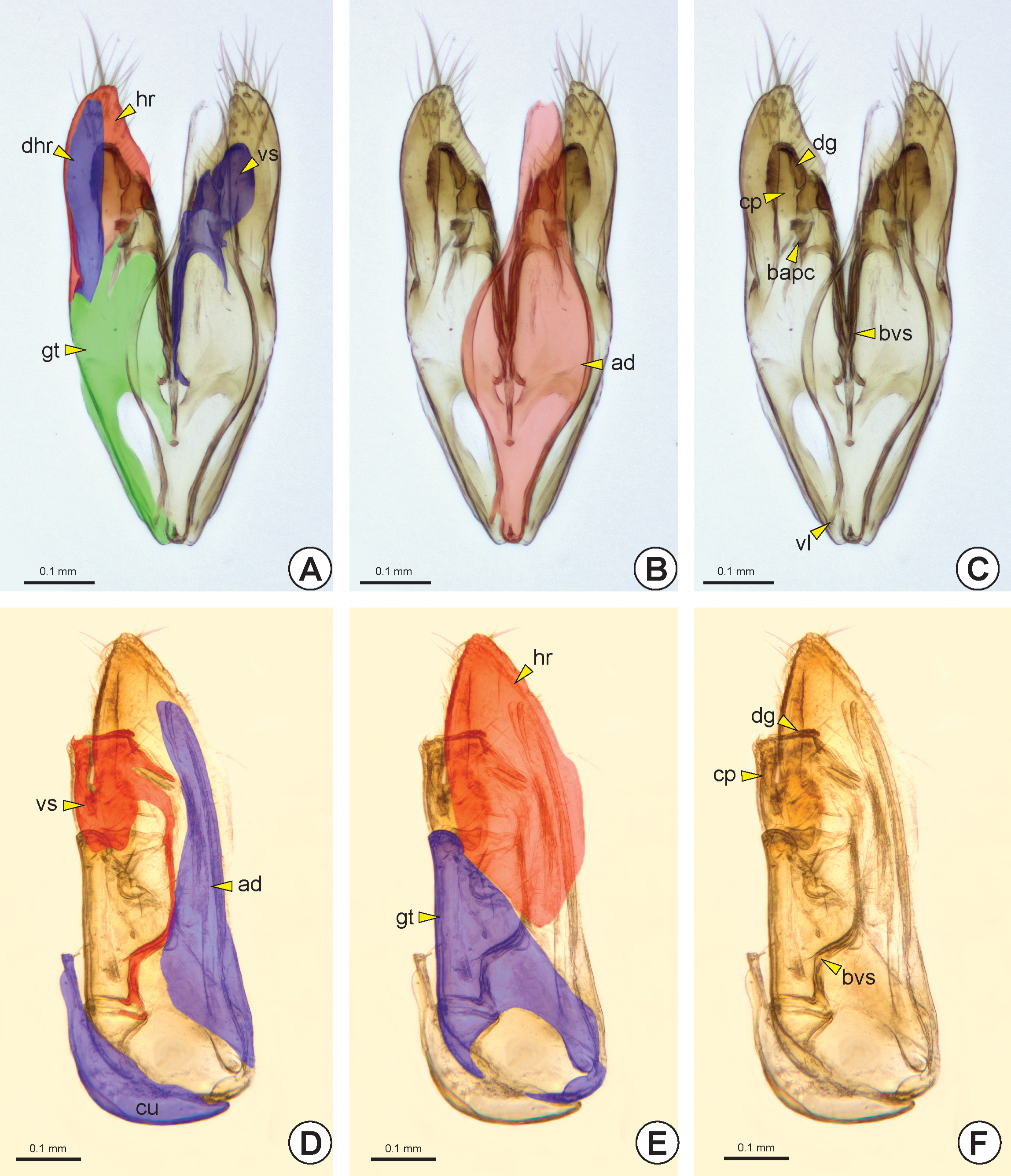
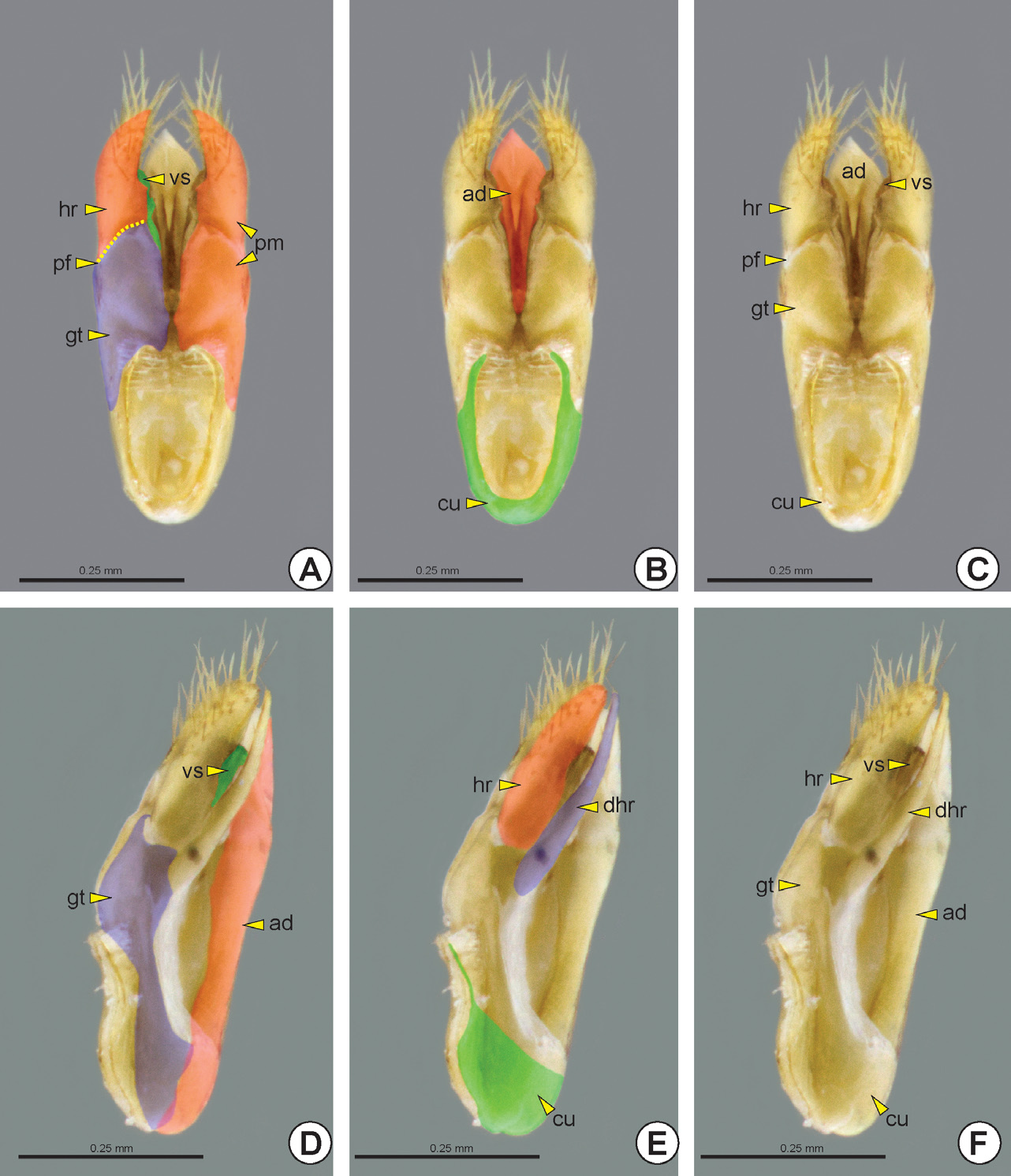
The paramere can be solid or divided into two sections by the parameral fissure* (pf: Figs 37A, 37B View FIGURE 37 , 39A, 39C View FIGURE 39 , 40B, 40C View FIGURE 40 ), delimiting an anterior gonostipes (gt: Figs 37 View FIGURE 37 , 38A, 38E View FIGURE 38 , 39A, 39C, 39D, 39F View FIGURE 39 , 40 View FIGURE 40 A–C) and a posterior harpe (hr: Figs 37 View FIGURE 37 , 38A, 38E View FIGURE 38 , 39A, 39C, 39E, 39F View FIGURE 39 , 40 View FIGURE 40 A–C). The gonostipes varies in width and length and it can be separated from the basivolsella (bvs: Figs 38C, 38F View FIGURE 38 ) or fully fused with it (gt+bvs: Fig. 37C View FIGURE 37 ). The harpe varies in length, width and shape and its basal margin usually has a concavity at the connection site with the gonostipes, the harpal chamber* (hrc). Some groups may have the harpe longitudinally subdivided, forming a second harpe on the dorsal surface of the genitalia, the dorsal harpe* (dhr: Figs 38A View FIGURE 38 , 39E, 39F View FIGURE 39 ).
