
Bartlett, Bahder & Bartlett & Barrantes Barrantes & Zumbado Echavarria & Humphries & Helmick & Ascunce & Goss, 2019
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4577.3.5 |
|
publication LSID |
lsid:zoobank.org:pub:5A2E47BD-8AD0-4F21-B6CC-F0475273CC4D |
|
DOI |
https://doi.org/10.5281/zenodo.5944511 |
|
persistent identifier |
https://treatment.plazi.org/id/B36AB07A-211E-FF87-8BEC-1D31FDB7707B |
|
treatment provided by |
Plazi |
|
scientific name |
Bartlett |
| status |
subgen. nov. |
Subgenus Agoo Bahder & Bartlett subg. n.
( Figs 2–6 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 )
Type species: Omolicna (Agoo) xavieri Bahder & Bartlett sp. n.
Diagnosis. Frons narrower and paranota more strongly foliate than subgenus Omolicna . Ventral lobe of pygofer (ventral view) broad, distally attenuating to rounded apex. Aedeagus and endosoma nearly bilaterally symmetrical. Segment 10 elongate, ventrally sinuate (lacking convexity found in most Omolicna s.s.).
Remarks. The composition of subgenus Agoo subg. n. is limited to Omolicna xavieri for the time being. At present, not all taxa of Omolicna are available for consideration of subgeneric status—this task is considered beyond our current scope and intended for future work. The erection of the new subgenus Agoo for Omolicna xavieri sp. n. is supported by molecular evidence (see below) that places this species as an outlier relative to O. uhleri (Ball) , O. joi , O. triata (Caldwell) and O. brunnea (McAtee) .
Etymology. The subgenus name was picked after the lead author asked his four-month-old daughter what name he should choose. Her response was to blow a raspberry, followed by ‘agoo’. Due to the difficulty in spelling the sound of a raspberry, the latter was selected as the subgenus name.
Omolicna (Agoo) xavieri Bahder & Bartlett sp. n.
Type locality. La Virgen de Sarapiqui , Heredia, Costa Rica .
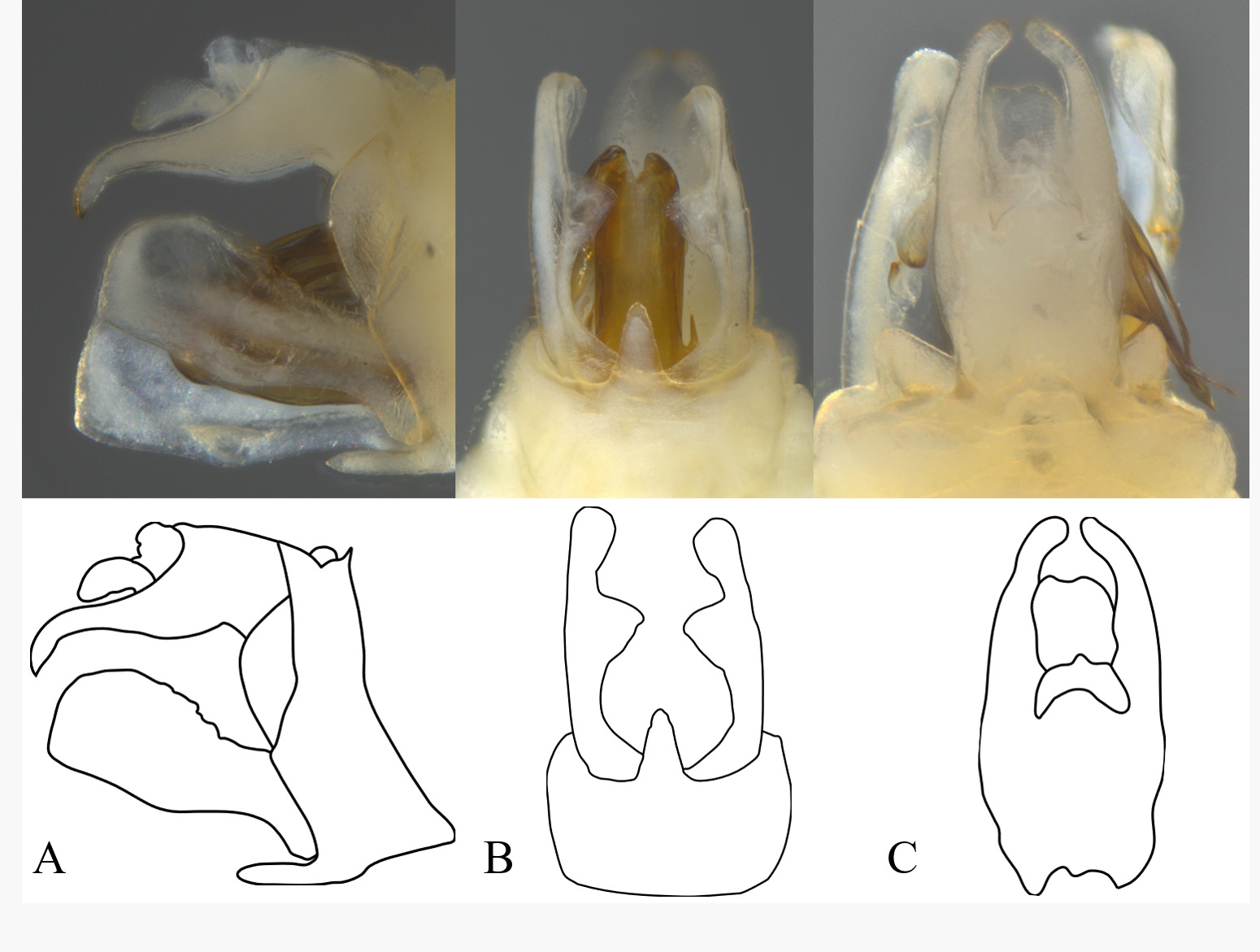
Diagnosis. Male pygofer with median ventral process broad near base, attenuating distally to broadly rounded apex (lateral teeth lacking). Segment 10 (= “anal tube”) in lateral view elongate, caudal portion elongate, narrow, and ventrally sinuate, apex acute and distinctly downcurved; in dorsal view, deeply notched on caudal aspect. Phallotheca with lateral elongate, acuminate tooth on each side near base; apex with 4 elongate processes on each side, 2 pair at apex, 2 pair subapically, in addition to a complex endosoma.
Description. Color. General body color yellowish-brown to orange; face deep orange above frontoclypeal suture and beneath antennae, lateral carinae of front and vertex darker. Mesonotum diffusely infuscate with orange. Wings, faintly yellowish, strong veins darkened, middle regions of remigium darkened so that in dorsal view wings appear to bear a dark vitta. Dorsum of abdomen orangish, aedeagus appearing dark brown within paler terminalia.
