
Burmalindenia imPerfecta
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4103.6.4 |
|
publication LSID |
lsid:zoobank.org:pub:09A664E2-4322-4ED9-B146-61EB64B7924C |
|
DOI |
https://doi.org/10.5281/zenodo.6079165 |
|
persistent identifier |
https://treatment.plazi.org/id/B2037513-FFD3-FF87-258B-7B45FEE5FA9F |
|
treatment provided by |
Plazi |
|
scientific name |
Burmalindenia imPerfecta |
| status |
|
Burmalindenia imPerfecta n. sp.
Figures 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 , 8 View FIGURE 8
Holotype. SMNS Bu-94, State Museum of Natural History Stuttgart, Germany.
Diagnosis. This species can be discriminated from all known Recent and fossil odonates by the following characters in the hindwing venation: bracket-like Ax1 and Ax2 with 3 secondary antenodals between them; straight arculus; basal radial area free; 7 or more antefurcal crossveins between RP and MA; distinctly curved hypertriangle with two cells; discoidal triangle elongated and with two cells; ddv/MAb not strongly sigmoid but with slight angle; distinct Tspl in the postdiscoidal space with 2 rows of cells; strong PsA (plesiomorphic compared to Recent Lindeniinae and Hageniinae); subdiscoidal triangle free; anal vein secondary branched; no anal loop; gaff not prolonged.
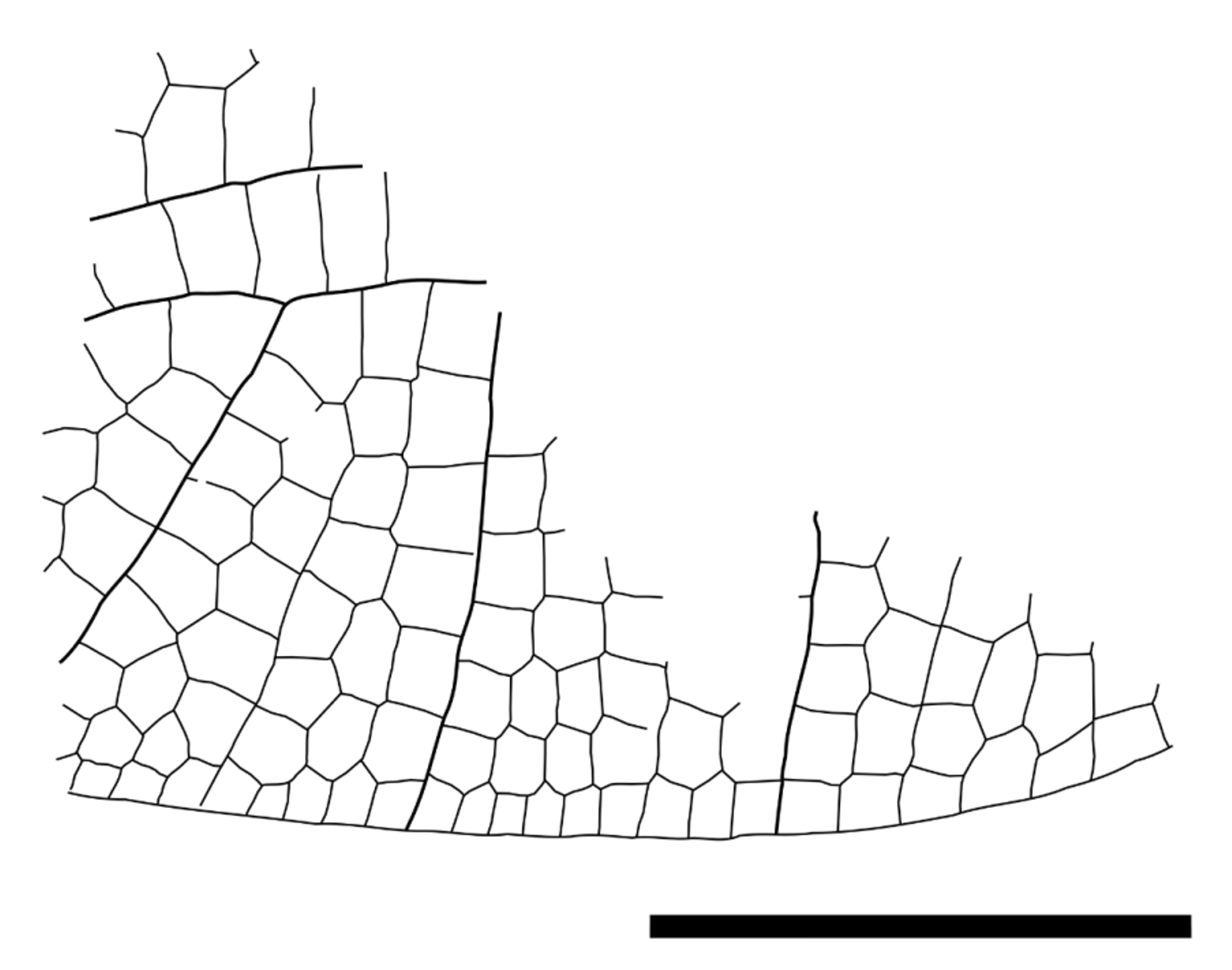
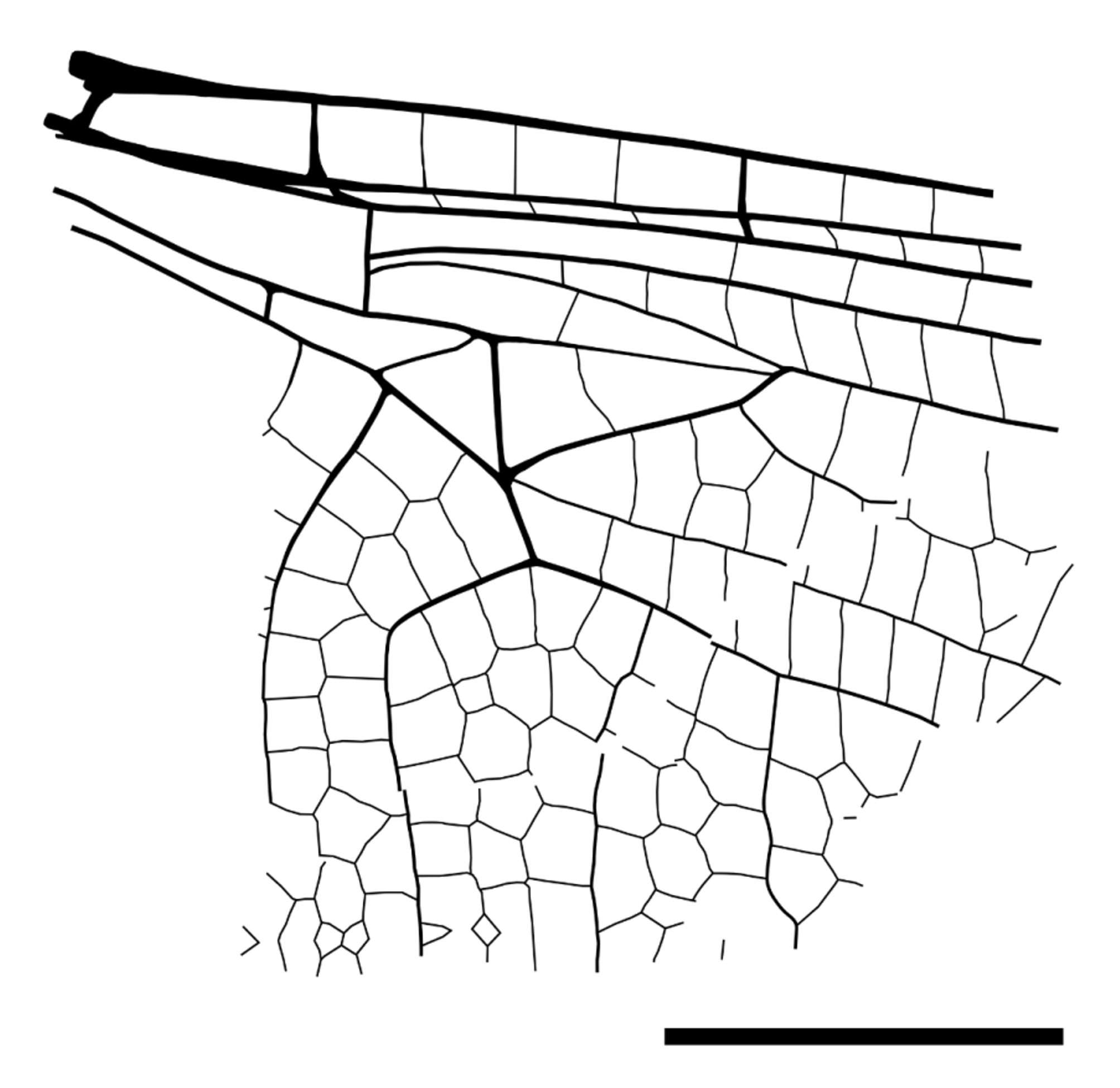
Description. The fossil-bearing piece of amber (SMNS Bu-94) is more or less oval shaped and measures 30.9 mm in length and 16.3 mm in width. It includes two completely hyaline fragments of the left and right hindwing of a dragonfly.
