
Smeringopina fang, Huber, Bernhard A., 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3713.1.1 |
|
publication LSID |
lsid:zoobank.org:pub:C5F0BC11-92C0-4B30-9DB3-200882AC8950 |
|
DOI |
https://doi.org/10.5281/zenodo.6161979 |
|
persistent identifier |
https://treatment.plazi.org/id/B20287ED-FFEB-FF8A-B990-C3EEFCE13835 |
|
treatment provided by |
Plazi |
|
scientific name |
Smeringopina fang |
| status |
sp. nov. |
Smeringopina fang View in CoL new species
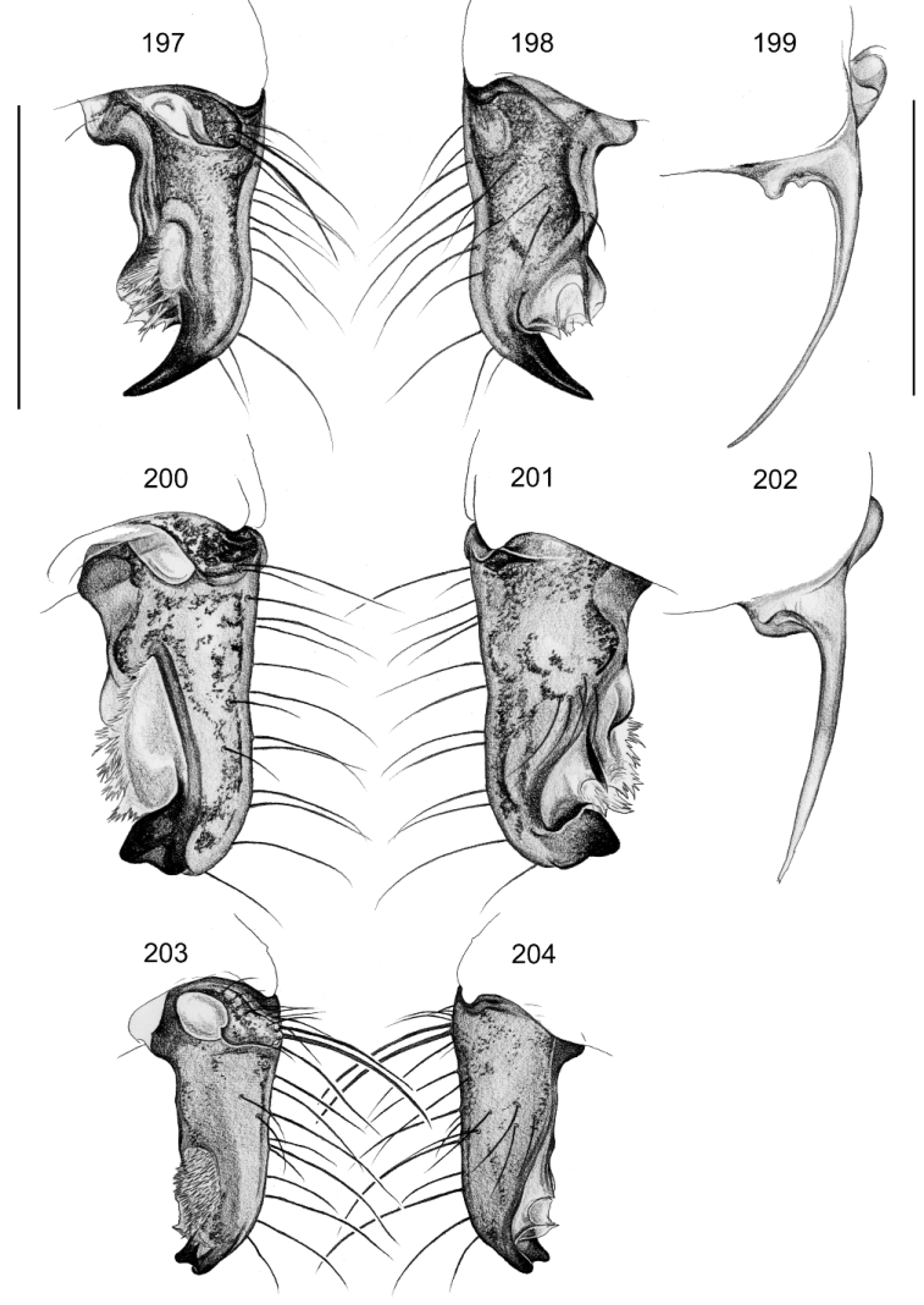
Figs. 29–30 View FIGURES 17 – 31 , 125–132 View FIGURES 125 – 132 , 165–166, 175–176 View FIGURES 163 – 183 , 197–224 View FIGURES 197 – 204 View FIGURES 205 – 210 View FIGURES 211 – 224
Type. ♂ holotype from Gabon, Ogooué-Ivindo, near Djidji (0°12.8’N, 11°49.3’E), 470 m a.s.l., forest, 12.viii.2011 (B.A. & S.R. Huber), in ZFMK (Ar 10193).
