
Dibamus deimontis, Kliukin & Bragin & Nguyen & Le & Tran & Gorin & Poyarkov, 2024
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5406.1.4 |
|
publication LSID |
lsid:zoobank.org:pub:F2775F85-B2B8-49AF-8802-7A2C6FFD7197 |
|
DOI |
https://doi.org/10.5281/zenodo.10624645 |
|
persistent identifier |
https://treatment.plazi.org/id/B22B87C1-CF16-3859-76D8-3F99FE12FF09 |
|
treatment provided by |
Plazi |
|
scientific name |
Dibamus deimontis |
| status |
sp. nov. |
Dibamus deimontis sp. nov. Kliukin, Bragin, Nguyen & Poyarkov
( Figs. 1–5 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 ; Tables 1 View TABLE 1 , 2 View TABLE 2 )
Holotype. Adult male ( ZMMU Re-17821; field tag NAP-13707 ) collected from eastern slope of Nui Chua Mountain within Nui Chua National Park , Thai An Village , Vinh Hai Commune, Ninh Hai District, Ninh Thuan Province, southern Vietnam (11.723°N, 109.137°E; elevation 695 m a.s.l.) on March 3, 2023, at 10:00 h by A. M. Bragin, N. A. Poyarkov, and S. X. Le. GoogleMaps
Paratypes (n=7). ZMMU Re-17822 (field tag NAP-13640 ) , ZMMU Re-17824 (field tag NAP-13706 ) and VRTC NAP-13708 (field tag NAP-13708 ) , three adult males collected at the same locality as the holotype on March 3–5, 2023 from the elevations 670–700 m a.s.l. GoogleMaps ; ZMMU Re-17823 (field tag NAP-13641 ) , ZMMU Re-17825 (field tag NAP-13709 ), and ZMMU Re-17826 (field tag NAP-13710 ) , ZMMU Re-17827 (field tag NAP-13711 ), two adult females and two subadult females, respectively, with the same collection information as the holotype and other paratypes .
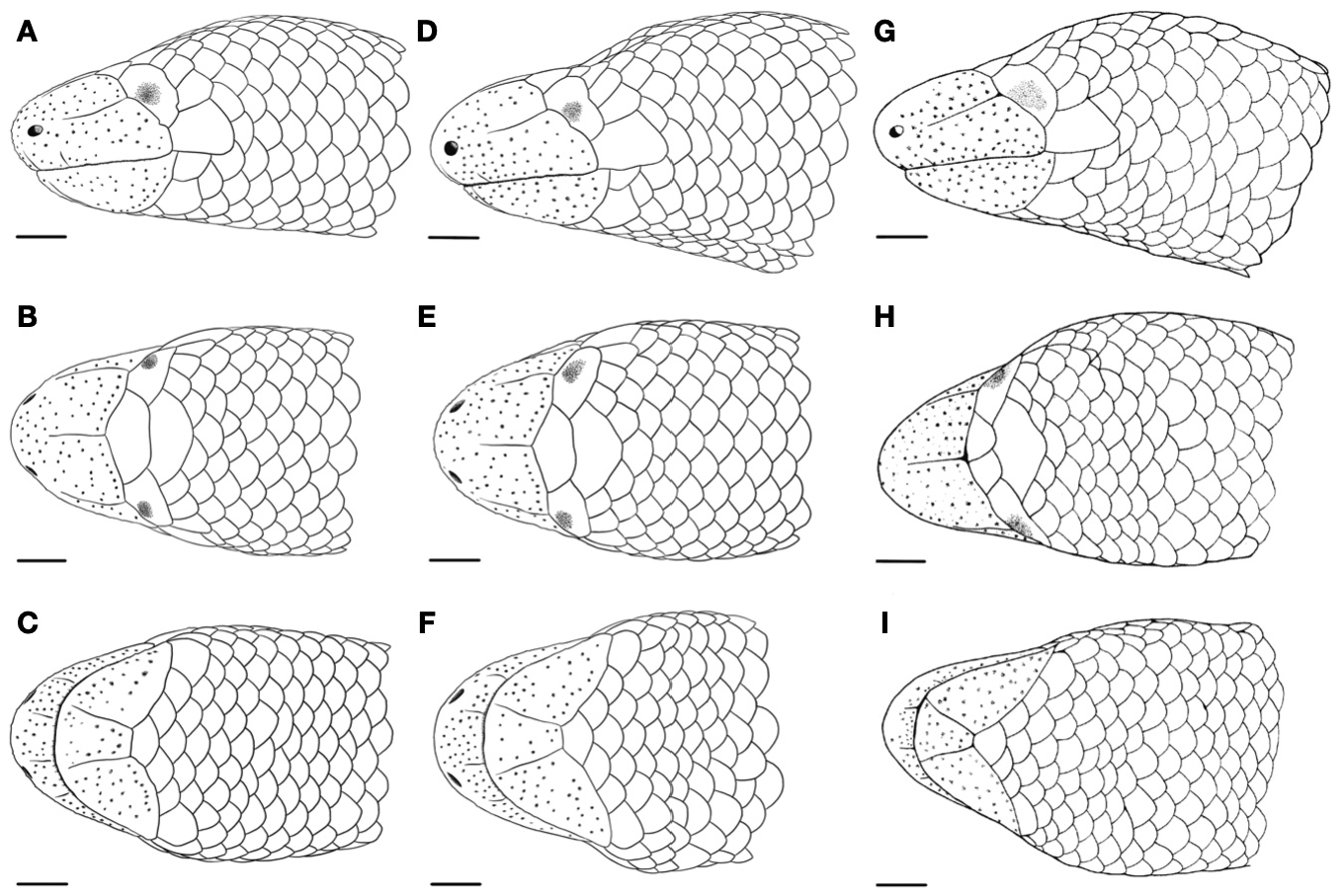
