
Lepidozia bragginsiana E.D.Cooper et M.A.M.Renner, 2014
|
publication ID |
https://doi.org/ 10.11646/phytotaxa.173.2.2 |
|
persistent identifier |
https://treatment.plazi.org/id/B40087D7-5E5E-FFCE-41FA-FBE126488A5D |
|
treatment provided by |
Felipe |
|
scientific name |
Lepidozia bragginsiana E.D.Cooper et M.A.M.Renner |
| status |
sp. nov. |
Lepidozia bragginsiana E.D.Cooper et M.A.M.Renner View in CoL sp. nov. Figs 2 View FIGURE 2 , 3A View FIGURE 3 , 4A–C View FIGURE 4 .
Type:— New Zealand, South Island , Westland , Grey River catchment, Paparoa Range , Blackball Stream, Blackball Road, 42°20’22”S 171°23’44”E, 230 m, 17 Feb 2012, M. A. M GoogleMaps . Renner 6190 (holotype: AK; isotypes: NSW, F) GoogleMaps .
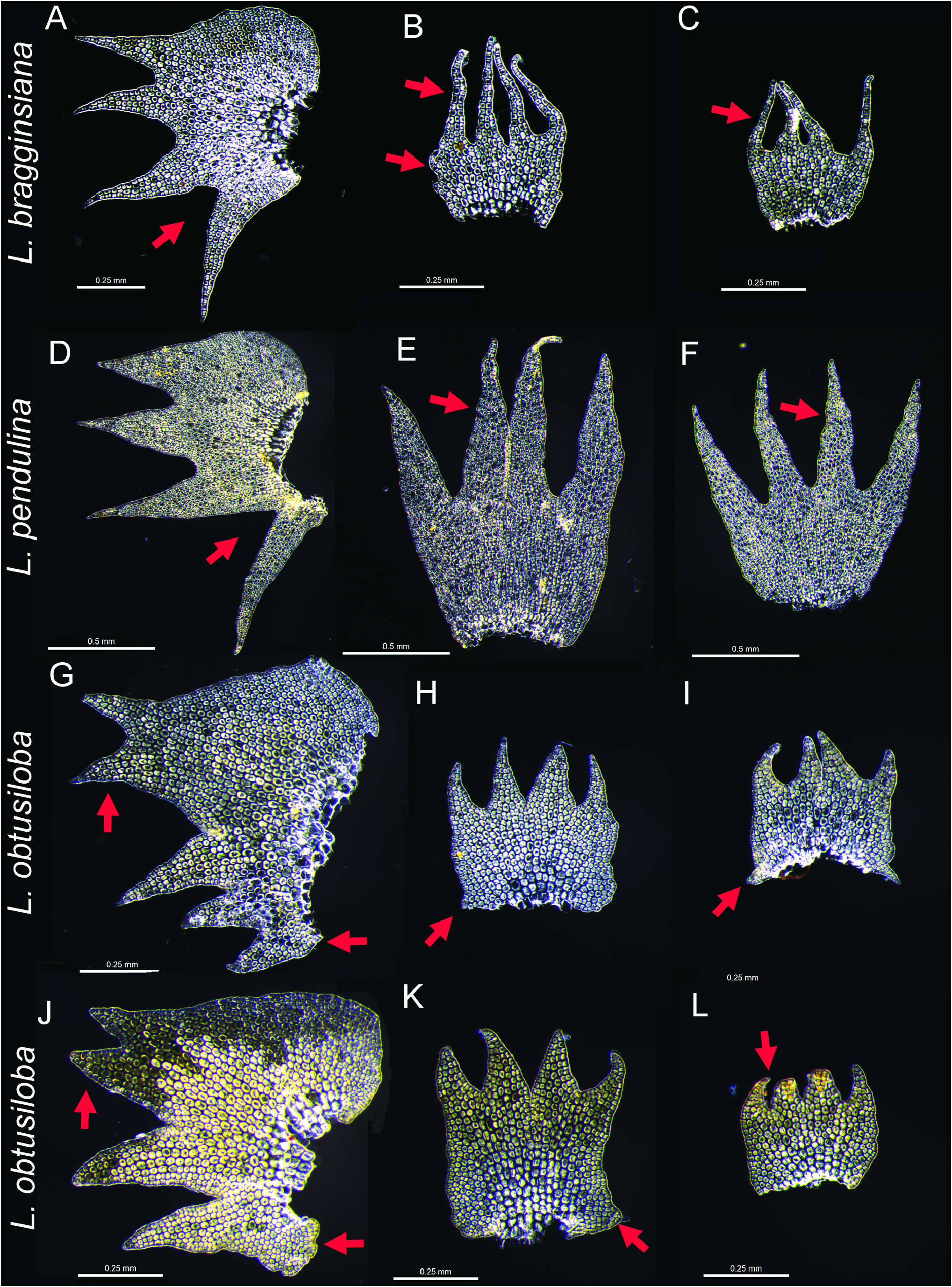
Diagnosis:— Distinctive within Lepidozia in its bipinnate branching, erect primary shoots and turf forming habit; leaves with an auriculate antical base and occasional production of accessory spurs or projections on the postical margin, projections often small and tooth-like but variably taking a range of forms through to having the size and shape of the adjacent postical leaf-lobe; leaf-lobe cells with evenly and continuously thickened cell walls, underleaves of primary shoots equal or narrower in width to stem, with a small projection on each side of the medial or upper disc margin, lobes attenuate, to 0.5× the underleaf length, often asymmetrically lobed, 4(-6) cells wide at their base; underleaves on secondary shoots wider than stem, with rounded to toothed bases, slightly larger in absolute size than underleaves on primary shoots, symmetrically lobed.
Description:— Plants erect, rigid, with ventrally secund branches, pale green to golden yellow-green, shoots robust, to 2 cm wide (branch extremity to branch extremity). Branching mostly of the Frullania - type, pinnate or bipinnate, the latter with branches typically originating from the side of the branch toward the shoot apex, secondary shoots almost equal to the primary shoots in stature, though often becoming flagelliform; branch half-leaf 2-lobed, acroscopic margin faintly and irregularly denticulate, often one or both margins with a triangular or spine-like tooth near the disc base, first branch underleaf (3–)4-lobed, inserted on ventral lateral side of main axis in a position half way between leaves and underleaves of main shoot. Ventral intercalary branching rare and sporadic. Stems rigid, 400–520 µm diameter on main shoots, 155–270 µm diameter on branches, cortical cells in 1 layer of slightly larger, quadrate cells with evenly thickened, yellow-brown pigmented cell walls, external cell wall differentially