
FAVIINAE GREGORY, 1900: 29
|
publication ID |
https://doi.org/ 10.1111/j.1096-3642.2012.00855.x |
|
persistent identifier |
https://treatment.plazi.org/id/B26DA91C-6D09-1729-E00A-FDB0D2B5F9C5 |
|
treatment provided by |
Marcus |
|
scientific name |
FAVIINAE GREGORY, 1900: 29 |
| status |
|
SUBFAMILY FAVIINAE GREGORY, 1900: 29 .
Type genus: Favia Milne Edwards, 1857
Original description: ‘Compound Aporosa in which the corallum is massive or dendroid; the septa are numerous, imperforate, radial and usually hexameral. The endotheca is dissepimental. Growth is in the main by fission.’ ( Gregory, 1900: 29).
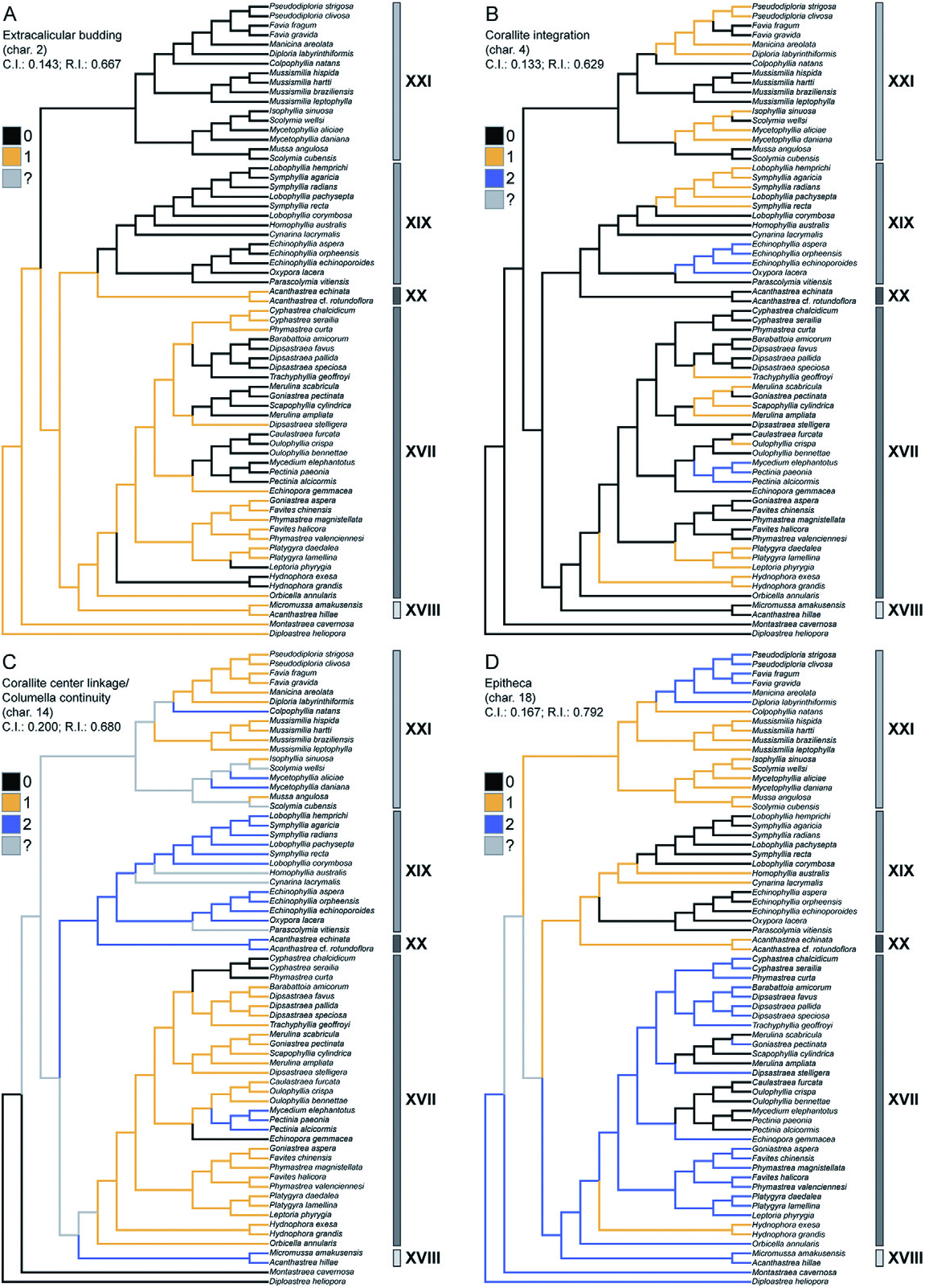
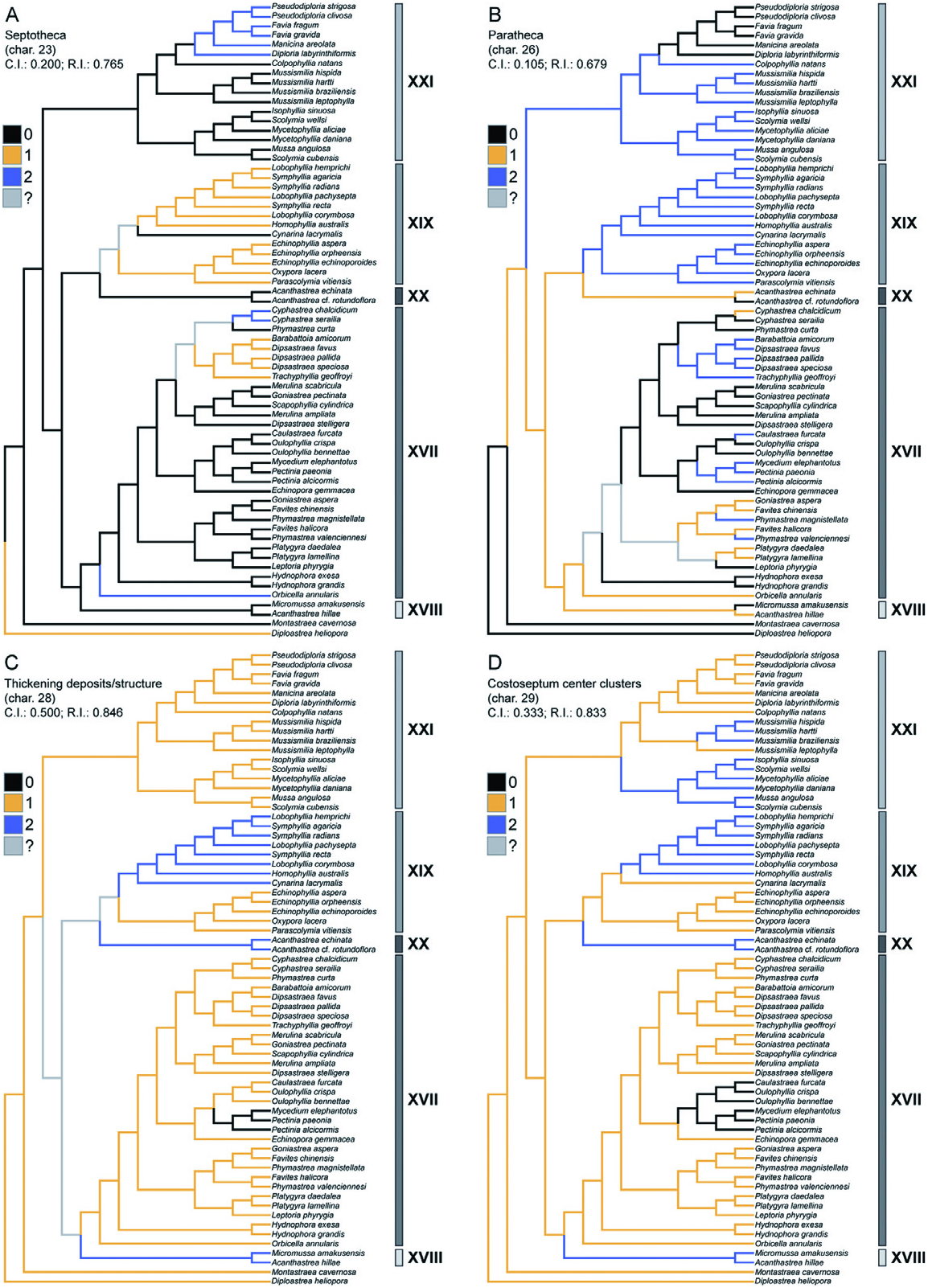