Diagnosis. The new species is assigned to Dibamus based on the following morphological characters: (1) worm-like body shape, fore limbs absent, hind limbs rudimentary, present only in males, forming flap-like structures located near the base of the tail above the vent; (2) eyes rudimentary, completely covered by scales; (3) external ear openings absent; (4) enlarged plate-like scales on the head; (5) postorbital bone absent; (6) epipterygoid absent; (7) temporal bone absent; (8) ribs on last presacral vertebra absent; (9) tail length less than 34.0% of SVL; (10) subcaudal scales in females reduced to 64; and (11) postsacral vertebrae reduced to 36 in a single male ( Greer 1985). The new species Dibamus deimontis sp. nov. can be distinguished from all other congeners by the following combination of morphological characters: (1) maximum SVL of 136.2 mm; (2) tail comparatively long, TL comprising 18.6–22.1% of SVL in males, 17.6–20.2% of SVL in females; (3) labial, nasal, medial and lateral rostral sutures present but incomplete; (4) postocular scales generally three, rarely two; (5) three to five scales bordering the posteromedial edge of the first infralabial; (6) the medial sublabial scale not enlarged; (7) 22–25 midbody scale rows; (8) 25–30 transverse scale rows just posterior to head; (9) 19–21 transverse scale rows just anterior to vent; (10) 193–225 ventral scales; (11) 47–55 subcaudal scales; (12) relative size of frontal to frontonasal 106.7–133.8%; (13) relative size of interparietal to surrounding scales 97.2–115.0%; and (14) the light colored band on the body generally present.