Right wing fragment. The fragment of the right hindwing mainly consists of the cubital region. Posterior wing margin intact at full length of the fragment. Cubital region anteriorly completed with CuA’. In distal-proximal direction no such completion could be determined. In most anterior part of fragment MP and few cells (at least two rows) of postdiscoidal field preserved. Total dimensions 13.8 mm in horizontal direction and 9.0 mm in vertical direction (Costa as reference—estimated on the basis of the CuA’-Costa angle).
Two vertical branches of CuA’ determined. A third vertical strong vein more basal could be either a CuA’ branch or the CuA’’ depending on the position of the fragment within the wing, but the visible terminal fork of CuA’ rather suggests that the third vein also is a vertical branch of CuA’, because CuA’ would otherwise be unusually short. There are 7 rows of cells in the antecubital field.
MP and the CuA’ run almost parallel, and only a single row of cells between them.
Left wing fragment. The fragment of the left hindwing consists of the proximal region of the wing. Anterior margin of wing is completed by Costa at full length of fragment. In distal direction the fragment ranges to the third antenodal region, in proximal direction to the ScA’’ (= Ax0). The fragment lacks a posterior margin of the wing, although large parts of the cubital and anal region are preserved. Total dimensions 13.7 mm in horizontal direction and 10.1 mm in vertical direction (Costa as reference for direction).
First and the second antenodal crossveins (Ax1 and Ax2) aligned and stronger than others (bracket-like); three more or less aligned secondary antenodal crossveins between Ax1 and Ax2; distal of Ax2 there are two secondary antenodals of the first row, and three of the second row preserved and not aligned.
Arculus close to the Ax1 and straight. Bases of RA and MA on arculus and separated. Hypertriangle 5.3 mm long and maximum 0.8 mm wide; costal side (MA) distinctly curved; separated by one secondary crossvein at about half length. Basal space free.
Discoidal triangle longitudinally stretched with a slight kink (origin of trigonal supplement Tspl) in posterodistal side (MAb). Divided by an oblique secondary crossvein at one third length of anterior side. Basal side of discoidal triangle 1.7 mm long; costal side 3.6 mm; posterodistal side ddv/MAb 3.6 mm. Two rows of cells in basal postdiscoidal space.
CuP crossing 0.4 mm long and 1.2 mm proximal of arculus. Quadrangular subdiscoidal cell divided by a distinct and strong pseudo-anal vein (PsA=AA0) creating an almost equilateral subdiscoidal triangle. Subdiscoidal triangle free. Basal part of cubital cell free as well.
Up to three rows of cells between AA1b and CuA’’; no anal loop; basally of AA1b there is one row of small cells preserved which is basally completed by a thin and errant vein suggesting that there were at least two rows of small cells located basally of AA1b.
Gaff (CuA from posterior angle of discoidal triangle to CuA’/CuA’’-branching) 1.1 mm long (not prolonged). CuA branches at an angle of 142° into CuA’ and CuA’’. Two posterior branches of CuA’ are preserved.
One row of cells between CuA’ and MP at a width of 1.1 mm. Postdiscoidal area (basally 2.4 mm wide) with two rows of cells, divided by trigonal supplement tspl. Basal arcular area with one row of cells. Basal radial area free almost up to level of Ax2.
Type locality and horizon. Burmese amber (Hukawng Valley, Kachin State, Myanmar). Mid-Cretaceous: earliest Cenomanian.
Etymology. The species name imperfecta refers to the incomplete preservation of the holotype as well as its unique teratological wing venation.