thickened, approximately twice as thick as internal cell walls, internal to the cortical layer are 3–4 tiers of smaller cells whose walls are continuously but unevenly thickened, these cells transition to larger cells whose walls are thickened by triangular to bulging trigones, and faint continuous medial thickenings. Leaves slightly concave, contiguous to imbricate, weakly spreading to appressed to stem, the insertion distinctly incubous; on primary shoots 850–1150 µm at longest point, 1100–1450 µm at widest point, on secondary shoots 680–880 µm at longest point, 740–1125 µm at widest point; asymmetrically 4-lobed, dorsal lobe divided to 0.33, ventral lobe divided to 0.75, lobes broadly attenuate, acute, divergent to widely divergent, the ventral lobe postically displaced, widely divergent as a ‘claw’ particularly on primary shoot leaves, the dorsal lobes entire, 8–12 cells wide at base, the lobes terminating in a uniseriate row of 3–4 cells slightly longer than wide and thick-walled; disc distinctly asymmetrical, 17–25 cells high at dorsal sinus, 6–12 cells high at ventral sinus, the dorsal margin broadly ampliate, more or less cordate at base, acroscopic margin typically faintly and irregularly denticulate, occasionally with a single triangular tooth at one third from the base, particularly on primary shoot leaves; basiscopic margin entire to toothed with one or more accessory spurs, spines, or (rarely) lobes particularly on primary shoot leaves. Cells of dorsal disc margin and lobes isodiametric to rectangular, 12–15 µm long and wide, forming indistinct border 2–7 cells wide, wider on dorsal margin; cells of mid-disc distinctly thick-walled 18–20 µm wide by 22–25 µm long; basal cells larger still; cell surfaces smooth, unornamented. Oil-bodies not known. Underleaves on main shoots spreading to squarrose, 460–640 µm tall, 370–700 µm wide, symmetrically or weakly asymmetrically 4-fid to 0.5– 0.66 (median sinus), sinuses sharp, plane or reflexed, lobes narrow, attenuate, acuminate, 4–6 cell rows wide at base, margins entire but the septa of marginal cells usually weakly thickened and swollen, sharp at apex with a uniseriate row of 2–6 cells, disc 6–13 cells high at median sinus, the margins with a prominent triangular to spine-like tooth at the base. Underleaves on branch shoots spreading, usually symmetrically 4-fid to 0.5 (median sinus), rarely asymmetrical, sinuses U-shaped, plane, lobes 4 cell rows wide at base, attenuate, margins entire, disc 8–15 cells high at median sinus, the margins usually entire, subcordate at base, often with a single triangular to spine-like tooth. Asexual reproduction absent. Reproductive structures not seen.