Description of holotype ( Fig. 2 A, B, D View FIGURE 2 ; Fig. 3A–C View FIGURE 3 ; Fig. 4A–C View FIGURE 4 ). An adult male in a good state of preservation; SVL 128 mm; tail length 23.8 mm (18.6% of SVL); head longer (HL 3.98 mm) than wide (HW 3.44 mm); snout bluntly rounded, projecting beyond the jaw ( Fig. 3A View FIGURE 3 , Fig. 4A View FIGURE 4 ; E-S 2.42 mm; E-N 1.65 mm; IN 1.05 mm; OI 2.13 mm); labial, nasal, and rostral sutures present, but incomplete ( Fig. 3A–C View FIGURE 3 ; Fig. 4A–C View FIGURE 4 ); rostral pad with a large number of evenly distributed sensory papillae; three postocular scales on each side; ear opening absent; eyes barely visible below the single ocular scale; supralabial single, slightly larger than ocular scale, bordering ocular ventrally; three scales posterior to supralabial ( Fig. 3A View FIGURE 3 , Fig. 4A View FIGURE 4 ); frontal scale slightly larger than frontonasal scale (FSW/FNSW 126%); frontonasal scale wider than long (FNSL/FNSW 52.1%); interparietal single, not enlarged, narrower than frontonasal and frontal, posteriorly bordered by four slightly smaller nuchal scales ( Fig. 3B View FIGURE 3 , Fig. 4B View FIGURE 4 ); infralabials lanceolate, separated by a smaller mental; mental narrow, trapezoid in shape, bordered by the first infralabial on each side; three scales contacting the first infralabial, a small medial scale posterior the mental, and three larger scales contacting the infralabial posteromedially ( Fig. 3C View FIGURE 3 , Fig. 4C View FIGURE 4 ). Body wormlike, almost cylindrical ( Fig. 2A,B View FIGURE 2 ); body scales smooth, subcycloid ( Figs. 3–4 View FIGURE 3 View FIGURE 4 ); 22 midbody scale rows; 27 scale rows just posterior to head; 21 scale rows anterior to vent; scales near vent thick; 51 subcaudals; 199 ventrals; tail complete, rudimentary flap-like hind limbs present ( Fig. 2D View FIGURE 2 ; HLL 4.54 mm); tail tip blunt, covered by a single rounded scale, not terminating in a spine. Morphometric data of the holotype are presented in Table 1 View TABLE 1 .
Coloration. In life, dorsum, flanks and tail light dark-brown and grey-brown to pinkish-brown ( Fig. 2A–B View FIGURE 2 ); ventral surface, snout and head slightly paler; rostral and mental pad lighter, cream coloured; anal region and tip of legs pinkish-cream ( Fig. 2D View FIGURE 2 ). Distinct pale grey transverse body band at tail base complete (ca. six or seven scales in width); incomplete pale grey band on neck (ca. five or six scales in width), large irregular grey spots on dorsum and a large light-grey spot anterior to vent. In preservative after ten months of storage in ethanol the general coloration pattern does not change, though pinkish and brownish fade to dull-white or beige.