The basivolsella is usually fully fused with the volsella (vs: Figs 37D View FIGURE 37 , 38A, 38D View FIGURE 38 , 39A, 39C, 39D, 39F View FIGURE 39 , 40A View FIGURE 40 ). Rarely there is a wide and sulcate lobe on the medial margin of the basivolsella, the basivolsellar vannus* (bvv). Apically, the volsella has two arms, the digitus (dg: Figs 37C, 37D View FIGURE 37 , 38C, 38F View FIGURE 38 , 40A, 40C View FIGURE 40 ) and the cuspis (cp: Figs 37C, 37D View FIGURE 37 , 38C, 38F View FIGURE 38 , 40A, 40C View FIGURE 40 ). The cuspis is the lateral arm and can comprise a double branch. Dorsally, the cus- pis has a basal projection, the basal appendicular process of the cuspis* (bapc: Figs 38C View FIGURE 38 , 40B, 40C View FIGURE 40 ), which is connected to the base of the digitus through a membrane. In some species the apex of the cuspis has an expansion directed to the base of the genitalia, the apical appendicular process of the cuspis* (aapc: see Figure 1 View FIGURE 1 in Stein & Azevedo 2007). The digitus is a median and mobile arm of the volsella and is curved toward the cuspis.
The aedeagus (ad: Figs 37 View FIGURE 37 , 38B, 38D View FIGURE 38 , 39 View FIGURE 39 B–D, 39F) is the penetration body of the male genitalia and varies in shape, width and length. The aedeagus consists of a pair of penisvalvae (pv: Fig. 40D View FIGURE 40 ), which surrounds the apical part of the ejaculatory duct, and the endophallus. In some genera the aedeagus is divided into two separate bodies: the aedeagal ventral ramus (avr) and the aedeagal dorsal body (adb); or the aedeagal ventral valve* (avv: Figs 40D, 40E View FIGURE 40 ) and aedeagal dorsal valve* (adv: Figs 40 View FIGURE 40 D–F). Rarely, the aedeagus is divided into three distinct bodies, the aedeagal ventral valve, aedeagal median valve* (amv) and aedeagal dorsal valve. In a few species, the aedeagus is simple, but its apex is bilobed. In the anterior margin of the simple aedeagus or the aedeagal dorsal structure (valve or body) there are two pairs of apodemes, valvura (vl: Figs 38C View FIGURE 38 , 40D, 40E View FIGURE 40 ) and ergot (er: Figs 40D, 40E View FIGURE 40 ).
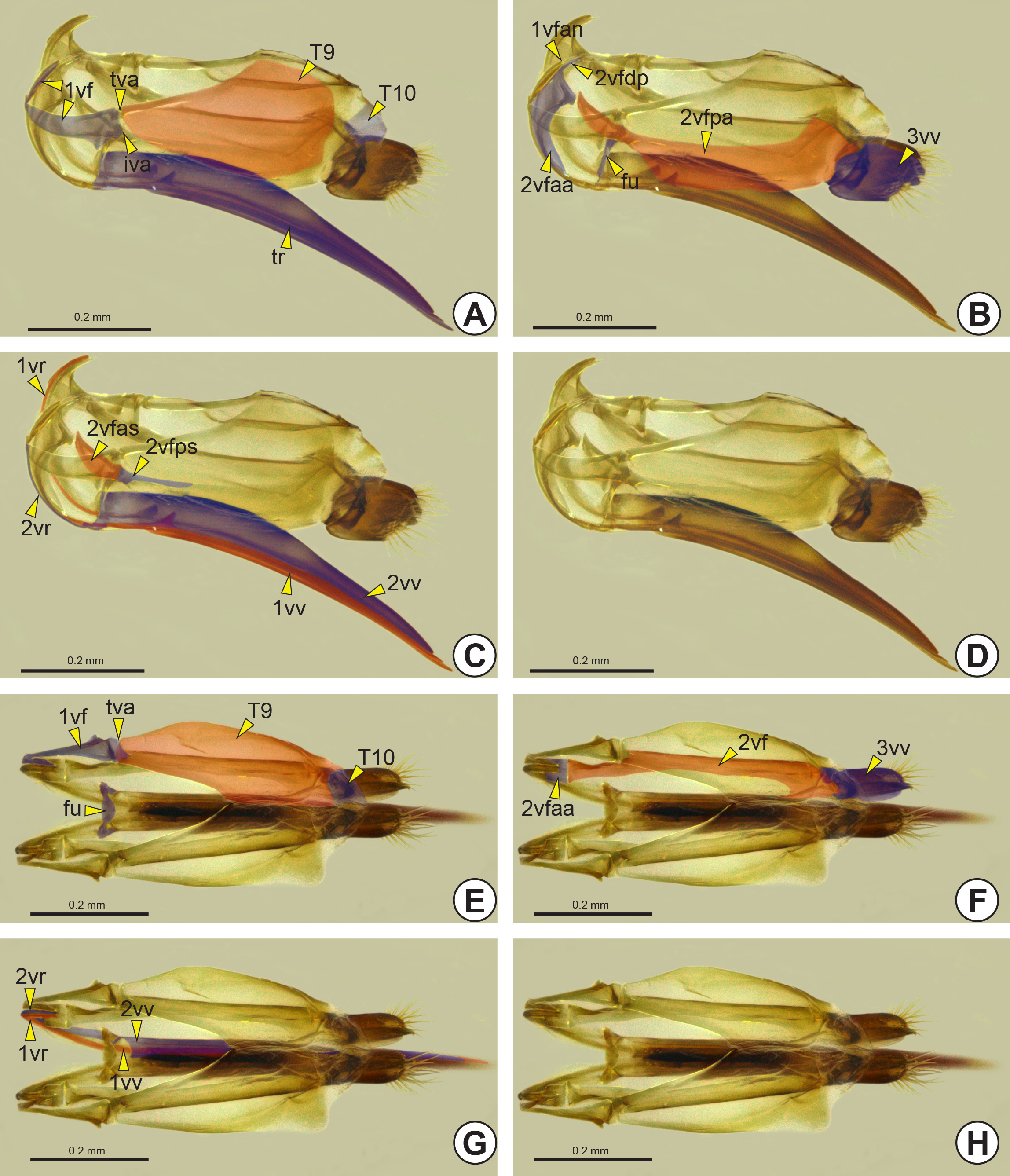