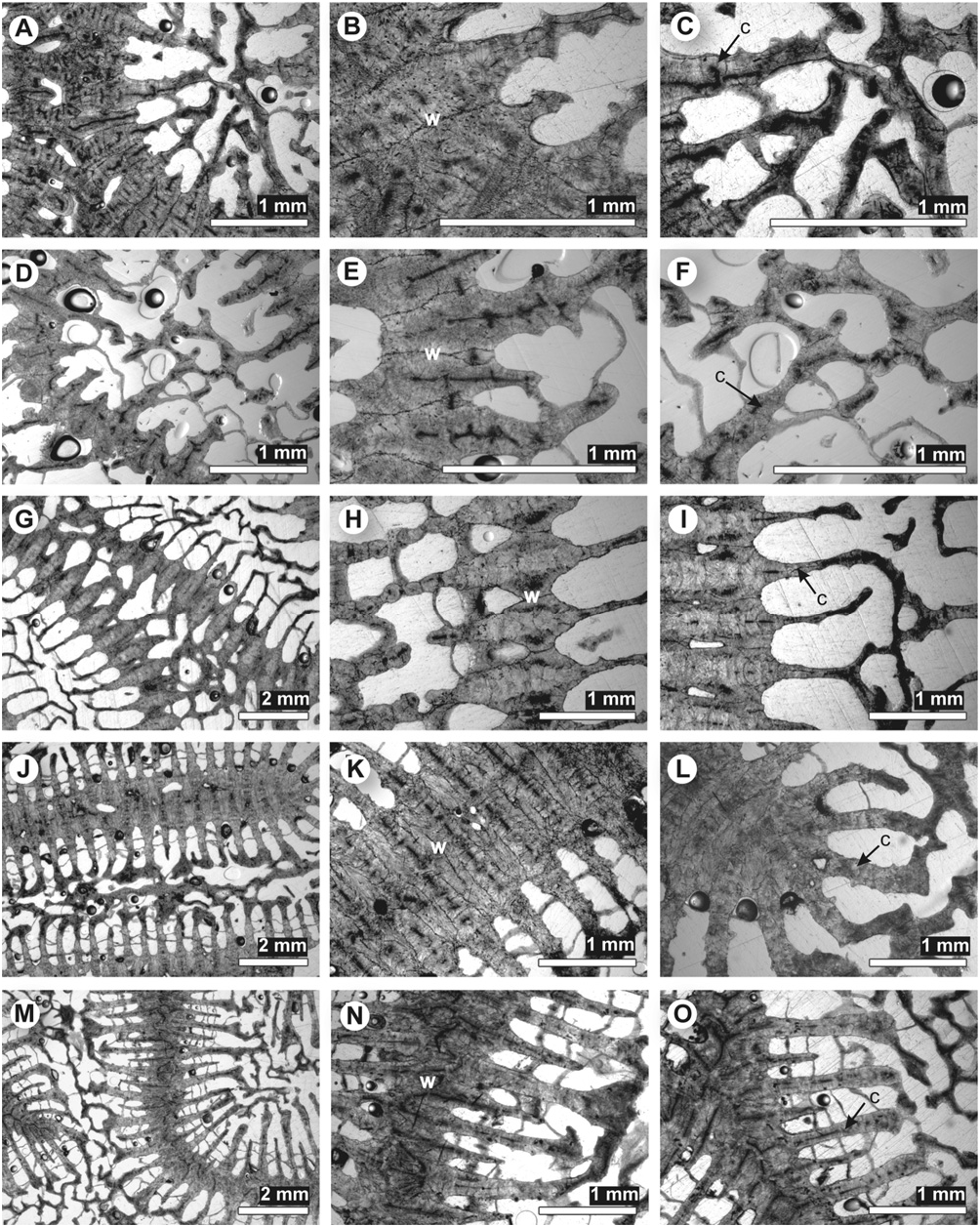
Diagnosis: Macromorphology: colonial only; corallites discrete or arranged in uniaxial series (includes plocoid, meandroid, and phaceloid forms); coenosteum usually present, although limited; costosepta not confluent; epitheca reduced or well developed; septal lobes (except Mussismilia ) ( Fig. 2 View Figure 2 ).