Variation. The measurements and counts of the type series revealed high variability in external morphological characters ( Table 1 View TABLE 1 ); variation of head scalation of male holotype and paratypes is presented in Figs. 3–4 View FIGURE 3 View FIGURE 4 . The female paratypes are generally similar to the holotype in morphology with the major difference being the absence of the two short flap-like hind limbs; in females cloacal opening represents small transverse slit ( Fig. 2C View FIGURE 2 ). This difference clearly reflects sexual dimorphism previously reported for Dibamus ( Greer 1985; Neang et al. 2011; Quah et al. 2017; Koppetsch et al. 2019; Kliukin et al. 2023). Significant variation was observed in the number of postocular scales: six members of the type series had three postocular scales (PO), but one male paratype (ZMMU Re-17822) and one female paratype (ZMMU Re-17823) had only two postocular scales ( Table 1 View TABLE 1 ). Moreover, three male paratypes (ZMMU Re-17822, Re-17824; and VRTC NAP-13708) and three female paratypes (ZMMU Re-17823, Re-17825, and Re-17826) exhibit a slightly higher number of midbody scale rows (MBSR 23–25), while the holotype ZMMU Re-17821 and female paratype ZMMU Re-17827 have only 22 midbody scale rows ( Table 1 View TABLE 1 ). One male paratype VRTC NAP-13708 and one female paratype ZMMU Re-17826 have higher number of scale rows posterior to head in comparison with the holotype (PHSR 28–30 vs. 27), while other paratypes (ZMMU Re-17822, Re-17823, and Re-17827) have lower number of scale rows posterior to head (PHSR 25–26 vs. 27). One male paratype VRTC NAP-13708 has lower number of scale rows anterior to vent (VSR 19 vs. 21) ( Table 1 View TABLE 1 ). In a single male specimen ZMMU Re-17822 tail is damaged and regenerated what can be noticed by a different structure of the scales at the tail tip (as described in Darevsky 1992). This specimen has comparatively shorter tail length and a notably lower number of subcaudal scales (SC 33 vs. 47–55 in other type specimens; Table 1 View TABLE 1 ); therefore these values were not used in morphological comparisons of the new species. It seems that the presence and degree of development of the light transverse band is highly variable in the new species: it was completely absent in a subadult female paratype ZMMU Re-17827; incomplete bands reduced to large irregular dull-white spots were present on body in subadult female paratype ZMMU Re-17826 and adult female paratype ZMMU Re-17825. At the same time, all other members of type series had at least one complete band on body (VRTC NAP-13708, ZMMU Re-17822), while one male paratype had two complete bands on body (ZMMU Re-17824), while one male paratype (ZMMU Re-17823) had two additional incomplete bands at body and tail. Finally, variation was observed in background coloration, which ranged from light brown to dark gray in life ( Fig. 2 View FIGURE 2 ). Coloration pattern and condition of light band might depend on age, sex or physiological condition of a given specimen; it appears that the use of these characters in Dibamus taxonomy is not advisable.