Female genitalia: The female genitalia comprise the S8, S9, median oviduct, spermatheca and ovipositor (ov). The ovipositor is an anatomical cluster consisting of the first valvifer (1vf: Fig. 41A View FIGURE 41 ), second valvifer (2vf: Fig. 41F View FIGURE 41 ), first valvula (1vv: Figs 41C, 41G View FIGURE 41 ), second valvula (2vv: Figs 41C, 41G View FIGURE 41 ), third valvula (3vv: Figs 41B, 41F View FIGURE 41 ), ninth abdominal tergum (T9: Fig. 41A View FIGURE 41 ), and tenth abdominal tergum (T10: Figure 41A View FIGURE 41 ). The portion that is exposed in the action of stinging is the terebra (tr: Figs 35B View FIGURE 35 , 41A View FIGURE 41 ) and it consists of the first and second valvulae. At the anterodorsal end of the terebra there is a small sclerite, the furcula (fu: Figs 41B, 41E View FIGURE 41 ), which may be V-shaped, U-shaped or ∆-shaped.
Anteriorly, both the first and the second valvulae are curved and thin, forming a medial first valvular ramus* (1vr: Figs 41C, 41G View FIGURE 41 ) and a lateral second valvular ramus* (2vr: Figs 41C, 41G View FIGURE 41 ), respectively. These rami form the connections between each valvula and its valvifer. The first valvifer is a triangular and short sclerite connected posterodorsally to the ninth abdominal tergum by the tergo-valvifer articulation (tva: Figs 41A, 41E View FIGURE 41 ), and posteroventrally with the second valvifer by the intervalvifer articulation (iva: Fig. 41A View FIGURE 41 ). The second valvifer is longer than the first valvifer and divided into the anterior area of the second valvifer (2vfaa: Figs 41B, 41F View FIGURE 41 ), and posterior area of the second valvifer (2vfpa: Fig. 41B View FIGURE 41 ). The posterior area of the second valvifer has two more sclerotized areas, the posterior section of the dorsal flange of the second valvifer (2vfps: Fig. 41C View FIGURE 41 ) and the anterior section of the dorsal flange of the second valvifer (2vfas: Fig. 41C View FIGURE 41 ). The posterior end of the first valvular ramus is limited by the anterior angle of the first valvifer (1vfan: Fig. 41B View FIGURE 41 ), which precedes the dorsal projection of the second valvifer (2vfdp: Fig. 41B View FIGURE 41 ).









No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.