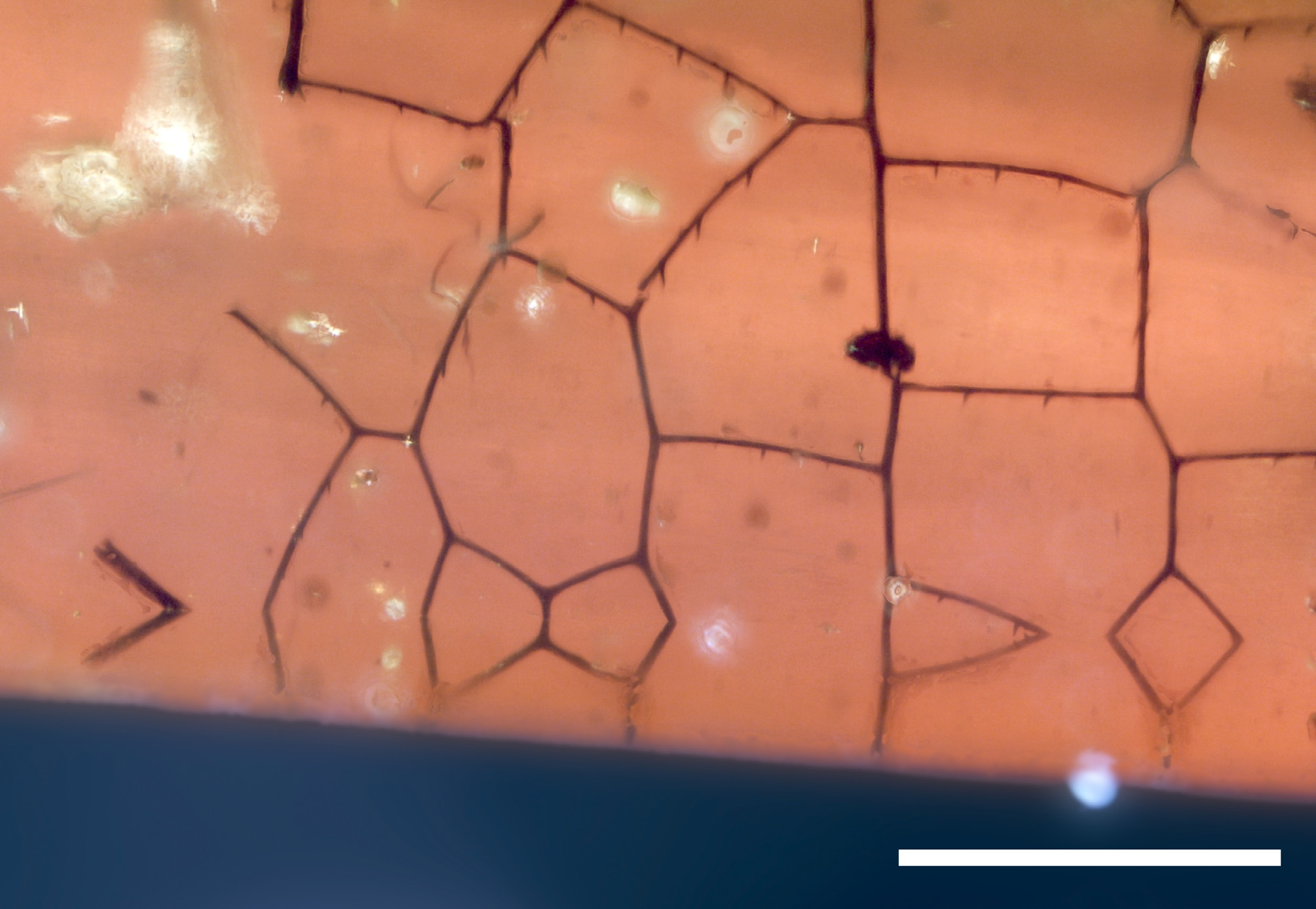
Syninclusions. Apart from the dragonfly remains the amber piece contains several interesting botanical and hexapodan syninclusions ( Figure 7 View FIGURE 7 ), of which the oldest fossil record for the mayfly subfamily Cloeoninae ( Ephemeroptera : Baetidae ) is the most interesting (Hodunko, in prep.).
Stellate hairs. Plants produce trichomes for various reasons. They can help to reduce air fluctuation on the plant surfaces in order to regulate evaporation or protect leaves and buds from excessive sunlight and herbivores ( Levin 1973, Werker 2000). Stellate hairs are a special morphotype of plant trichomes that occur in various amber sites.
Stellate hairs are characteristic of the Eocene Baltic Amber, where they have been determined as a product of oak trees ( Fagaceae ) ( Weitschat et al. 2010). They also occur in Cretaceous ambers from Spain ( Penalver et al. 2007), Lebanon ( Koteja and Azar 2008), and Ethiopia ( Schmidt et al. 2010). In the Cenomanian amber site in Ethiopia tree ferns and epiphytic ferns were determined as the source of the stellate trichomes ( Schmidt et al. 2010). Ferns and conifers were suggested for the Early Cretaceous amber from Spain ( Penalver et al. 2007). These two attempts of determination are based on comparison with Recent plant trichomes.
Stellate hairs have also already been mentioned as syninclusions in Burmese Amber ( Koteja 2004). In the studied amber piece there are numerous stellate hairs embedded as syninclusions, but their botanical origin is still undetermined.




| SMNS |
Staatliches Museum fuer Naturkund Stuttgart |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.