Hemipenial morphology. Male paratype specimen VRTC NAP-13708 had two partially everted hemipenes, which represent quite smooth conic structures located medially from each rudimentary hindlimb ( Fig. 2E View FIGURE 2 ). Each hemipenis is tapering to the apex with a small hollow near its tip. Everted organ was ca. 1.0 mm in length with ca. 0.5 mm in width. Our observations match well with the description of hemipenial structures of D. greeri presented by Darevsky (1992) .
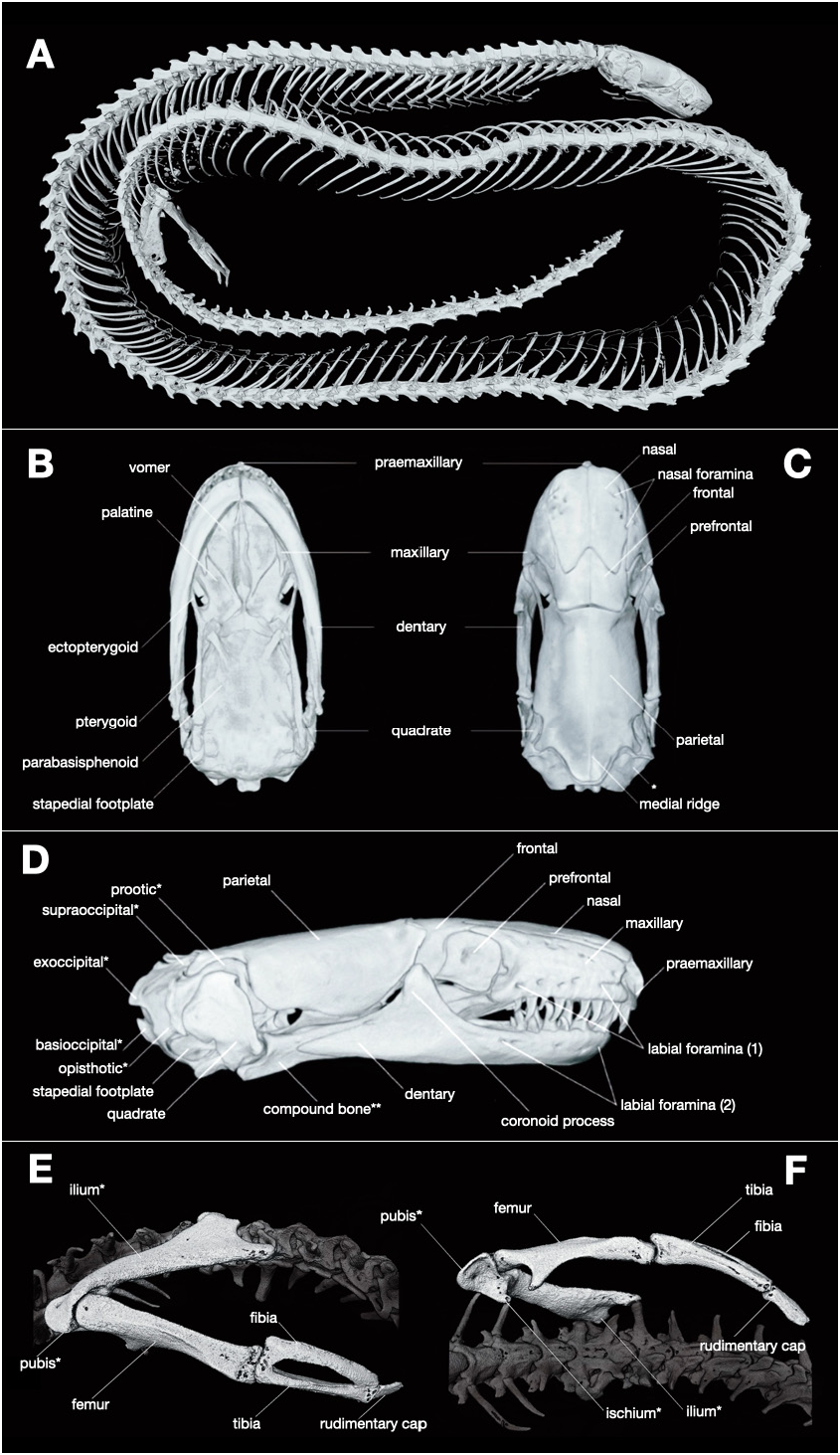
Osteological description. The following description of skeletal features of the new species is based on the microCT data obtained from the male paratype specimen ZMMU Re-17824. Skeletal morphology of this specimen is presented in Fig. 5 View FIGURE 5 ; osteological terminology generally follows Greer (1985), Rieppel (1984), and Kliukin et al. (2023). Skull small relative to the body, elongated; shoulder girdle absent; pelvic girdle and rudiments of hind limbs present ( Fig. 5A View FIGURE 5 ). Body with 115 presacral vertebrae, 2 fused sacral vertebrae, and 27 tail vertebrae ( Fig. 5A View FIGURE 5 ). The pelvic girdle includes fused ilium, ischium, and pubis; hind limbs include femur, tibia, fibula, and a rudimentary bony cap ( Fig. 5E–F View FIGURE 5 ).
The premaxilla is an unpaired bone ( Fig. 5B–D View FIGURE 5 ); the transverse process of premaxilla pierced by paired apical foramina. Seven labial foramina present on the left maxilla, six labial foramina present on the right maxilla. Each of the maxillary bones bearing eight slightly curved teeth ( Fig. 5D View FIGURE 5 ). The nasal bones distinct, perforated by five or six foramina on each side ( Fig. 5C View FIGURE 5 ). The prefrontal represents a rather rectangular bone forming the posterodorsal edge of the lacrimal foramen. The lacrimal, postfrontal, postorbital and jugal bones absent. The paired frontals form the anteromedial processes dividing the posterior portions of the nasals ( Fig. 5C View FIGURE 5 ), anterolateral processes of frontals well-developed, protruding anteriorly further than anteromedial processes. The unpaired parietal bone forms supratemporal processes which overlay the dorsal portions of the prootics and a wide posterior medial process which meets the supraoccipital in a suture ( Fig. 5C View FIGURE 5 ). The sagittal ridge on the parietal present; notably developed on supratemporal process ( Fig. 5C View FIGURE 5 ). The vomers, palatine, pterygoids, and ectopterygoids are paired bones ( Fig. 5B View FIGURE 5 ) with morphology similar to that described for the other members of the genus Dibamus by Greer (1985).