Etymology:— named to honour Dr John E. Braggins (1944-), for his contribution to botany and bryology in New Zealand (see Anon. 2013). In particular the name recognises John’s role in guiding the liverwort collecting expeditions of many overseas researchers in New Zealand, including the trip that yielded the specimen from which his Lepidozia species was first recognised. It also recognises the inspiring role John has played in mentoring and encouraging a younger generation of bryologists trained in New Zealand.
Recognition:— The combination of bipinnate branching and asymmetric leaves with a wide sinus between the 3 rd and 4 th lobes, are characters that have been associated with L. pendulina , a large and widespread forest inhabiting species. The shared possession of these two characters may lead to confusion between L. bragginsiana and L. pendulina , and may explain the comment regarding branching variability by Engel & Glenny (2008) that small, poorly developed populations of L. pendulina have secondary branches weakly developed or absent. In our experience, L. pendulina is always a large, robust plant with a characteristic shoot architecture including well-developed secondary branches. In the field the combination of bipinnately branched shoot systems with remote and widely spreading leaves on primary shoots is diagnostic of L. pendulina ( Fig. 3 View FIGURE 3 ).
Other characters separating L. pendulina from Lepidozia bragginsiana , include 1) leaves lacking appendages on postical margin vs with appendages; 2) absence of a well-defined field of large cells in middle and base of leaf disc vs presence of such; 3) triangular underleaf lobes 10–13 cells broad at their base vs attenuate underleaf lobes 4 cells broat at their base ( Fig 4 View FIGURE 4 ).
Sporadic bipinnate branching also occurs in L. obtusiloba . Characters separating L. bragginsiana from L. obtusiloba and L. pendulina are summarized in Fig. 4 View FIGURE 4 .
The presence of ornamentation on the basal underleaf margins, and particularly the postical margin of the leaf lobe, has been associated with L. ornata Engel in Engel & Schuster (2001: 49). However, in L. ornata 1) the shoots are pinnately branched, and branching may be irregular and secondary shoots may differ in stature; 2) the sinus between 1 st and 2 nd leaf lobes is shallow, as in L. obtusiloba ; 3) the 4 th lobe points in same direction as other lobes; 4) the antical half of leaf is patent but lies in plane with the stem; 5) the underleaf margin at bottom of sinus between underleaf lobes is reflexed; 6) the underleaf lobes are typically 4–6 cells broad at base, and broader toward the apex; 7) accessory lobes on underleaves lobe-like rather than spine or spur-like; 8) the same is true for leaves; 9) accessory lamellae and flattened paraphyllia like appendages may occur on surface of leaf disc; 10) the underleaf lobe apices are obtuse, ending in one, two, or more cells, rarely if ever with uniseriate apical portion, and 11) the lobes are two or more cells wide for their entire length.
While the single accession of L. ornata included in the phylogeny nests within a clade alongside individuals of L. obtusiloba , we accept species status for L. ornata on the basis of morphological and ecological differences from L. obtusiloba ( Engel & Schuster 2001) . New Zealand’s Southern Alps are geologically young as probably are the alpine habitats within which L. ornata grows. Recent divergence and incomplete lineage sorting may explain the nestedness of morphological entities in this instance.
The presence of a well-defined field of large disc cells and deeply divided underleaves with acute lobe apices are distinctive features shared by L. concinna Colenso (1886: 244) and L. bragginsiana . Lepidozia concinna differs from L. bragginsiana in 1) underleaves on main branches symmetrically divided, wider than stem and narrowed to their insertion vs asymmetrically divided, as wide or narrower than the stem, and adnate, 2) lack of accessory armature on the basal margin of the underleaves vs present, 3) lack of accessory armature on postical leaf margin vs present, 4) lack of indistinct tooth on dorsal leaf margin vs usually present, 5) the generally lower leaf disc, 12–19 vs 17–25 cells high 6) pinnate vs bipinnate branching.