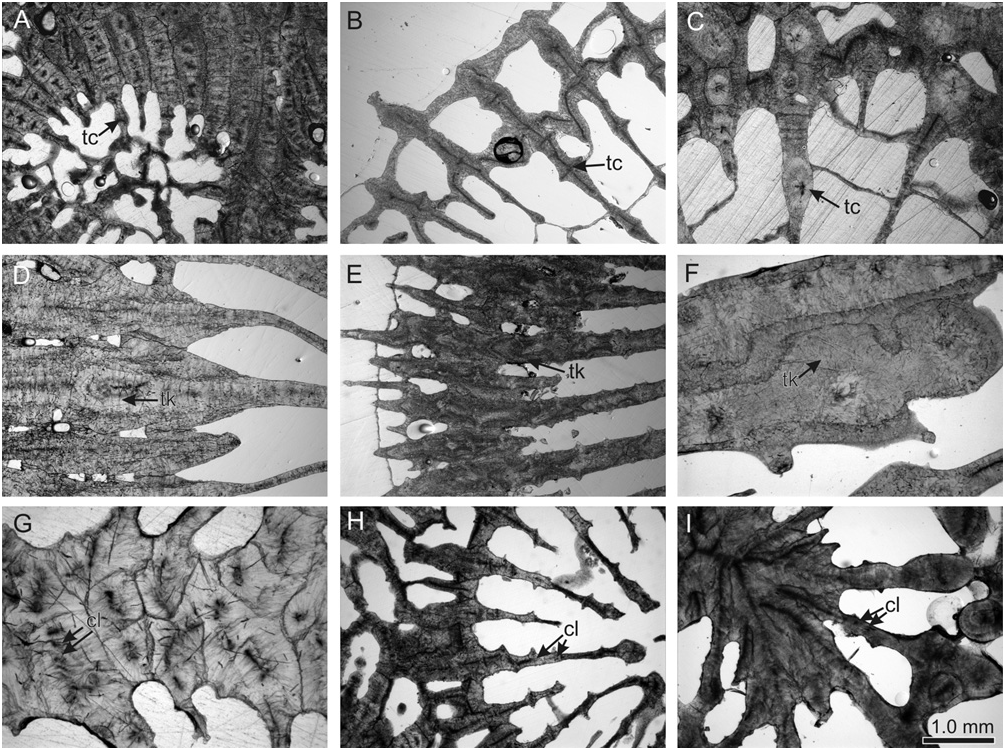
Micromorphology: regular, narrowly spaced (<1 mm), tricorne or fan-shaped septal teeth (never spine-shaped or triangular), with elliptical bases orientated transverse to the septal plane; interarea of teeth smooth or consisting of horizontal bands; septal granules strong and aligned ( Figs 3 View Figure 3 , 4 View Figure 4 ).
Microstructure: septothecal or parathecal corallite walls, containing occasional trabeculothecal elements; clusters of centres of calcification within the costosepta and columella weakly developed (except Mussismilia ), narrowly separated (<0.6 mm), and connected by medial lines; carinae usually well developed ( Figs 5 View Figure 5 , 6 View Figure 6 ).
Genera included:
Favia Milne Edwards, 1857 View in CoL
Colpophyllia Milne Edwards & Haime, 1848 View in CoL Diploria Milne Edwards & Haime, 1848 View in CoL
Manicina Ehrenberg, 1834 View in CoL
Mussismilia Ortmann, 1890 View in CoL
Pseudodiploria Fukami, Budd & Knowlton View in CoL gen. nov.