The parabasisphenoid complex (corresponding to the ‘parasphenoid + basisphenoid’ of Rieppel 1984) forms a broad massive bone meeting the basioccipital without a distinct suture, its lateral boarders rather straight. Exoccipitals, basioccipitals, supraoccipitals, prootics and opisthotics fused ( Fig. 5B–D View FIGURE 5 ). The stapes is characterized by a broad stapedial footplate slightly elongated anteroposteriorly ( Fig. 5D View FIGURE 5 ). The quadrate bone short and flattened; its vertical axis is slightly tilted posteriorly.
Five labial foramina present on the each side of the dentary ( Fig. 5D View FIGURE 5 ). The dentary bears seven teeth on the left and eight teeth on the right side. Coronoid process of the dentary comparatively high; a distinct coronoid bone absent. Compound bone is formed by the fused splenial, articular, angular and supraangular bones; the retroarticular process of the compound bone comparatively short ( Fig. 5D View FIGURE 5 ). The lower jaw with mandibular, anterior and posterior suprangular foramina.
...Coninued on the next page
Etymology. The epithet of the new species is a Latin toponymic adjective in genitive singular, and is given in reference to its type locality on Nui Chua Mountain, Ninh Thuan Province, Vietnam. As Nui Chua Mt. is the highest peak in the area, in Vietnamese “Núi Chúa” literally means “the Mountain of God”, or “the Mountain of Lord”, which can be translated to Latin as “ deimontis ”. We recommend the names “ Nui Chua Blind Skink ”, “ Nuichuyskaya cherveobraznaya yascheritsa ”, and “ Thằn lằn giun Núi Chúa ” as common names of the new species in English, Russian, and Vietnamese, respectively.
Comparisons. Comparative morphological data for the new species and the currently recognized 25 nominal species of the genus Dibamus is presented in Table 2 View TABLE 2 . Dibamus deimontis sp. nov. is steadily differentiated from other congeners by having generally three postoculars, with only three other Dibamus species sharing this character states namely: D. celebensis Schlegel , D. novaeguineae Duméril & Bibron , and D. taylori Greer. All these three species occur on Southeast Asian Islands in Indonesia and Philippines; no species of Dibamus from the mainland Asia was reported to have three postocular scales. The new species can be further distinguished from Dibamus celebensis by having incomplete labial and nasal suture (vs. complete), by the presence of medial rostral suture (vs. absence), by a lower number of midbody scale rows (MBSR 22–25 vs. 26–30), by a higher number of subcaudal scales (SC 47–55 vs. 38–40), and by longer tail (TL/SVL 14.5–24.3% vs. 10–13%). Dibamus deimontis sp. nov. is further diagnosed from D. novaeguineae by having incomplete labial and nasal suture (vs. complete), by the presence of medial rostral suture (vs. absence), by a higher number of subcaudal scales (SC 47–55 vs. 38–40) and by longer tail (TL/SVL 14.5–24.3% vs. 10–19%). The new species can be further distinguished from Dibamus taylori by incomplete labial and nasal suture (vs. complete), by presence of medial rostral suture (vs. absence), by generally higher number of scales on the posteromedial edge of infralabial (PIS 3–5 vs. 2–4), and by generally longer tail (TL/SVL 14.5–24.3% vs. 14–19%).