The erect habit and presence of ventrally secund branches are shared by Lepidozia obtusiloba and L. bragginsiana . However, L. obtusiloba can be distinguished from L. bragginsiana by its underleaves divided to 0.25 with triangular lobes, the shallow sinus between the first and second leaf lobes, and the apex of the 4 th leaf lobe pointing in the same direction as the other lobes.
Distribution and ecology:— Lepidozia bragginsiana seems to be widespread in hyper-humid forested habitats on the western side of the Southern Alps in New Zealand’s South Island, from Nelson in the north to Fiordland. The species also occurs in the North Island as far north as Mt Te Aroha but does not appear as common there as in the south. The species is probably more widely distributed and may eventually be found in suitable habitats in the central and southern North Island and on Stewart Island/Rakiura. Lepidozia bragginsiana occurs across a wide elevational range, from c. 200 m to 1000 m, and grows in association with both podocarp-broadleaf and beech forests, so long as these are sufficiently wet. Lepidozia bragginsiana occupies raised microsites on the forest floor, such as hummocks and soil-mounds, rotting logs, and the bases of tree trunks. Occasionally it may live on humus covered tree trunks, particularly larger trees where epiphytic bryophyte mats are well-developed. The preference for raised microsites is apparently consistent across the range of this species, Child’s collection (J. Child 2876) from Siberia River was from on old logs in wet beech forest and was recorded as being abundant, the species was epiphytic in forest on Sewell Peak, and at Key Summit grew on an old log. At the type locality Lepidozia bragginsiana grew atop a boulder on the floor of forest with emergent Libocedrus Endlicher (1847: 42) and Dacrydium Lambert (1806: 93) over a 14–18 m tall canopy of Quintinia de Candolle (1830: 5) and Weinmannia racemosa Linné (1781: 227) , on a steep slope with NE aspect. Lepidozia bragginsiana often grows in association with other large Lepidozia species , including L. microphylla ( Hooker 1818: 80) Lindenberg in Gottsche et al. (1845: 202), L. spinossissima (Hooker & Taylor in Taylor 1846: 373) Mitten in Hooker (1854: 146), and L. pendulina , with these species occupying depressions and hollows adjacent to the raised microsites it occupies. A typical pattern is observed at Cross Creek in Haast Pass where L. bragginsiana , L. obtusiloba and L. pendulina all co-occur, the first two occupying raised microsites including hummocks, logs and stumps and tree trunk bases, while the last occupies flat sites and depressions ( Fig. 5 View FIGURE 5 ). Between these microsites the species abut, and intermix across the boundaries between these zones.
Remarks:— In the key to species presented by Engel & Glenny (2008) Lepidozia bragginsiana will usually key to L. pendulina on the basis of its bipinnate shoot systems ( Fig. 2B View FIGURE 2 ). We propose the following couplets as an aid to species identification in conjunction with Volume 1 of the Liverwort Flora of New Zealand:
21b. Leaves conspicuous, wider than stem, patent or appressed to stem, sinus between 3 rd and 4 th lobes divided to 0.65–0.9 .............22.
22a. Sinus between 3 rd and 4 th lobes on leaves from primary shoots wide, 4 th lobe apex pointing postically, at nearly 90° to other leaf lobe apices. Underleaves divided to 0.5 their length......................................................................................................................23.
22b. Sinus between 3 rd and 4 th lobes on leaves from primary shoots narrow, 4 th lobe apex pointing in more or less the same direction as other leaf lobe apices. Underleaves divided to 0.25 their length....................................................................... Lepidozia obtusiloba .
23a. Leaves on primary shoots remote and widely spreading. Accessory armature on postical leaf margin always absent. Underleaves on primary shoots with spreading lobes, lobes narrowly triangular, more than 4 cells broad to near the apex and 10–13 or more cells broad at their base ...................................................................................................................................... Lepidozia pendulina .