Structure. Body length males: 5.6–6.2 mm (n = 9) with wings; females: 8.0– 8.2 mm (n = 9) with wings and males: 3.3–3.8 mm without wings; females: 4.2–4.7 mm without wings. Head. Anterior margin of head, in lateral view, rounded ( Fig. 3C View FIGURE 3 ). Vertex deeply concave posteriorly, notched distally ( Fig. 3B View FIGURE 3 ), broadest near base, tapering distally; lateral margins decidedly keeled, bearing 2 (somewhat irregular) rows of sensorial pits, disc depressed, without distinct carinae. Transverse apical carina separating vertex from frons absent. Vertex length males: 0.3–0.4 mm; females: 0.4–0.5 mm. Vertex width at hind margin males: 0.19–0.24 mm; females: 0.30–0.34 mm. Vertex width at distal margin males: 0.06–0.07 mm; females: 0.08–0.09 mm. Frons with lateral carinae strongly keeled, narrowest between compound eyes, diverging slightly ventral to become parallel between ocelli and frontoclypeal suture ( Fig. 3A View FIGURE 3 ); sensorial pits next to each carina for entire length, carinae absent on disc of frons. Frons length males: 0.65–0.77 mm; females: 0.9 4– 0.98 mm. Frons dorsal width males: 0.11–0.17 mm; females: 0.19–0.22 mm. Frons frontoclypeal margin width, males: 0.19–0.25 mm; females: 0.26–0.30 mm. Clypeus with lateral carinae keeled, sensorial pits absent, converging near midlength to labrum. Clypeus length males: 0.39–0.56 mm; females: 0.58–0.60 mm.
Thorax. Pronotum short, anterior margin following contours of posterior margin of head ( Fig. 3B View FIGURE 3 ; convex, anteriorly truncate behind vertex, narrowed behind eyes); posterior margin weakly concave; paranotal regions strongly foliate ( Figs 3 View FIGURE 3 A–C), in lateral view forming deep fossae, forming posterior cup for the antennae, partially surrounding eyes both dorsad and ventrad. Pronotum length at midline males: 0.19–0.24 mm; females: 0.33–0.36 mm. Mesonotum appearing slightly elevated in lateral view ( Figs 2A View FIGURE 2 , 3C View FIGURE 3 ), in dorsal view with 3 subparallel longitudinal carinae, indistinctly reaching posterior margin. Mesonotum length at midline males: 0.75–0.91 mm; females: 1.00– 1.04 mm. Mesonotum width males: 1.02–1.11 mm; females: 1.40–1.47 mm.
Forewing ( Fig. 6 View FIGURE 6 ) with a row of sensory pits along basal 2/3 rd of Sc+R (nearly to fork of Sc) and basal half of postcubitus. Forks of R and CuA veins at approximately the same level, both well proximad of the claval apex. Claval apex just beyond midpoint of wing, fork of M just distad of claval apex; R 3-branched, M 4-branched and CuA 3-branched. The junction of the Pcu+A1 occurs in the distal half of the wing (A1 closely approximated to the trailing margin of the clavus). Forewing length males: 4.9–5.9 mm; females: 5.7–6.9 mm.