Other material examined. GABON: Ogooué-Ivindo: near Djidji , same data as holotype, 6♂ 7♀ 1 juv. (2 vials) in ZFMK (Ar 10194-95); same data, 1♂ 1♀ 2 juvs. in pure ethanol, in ZFMK (Gab 239). Monts de Belinga , forest near Mayebout (1°06.7’N, 13°06.6’E), 500 m a.s.l., 13.–14.viii.2011 (B.A. & S.R. Huber), 3♂ 14♀ 1 juv. in ZFMK (Ar 10196); same data, 5♀ 1 juv. in pure ethanol, in ZFMK (Gab 197). Near Ntenkélé (0°31.4’N, 12°31.5’E), 550 m a.s.l., forest, 12.viii.2011 (B.A. & S.R. Huber), 2♀ in ZFMK (Ar 10197); same data, 1♀ 1 juv. in pure ethanol, in ZFMK (Gab 236). Makokou, CNRS [0°30.8’N, 12°48.2’E; not 0°04’N, 12°08’E as on label], ix.–xi.1976 (A.L. Rypstra), 8♂ 18♀ 4 juvs. in USNM. Moyen-Ogooué: near Ndjolé , “site 1” (0°09.0’S, 10°40.0’E), 120 m a.s.l., forest near brook, 11.viii.2011 (B.A. & S.R. Huber), 6♂ 3♀ 2 juvs. in ZFMK (Ar 10198); same data, 2♀ 1 juv. in pure ethanol, in ZFMK (Gab 230). Ngounié: Massif du Chaillu , “site 2”, near Moukabou (1°36.6’S, 11°40.7’E), 560 m a.s.l., forest, 25.viii.2011 (B.A. & S.R. Huber), 3♂ 2♀ in ZFMK (Ar 10199); same data, 1♀ 1 juv. in pure ethanol, in ZFMK (Gab 175). Massif du Chaillu , “site 3”, between Mimongo & Yéno (1°38.1’S, 11°32.6’E), 570– 650 m a.s.l., forest, 26.viii.2011 (B.A. & S.R. Huber), 2♂ 6♀ in ZFMK (Ar 10200); same data, 1 juv. in pure ethanol, in ZFMK (Gab 235).
Etymology. Named for the Fang , an ethnic group ranging from southern Cameroon south into Gabon and Congo; noun in apposition.
Diagnosis. Easily distinguished from known congeners by distinctive long and slender process of bulb ( Fig. 199 View FIGURES 197 – 204 ) and by highly distinctive pair of processes on epigynum ( Figs. 206 View FIGURES 205 – 210 , 221 View FIGURES 211 – 224 ); also by pair of large sclerotized areas medially on male chelicerae ( Fig. 205 View FIGURES 205 – 210 ; sometimes indistinct) and small retrolateral apophysis on male palpal femur directed toward dorsally ( Fig. 129 View FIGURES 125 – 132 ).