Regarding the remaining species of Dibamu s, the absence of lateral rostral sutures distinguishes the new species from D. bogadeki Darevsky and D. bourreti , the only two species of Dibamus in which these sutures are present. By the presence of incomplete (rudimental) medial rostral suture Dibamus deimontis sp. nov. can be differentiated from species which completely lack rostral suture, namely: D. alfredi Taylor , D. bogadeki , D. booliati Das & Yaakob , D. bourreti , D. leucurus (Bleeker) , D. seramensis Greer , and D. smithi , and also from the two species which have a complete rostral suture, namely: D. montanus and D. somsaki Honda, Nabhitabhata, Ota & Hikida. The presence of an incomplete (rudimental) nasal suture further differs Dibamus deimontis sp. nov. from the species in which this suture is complete, including: D. bogadeki , D. bourreti , D. dalaiensis Neang, Holden, Eastoe, Seng, Ith & Grismer , D. deharvengi , D. dezwaani Das & Lim , D. floweri Quah, Anuar, Grismer & Grassby-Lewis , D. ingeri Das & Lim , D. kondaoensis , D. leucurus , D. manadotuaensis Koppetsch, Böhme & Koch , D. montanus , D. nicobaricus (Steindachner) , D. seramensis , D. somsaki , D. tebal Das & Lim , and from D. tiomanensis Diaz, Leong, Grismer & Yaakob in which this suture is completely absent. By having the incomplete (rudimental) labial suture Dibamus deimontis sp. nov. can be differentiated from those species which have complete labial suture, namely: D. bogadeki , D. bourreti , D. dalaiensis , D. dezwaani , D. ingeri , D. kondaoensis , D. manadotuaensis , D. montanus , D. nicobaricus , D. novaeguineae Duméril & Bibron , D. seramensis , D. somsaki , D. taylori Greer , D. tebal , and D. tiomanensis , as well as from two species in which this suture is completely absent, namely: D. deharvengi and D. floweri .
When compared to D. tropcentr , another congener inhabiting the Nui Chua N.P., Dibamus deimontis sp. nov. can be easily differentiated by having: generally three (rarely two) postoculars (vs. always two postoculars), by higher number of midbody scale rows (MBSR 22–25 vs. 19–21), by higher number of scale rows posterior to head (PHSR 25–30 vs. 23–24), by higher number of scale rows anterior to vent (VSR 19–21 vs. 23–24), by lower number of subcaudal scales (SC 47–55 vs. 64–65), and by notably shorter tail (TL/SVL 14.5–24.3% vs. 24.4–30). In general appearance, Dibamus deimontis sp. nov. is a much larger and a more robust species with thick cylindrical body (BW 4.2–4.5 mm in adults) and can be easily diagnosed from a much more slender in D. tropcentr (BW 1.3–2.4 mm in adults).
Furthermore, Dibamus deimontis sp. nov. can be readily distinguished from D. tropcentr in a number of osteological traits (for osteological description of D. tropcentr see Kliukin et al. 2023). Dibamus deimontis sp. nov. has slightly lower number of presacral vertebrae (115 vs. 118) and significantly lower number of tail vertebrae (27 vs. 36). Although the pelvic girdle contains the same set of bones in both species, in Dibamus deimontis sp. nov. pubis, ischium and ilium are fused, while in D. tropcentr all pelvic girdle bones remain separate. Dibamus deimontis sp. nov. can be further diagnosed from D. tropcentr by the presence of the sagittal ridge on the parietal (vs. absence) and by having a higher number of labial foramina on maxillae (6–7 vs. 3–4); by having prefrontal bone quadrate in shape (vs. triangular), and by anterolateral processes of frontal bones present (vs. absent). The new species can be further distinguished from D. tropcentr by having exoccipitals, basioccipitals, supraoccipitals, opisthotics and prootics fused, while in D. tropcentr exoccipitals and opisthotics are fused, and the remaining bones are separate. In the new species the parabasisphenoid complex is fused with the basioccipital, while D. tropcentr has a distinct suture between these parts of the skull. Dibamus deimontis sp. nov. has a higher number of labial foramina on the each side of the dentary (5 vs. 2 in D. tropcentr ) and generally higher number of teeth on the dentary (7–8 vs. 4–5). Both Dibamus deimontis sp. nov. and D. tropcentr remarkably differ from all other congeners for which skull morphology was described by the absence of a separate coronoid bone in the lower jaw.