Terminalia. Pygofer, in lateral view, narrow, distal and proximal margins sinuate ( Fig. 4A View FIGURE 4 ); broadest ventrally. In ventral view, ventral opening of pygofer bearing a rounded lobe ( Fig. 4B View FIGURE 4 ), widest at base and attenuating distally to rounded apex. Gonostyli (= parameres), in ventral view, broadest ventrally, nearly meeting medially at base, then abruptly narrowed with large, rounded tooth just past midlength, slightly hooked medially at apex; in lateral view, dorsal and ventral margin sinuate, concave and narrowest in basal third, broadened distally to broadly rounded apex, dorsal projection absent. Phallotheca with lateral pair of large subbasal acuminate teeth, one each on left and right sides ( Fig. 5A, B View FIGURE 5 ); apex with pair of very long retrorse processes on both left and right side, and elongate subapical retrorse processes, the most distal very broad at base, strongly narrowed to acuminate apex, proximal process connected near base of previous, sinuate and asymmetrical on left and right sides. Endosoma complex, retrorse, bearing additional projections and asymmetrical 3-dimensional structures, but appearing to have 2 dorsal flattened projections, 2 ventral flattened processes, and two elongate ventral spines. Segment 10 (= “anal tube”), in lateral view ( Fig. 4A View FIGURE 4 ), elongate, caudally narrow and sinuate, apex curved downward with acute apex; in caudal view, deeply notched ( Fig. 4C View FIGURE 4 ).
Plant associations. Coconut palm ( Cocos nucifera ), Arecaceae .
Distribution. Costa Rica (Heredia, Tortugeuro).
Etymology. The specific name given is a reference to first name of the lead authors first born son, Xavier Jacques Bahder.
Material examined. Holotype male “ Costa Rica, Heredia / La Virgen de Sarapiqui / Brian W. Bahder; 11 May 2018 / aspirated from coconut // Holotype / Omolicna /xavieri” ( FSCA) ; Paratypes, Tortuguero [12 May 2018] (9 males, 10 females, FLREC and FSCA) .
Sequence Data. A total of 703 bp of the COI gene was sequenced for O. xavieri and submitted to GenBank (Accession No. MK443068 View Materials ). In addition, 703 bp of the COI gene were sequenced for O. triata (Accession No. MK443069 View Materials ) and O. brunnea (Accession No. MK443070 View Materials ). For the region sequenced, no other Omolicna species were represented in BLAST hits, with the closest match being Kallitaxila sinica (Walker) ( MG099955 View Materials ) ( Tropiduchidae ) at 99% query coverage and 82% similarity. Omolicna joi is present in GenBank (Accession No. KF472312 View Materials ) while O. fulva (CNCHG1424-12.COI-5P) and O. uhleri (CNCHG1197-12.COI-5P) were both present in the Barcode of Life Database (BOLD). All sequences were aligned and compared to assess variability among the currently available Omolicna for the region that does overlap among all sequences. For the COI sequences available, pairwise genetic distance values comparing percent nucleotide difference ranges from 24.4% to 31.1% between O. xavieri sp. n. and the other species included ( Table 3). Among O. triata , O. brunnea , O. joi , and O. uhleri there was between 14.4% and 19.5% variation for the region analyzed ( Table 3). Finally, O. fulva differed by between 24.4% and 30.4% from all other species in the study ( Table 3).
A total of 1,493 bps for the 18S gene was sequenced for O. xavieri (Accession No. MK443073 View Materials ) in addition to O. brunnea (Accession No. MK443071 View Materials ), O. triata (Accession No. MK443072 View Materials ), and O. joi (Accession No.) . Overall, O. xavierii differed by about 10.6% from the other species of Omolicna ( Table 4) whereas among O. joi , O. brunnea , and O. triata , there was between 0.1% to 0.9% of pairwise differences ( Table 4).
Remarks. Omolicna xavieri sp. n. is unusual among Omolicna species in possessing a patterned face (beneath the antennae and along the frontoclypeal margin, the faint pattern on the dorsum of the thorax and on the wings). It is also unusual in that the ventral margin of the anal tube lacks the convexity found commonly (but not uniformly) in the genus. The new species also appears to differ qualitatively from many congeners in that the paranotal regions of the pronota are more expansively foliate, and the frons more narrowed than most of the available congeners.
The aedeagus in the new species is also distinct compared to other Omolicna species in that the spines are more elongate and slender. Additionally, the aedeagus in the new species is noticeably more symmetrical than that observed in other members of the genus.




| FSCA |
Florida State Collection of Arthropods, The Museum of Entomology |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.