Remarks: In the classification system of Vaughan & Wells (1943: 153) and Wells (1956: F400), the family Faviidae is distinguished by having a septothecal or parathecal wall; simple trabeculae, in one or two fan systems; dentate septal margins; dissepiments; and relatively small corallites (<10 mm in diameter). As herein defined, the subfamily Faviinae is monophyletic ( Fig. 7 View Figure 7 ) and restricted to taxa whose septal teeth are regular and narrowly spaced, with elliptical bases orientated transverse to the septal plane (i.e. paddleshaped), and whose septal granules are strong, spiky, and aligned.
Unlike the subfamily Mussinae , which possesses spine-shaped or triangular teeth and diffuse granulation, the septal teeth of the Faviinae are tricorne or fan-shaped (elliptical bases); and septal granulation is well developed (evenly scattered or organized in lines).
Traditional Atlantic ‘faviids’ (including Montastraea cavernosa and the Orbicella annularis complex) are also distinguished from traditional Indo-Pacific ‘faviids’ on the basis of tooth shape. Indo-Pacific taxa have irregular spine-shaped or multidirectional teeth (often lacerate); whereas Atlantic taxa (excluding the Orbicella annularis complex) have regular, blocky teeth that are often paddle-shaped (see description in Budd & Stolarski, 2011).
Distribution: Atlantic only.
GENUS FAVIA MILNE EDWARDS, 1857: 426 View in CoL
( FIGS 10A–D View Figure 10 , 15A–D, 20A–F View Figure 20 , 24A–F View Figure 24 )
[not Oken, 1815: 67, because all taxa in Oken book rejected by ICZN opinion 417 ( ICZN Commission, 1956)]
Synonyms: None.
Type species: Madrepora fragum Esper, 1795: 79 , pl. 64, figs 1, 2 [= Madrepora ananas Pallas, 1766 ]; by subsequent designation, Verrill, 1901: 88–91. Holotype is lost ( Scheer, 1990; Cuif & Perrin, 1999). We herein designate specimen MNHN-scleFAV560 collected by Mr Ricord in Haiti ( Cuif & Perrin, 1999) as the neotype ( Fig. 10C, D View Figure 10 ).
Original type species locality: ‘südlichen Americanischen Meeren’ ( Esper, 1795: 80) [Recent].
Original descriptions:
1. ‘Röhren (=walls) in einem kitt, öffnen sich oben gleich hoch, laufen in leinen gemeinschaftlichen Stamm zusammen.’ ( Oken, 1815: 67).
2. ‘Le polypier se compose de polypiérites unis entre eux par des côtes plus ou moins développées et une exothèque celluleuse. Les calices coservant des bords libres, soit subcirculaires, soit ovalaires. La columelle est spongieuse. Les cloisons sont débordantes; leurs dents internes sont les plus grandes et souvent même simulent des palis. L’endothèque est bien développée.’ ( Milne Edwards, 1857: 426–427).
Subsequent morphological descriptions ( Matthai, 1928 and later): Not in Matthai, 1928 (but see Matthai, 1919); Vaughan & Wells (1943: 163, 166); Wells (1956: 402); Laborel (1969: 191–198); Cuif & Perrin (1999: 137–156); Walton Smith (1971: 79–80); Zlatarski & Estalella (1982: 61–66); Veron (2000: vol. 3: 100–131).
Diagnosis: Macromorphology: plocoid; discrete (one to three centres per series); costate coenosteum; small calices (<4 mm) with three or more septal cycles; continuous, compact trabecular columella; well-developed epitheca; moderate endotheca; small septal lobes ( Figs 10A–D View Figure 10 , 15A–D).
Micromorphology: distinctive paddle-shaped teeth, occasionally tricorne, with elliptical bases orientated transverse to the septal plane; smooth interarea of teeth; spiked granules organized in lines; teeth in major and minor septal cycles differ slightly in size ( Fig. 20A–F View Figure 20 ).