Male (holotype). Total body length 3.2, carapace width 1.1. Leg 1: 34.3 (8.1 + 0.4 + 8.3 + 15.8 + 1.7), tibia 2: 4.8, tibia 3: 3.3, tibia 4: 5.1; tibia 1 L/d: 94. Distance PME-PME 95 µm, diameter PME 115 µm, distance PME- ALE 55 µm, distance AME-AME 25 µm, diameter AME 90 µm. Carapace ochre-yellow with brown triangular mark posteriorly and brown lateral margins; ocular area with brown mark posteriorly; clypeus with pair of brown bands, sternum brown; legs light brown, with indistinct darker rings subdistally on femora and tibiae and in patella area, tips of femora and tibiae whitish; abdomen ochre-gray with dark pattern dorsally, laterally, and ventrally, ventral dark bands with lateral constriction. Habitus as in Figs. 125–126 View FIGURES 125 – 132 , ocular area slightly elevated, secondary eyes with indistinct ‘pseudo-lenses’; clypeus with pointed and slightly hooked apophysis near rim ( Figs. 211, 213– 214 View FIGURES 211 – 224 ); deep thoracic pit and pair of shallow furrows diverging behind pit. Chelicerae as in Figs. 205 View FIGURES 205 – 210 and 213–215 View FIGURES 211 – 224 , with lateral apophyses in very distal position, with distinctive pair of large sclerotized areas medially, with rows of small frontal apophyses, without modified hairs. Palps as in Figs. 127–129 View FIGURES 125 – 132 ; coxa with indistinct retrolateral apophysis; trochanter with simple ventral apophysis; femur with large retrolateral apophysis directed toward ventrally, small retrolateral apophysis directed toward dorsally, small proximal prolateral process, and weakly sclerotized ventral projection distally; prolateral femur-patella joint strongly shifted toward ventrally; tarsus with some longer and slightly stronger hairs dorsally ( Fig. 216 View FIGURES 211 – 224 ); procursus with large distal apophysis and complex membranous prolatero-ventral structures, without hinge ( Figs. 197–198 View FIGURES 197 – 204 ); bulb with long slender process (sperm duct opens at basis of this process from weakly sclerotized bulge; Figs. 199 View FIGURES 197 – 204 , 218 View FIGURES 211 – 224 ). Legs without spines and curved hairs, with few vertical hairs; retrolateral trichobothrium on tibia 1 at 1%; prolateral trichobothrium present on all tibiae; pseudosegments barely visible. ALS with eight spigots each; male gonopore with two epiandrous spigots ( Fig. 219 View FIGURES 211 – 224 ).
Variation. There is conspicuous variation among localities and the species may eventually turn out to include several reproductively isolated communities. They are here treated as one species because different characters suggest different groupings and some characters seem to vary continuously and are difficult to evaluate given the small sample of localities and specimens. The distal procursus apophysis may be short and thick ( Ndjolé ; Figs. 200–201 View FIGURES 197 – 204 ), short and bifid (Massif du Chaillu ; Figs. 203–204 View FIGURES 197 – 204 ), or just much shorter than in the holotype ( Mayebout , Makokou); the membranous structures prolatero-ventrally on the procursus seem to differ among localities but are difficult to evaluate; the same is true for the slender process of the bulb that may be variably curved ( Figs. 199, 202 View FIGURES 197 – 204 ). Tibia 1 in 23 other males: 7.7–9.2 (mean 8.3).
Female. In general similar to male; clypeus unmodified. Tibia 1 in 44 females: 4.8–5.8 (mean 5.2). Epigynum anterior plate with pair of distinctive projections ( Figs. 206 View FIGURES 205 – 210 , 221 View FIGURES 211 – 224 ) whose shapes and positions vary among localities (more rounded than in specimens from type locality in specimens from Massif du Chaillu , Ntenkélé, Ndjolé : Fig. 209 View FIGURES 205 – 210 ; closer together in females from Ntenkélé); entire anterior plate sometimes relatively narrower ( Mayebout , Makokou, Ntenkélé); posterior indentation sometimes much deeper (Massif du Chaillu , Makokou, Ndjolé : Fig. 209 View FIGURES 205 – 210 , Mayebout : Figs. 222, 224 View FIGURES 211 – 224 ); with rugose area in anterior part of anterior epigynal plate; posterior plate laterally with overhanging folds; internal genitalia as in Figs. 175–176 View FIGURES 163 – 183 , 207, 210 View FIGURES 205 – 210 .
Natural history. Litter-dwelling species with small domed webs under dead curved leaves on the ground. At “site 1” near Ndjolé this species was found to share the leaf-litter with the superficially very similar S. ndjole .
Distribution. Widely distributed in Gabon ( Fig. 114 View FIGURE 114 ).






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.