Distribution and natural history notes. All specimens of Dibamus deimontis sp. nov. were collected from March 1 to March 3, 2023 during daytime (from 10:00 h to 15:00 h) and night (from 20:00 h to 22:00 h) excursions at elevations from 670 to 700 m a.s.l. The habitat represents perhumid mixed montane evergreen forest intermittent with large open areas covered with grasses ( Fig. 6 View FIGURE 6 ); with narrow valleys and temporary brooks with large granite boulders and rocks. The evergreen forest with dominated by different species of palm trees ( Arecaceae ) along with Quercus poilanei Hickel & A.Camus , Distylium sp. , Garcinia hanburyi Hook.f. , Gluta wrayi King , Parinari anamensis Hance , Syzygium zeylanicum (L.) DC., Vatica harmandiana Pierre , Ilex cochinchinensis (Lour.) Loes. , Calophyllum ceriferum Gagnep. ex P.F.Stevens , and Cinnamomum cambodianum Lecomte with the presence of Dacrycarpus imbricatus (Blume) de Laub. , Podocarpus neriifolius D. Don , and Nageia wallichiana (C.Presl) Kuntze. Large rocks and boulders ca. 2–5 m in diameter along the stream beds were covered by a thick layer of mosses intertwined with roots of young undergrowth, rhizomes of ferns, and leaf litter. The lizards were found on the surface of granite rocks or inside the layer of mosses and roots. We recorded from one to three specimens of different sex and age on the same rock. When disturbed, the lizards would wiggle in an attempt to escape and fall down from the rocks.
On the same rocks under the moss layer we also recorded Sphaerotheriida Brandt and Glomerida Brandt millipedes, Scolopendromorpha centipedes and earthworms of the family Megascolecidae Rosa. On one stone together with a specimen of Dibamus deimontis sp. nov. we recorded a specimen of a kukri snake, Oligodon annamensis Leviton ( Colubridae ). Other herpetofaunal species co-occurring in the same forest included: Scincella rufocaudata Darevsky & Nguyen ( Scincidae ), Cyrtodactylus caovansungi Orlov, Nguyen, Nazarov, Ananjeva & Nguyen , C. sangi Pauwels, Nazarov, Bobrov & Poyarkov , Gekko cf. russelltraini Ngo, Bauer, Wood & Grismer ( Gekkonidae ), Bronchocela vietnamensis Hallermann & Orlov ( Agamidae ), and Trimeresurus cf. macrops Kramer ( Viperidae ).
Conservation status. Dibamus deimontis sp. nov. is to date known only from eight specimens collected from one locality within ca. 500 m 2 area on the slopes of Nui Chua Mt. within Nui Chua N.P., Ninh Thuan Province, southern Vietnam. Though these peculiar lizards appear to be locally abundant, it seems that their populations are locally concentrated in quite small areas. Any specific threats to Dibamus deimontis sp. nov. are not known, however the new species is likely associated with the specific microhabitats found in high-elevation perhumid montane evergreen forests. Most likely, the new species is endemic to Nui Chua Mt. and, may be, the adjacent peaks within the territory of Nui Chua N.P. (see Fig. 1 View FIGURE 1 ). Given the available information, we suggest Dibamus deimontis sp. nov. to be considered as a Vulnerable (VU) species following IUCN’s Red List categories ( IUCN Standards and Petitions Subcommittee 2019).




| ZMMU |
Zoological Museum, Moscow Lomonosov State University |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.