Microstructure: septothecal wall; small (<0.6 mm), well-defined clusters of calcification centres within costosepta, forming carinae orientated transverse to the septal plane; layered fibrous thickening deposits ( Fig. 24A–F View Figure 24 ).
Species included: Favia fragum ( Esper, 1795: 79, pl. 64, figs 1, 2). Holotype is from ‘südlichen Americanischen Meeren’, and is lost; neotype (herein designated) = MNHN-scleFAV560, Figure 10C, D View Figure 10 , Haiti.
Favia gravida Verrill, 1868: 354 View in CoL [syntypes = YPM1465A, B; Fig. 15C, D, Abrolhos Archipelago, Brazil] (= Favia conferta Verrill, 1868: 355 View in CoL ) [syntypes = YPM1466A, B; YPM8268, Abrolhos Archipelago, Brazil].
Remarks: The genus Favia was originally described by Oken (1815: 67), who listed ‘ Favia ananas ’ first as belonging to the genus. However, according to ICZN opinion 417 ( ICZN Commission, 1956), the names originally proposed by Oken (1815) are rejected. Therefore authorship is based on subsequent use of the name. Ehrenberg (1834) was the second to use the name, but assigned an assortment of species to the genus, all of which have been subsequently assigned to other genera. Cuif & Perrin (1999) indicate ‘Milne-Edwards, 1857’ as the author of Favia , after Verrill (1901). This usage is followed here.
In the classification system of Vaughan & Wells (1943) and Wells (1956), the genus Favia is distinguished by having a plocoid colony form; a costate coenosteum; a parietal (= trabecular) columella; and intracalicular budding. Septal margins are strongly dentate; trabeculae are usually simple, in one or two fan systems; and corallites are relatively small (<10 mm) ( Vaughan & Wells, 1943: 153–154, 163, 166). As discussed above, it is distinguished from Indo-Pacific ‘ Favia ’, herein referred to as Dipsastraea de Blainville, 1830 , on the basis of its paddle-shaped septal teeth and septothecal corallite wall structure.
Our work shows that species traditionally assigned to the genus vary considerably in corallite wall structure (including parathecal, septothecal, and trabeculothecal walls) and in the shape of their septal teeth. We hereby restrict the definition of the genus to species with septothecal walls and multicentred paddleshaped teeth orientated transverse to the septal plane, three characters that the genus Favia shares with Manicina as defined below. Favia is distinguished from Manicina on the basis of its short corallite series (= valleys), small calice width (<4 mm) with low relief, its regular minor septa, and its compact (one to three threads) columella. In addition, the corallite walls of Favia are exclusively septothecal, whereas those of Manicina contain trabeculothecal elements.
Following Laborel (1969), Cairns, Hoeksema & Land (1999) listed two species of Favia ( Favia fragum , Favia gravida ) with septothecal walls from the Caribbean and Brazil; however, both Zlatarski & Estalella (1982) and Veron (2000) synonymized the two species. Walton Smith (1971) recognized F. fragum as distinct, and further split F. gravida into two species ( F. gravida , F. conferta ). Here we list and illustrate two species of Favia ( F. fragum , F. gravida ) following Nunes et al. (2008), who showed F. fragum and F. gravida to be genetically distinct. Favia gravida ( Fig. 15C, D) is distinguished from F. fragum ( Fig. 15A, B) on the basis of its more numerous septa (four complete septal cycles), somewhat longer series, higher calice elevation, and taller, more widely spaced septal teeth.
Following the molecular results of Fukami et al. (2004, 2008), we restrict the definition of the genus Favia to include only Atlantic taxa. The emended genus does not include the following species, which were assigned to it by Veron (2000) and are herein assigned to the genus Dipsastraea de Blainville, 1830 [type species: Madrepora favus Forskål, 1775 (SD: Wells, 1936)].
Species Assigned to Dipsastraea :
1. Favia favus ( Forskål, 1775: 132) [lectotype = ZMK28, unknown locality]; not seen but illustrated in Veron et al., 1977: 26 (fig. 28).
2. Favia albida Veron, 2000 (3): 112 [‘Holotype’ in Veron, 2002: 140 = G55788 (MTQ), Sharm al-Sheikh, Sinai Peninsula, Egypt], seen.
3. Favia danai ( Milne Edwards, 1857: 442) [holotype = USNM00032 About USNM , Tonga-Tabou], seen .
4. Favia helianthoides Wells, 1954: 458 [holotype = USNM44980 About USNM , Bikini Atoll], seen .
5. Favia lacuna Veron, Turak & DeVantier ; in Veron, 2000 (3): 111 [‘holotype’ in Veron, 2002: 139 = G55836 (MTQ), northern Red Sea coast of Saudi Arabia], seen.
6. Favia laxa ( Klunzinger, 1879: 49) [holotype = ZMB-Cni2193, Red Sea]; seen .
7. Favia lizardensis Veron & Pichon ; in Veron et al. (1977: 45 [holotype = BM (NH)1977.1.1.2, MacGillivray , eastern Australia], seen.
8. Favia maritima ( Nemenzo, 1971: 169) [syntypes = UP C-859, C-861, Puerto Princesca Bay , Palawan, Philippines]; not seen .
9. Favia marshae Veron, 2000 (3): 122 [‘holotype’ in Veron, 2002: 145 = WAM Z12910 View Materials ], Ashomore Reef, north-west Australia, not seen.
10. Favia matthaii Vaughan, 1918: 109 [holotype = USNM38381 About USNM , Indian Ocean], seen .
11. Favia maxima Veron & Pichon ; in Veron et al. (1977): 43 [holotype = BM (NH)1977.1.1.1, Hook Island , Whitsunday Islands, Great Barrier Reef], seen .
12. Favia pallida ( Dana, 1846: 224) [syntype = USNM00076 About USNM , ‘Feejee Islands’], seen .
13. Favia rosaria Veron, 2000 (3): 119 [‘holotype’ in Veron, 2002: 143 = G55822 (MTQ), Milne Bay, eastern Papua New Guinea], seen.
14. Favia rotumana ( Gardiner, 1899: 750) [Neotype = ZMA Coel. 5686, Rotumana, Fiji], not seen but illustrated in Wijsman-Best, 1972: pl. 3, fig. 2.
15. Favia rotundata (Veron & Pichon) ; in Veron et al. (1977): 64 [holotype = BM (NH)1977.1.1.6, southwest Swain Reefs , eastern Australia], not seen .
16. Favia speciosa Dana, 1846: 220 [syntype = USNM00037 About USNM , ‘ East Indies’ ], seen .
17. Favia stelligera ( Dana, 1846: 216) [syntype = USNM00055 About USNM , ‘Feejee Islands’], seen .
18. Favia truncata Veron, 2000 (3): 113 [‘holotype’ in Veron, 2002: 142 = G55836 (MTQ), Milne Bay, eastern Papua New Guinea], seen.
19. Favia veroni Moll & Borel Best, 1984: 48 [holotype = RMNH 15209 About RMNH , Kudigareng Keke , Spermonde Archipelago, Indonesia], seen .
20. Favia vietnamensis Veron, 2000 (3): 127 [‘holotype’ in Veron, 2002: 146 = G55859 (MTQ), Nha Trang, Vietnam], seen.
Note: Following ICZN Commission (2011: 162–166), the species named in Veron, (2000) are valid. However, the type specimens designated in Veron (2002) are not valid.
The Atlantic species Favia leptophylla Verrill, 1868: 353 [holotype = YPM1517, Abrolhos Archipelago, Brazil] is transferred for the first time to the genus Mussismilia , as described below under Mussismilia .




| BM |
Bristol Museum |
| UP |
University of Papua and New Guinea |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
FAVIINAE GREGORY, 1900: 29
| Budd, Ann F., Fukami, Hironobu, Smith, Nathan D. & Knowlton, Nancy 2012 |
FAVIINAE GREGORY, 1900: 29
| Gregory JW 1900: 29 |
Favia gravida
| Verrill AE 1868: 354 |
| Verrill AE 1868: 355 |
FAVIA MILNE EDWARDS, 1857: 426
| Milne Edwards H 1857: 426 |