23b. Leaves on primary shoots imbricate and appressed. Accessory armature on postical leaf margin often present, particularly on leaves from primary shoots, as irregular spurs or spines. Underleaves on primary shoots with parallel or convergent loans, lobes attenuate, 1–3 cells broad throughout their upper half, and uniseriate for 5–10 cells toward their apex, 4–6 cells broad at their base................................................................................................................................................................ Lepidozia bragginsiana .
Nomenclature: Lepidozia bragginsiana is not known to be represented in the type material of any other Lepidozia species.
Specimens examined:— NEW ZEALAND: orth N Isla Nd, South Auckland Province, Mt Te Aroha , 37°32’S 175°45’E, 900–940 m, 12 Mar 1995, J. J GoogleMaps . Engel 22140, F ; Gisborne Province, Urewera National Park , Panekiri Range, summit of Pukenui in vicinity of Panekiri Bluff , 38°47’S 177°4’E, 1180 m, 24 Mar 1997, J. J GoogleMaps . Engel 23321, F ; Urewera National Park , track to Whatakaka Hut from Lake Waikaremoana , north of western extremity of lake, 38°42’S 177°00’E, 1160 m, 23 Mar 1997, J. J GoogleMaps . Engel 23298, F ; Wellington Province, Ruahine Range, Pohangina Valley , 40°10’S 175°50’E, c. 900 m, 16 Nov 1969, J GoogleMaps . Child H201 , F1034799 ; south Isla Nd : Nelson Province, Mt Duppa , 41°14’S 173°30’E, c. 1100 m, 1 Feb 1981, J GoogleMaps . Child H3389 , F1086726 ; Canterbury Province, Arthurs Pass National Park , near Bealey Glacia Vista , 23 Jan 1971, J. J . Engel 6839, F1104555 ; Arthurs Pass National Park , Bealey Valley Track, 42°55’S 171°33’E, 900 m, 5 Mar 1997, J. J GoogleMaps . Engel 22873, F ; Westland, Grey County, Paparoa Range , cirque on east flank of Mt Priestly , 960 m, 13 Apr 1983, A. J . Fife 5573, F1104682 ; Westland, Paparoa Range, Mt Sewell , 3000 ft, 23 Aug 1975, J . Child H2783 , BM, F1034957 ; Tributary of Siberia River , opposite and downstream of Siberia Hut , 2500 ft, 17 Jan 1976, J . Child 2876, as L. concinna BM ; Key Summit , 3000 ft, 15 Apr 1974, J . Child 2585, BM ; Haast Pass, Cross Creek , 2008, E. A . Brown 08/266, NSW ; Westland Province, Camp Creek west of Alexander Range , 42°42’S 171°33’E, 840 m, Dec 1982, A GoogleMaps . Reif C170 D, F1062778 ; ibid, A . Reif C187 C, F1063049 ; ibid, 1040 m, A . Reif C216 C, F1063054 ; ibid, 1030 m, A . Reif C255 B, F1063060 ; ibid, 1020 m, A . Reif C127 C, F1062799 ; ibid, 980 m, A . Reif C254 B, F1063059 ; ibid, 950 m, A . Reif C214 F, F1063053 ; ibid, 700 m, A . Reif C232 E, F1063055 ; ibid, 585 m, A . Reif C282 D, F1063073 ; ibid, 770 m, A . Reif C249 E, F1062777 ; Westland Province, Arthurs Pass National Park , north of Kellys Creek near highway 73, above campground north of Otira , 42°48’S 171°34’E, 420–475 m, 5 Jan 1983, J. J GoogleMaps . Engel 18353, F1105966 ; south Westland , Lake Ellery , 44°5’S 168°43’E, c 60 m, 12 Dec 1981, J GoogleMaps . Child H4574 , F1088411 ; Southland Province, Fiordland National Park , Moraine Creek track, area north of Morain Creek , west of Hollyford River , 44°44’S 168°5’E, 610 m, 16 Mar 1997, J. J GoogleMaps . Engel 23218, F ; Fiordland National Park , off track along East Branch of Eglinton River , SE of Mt Eglinton, 45°04’S 168°01’E, 440 m, 20 Jan 1983, J. J GoogleMaps . Engel 18760, F1105967 ; Fiordland , between Te Anau and Homer Tunnel, 9 Feb 1955, R. E . Hatcher 1560, F1106026 ; Southland Province, Cascade Creek , near Lake Gunn , tributary of Eglinton River , 44°55’S 168°6’E, c. 400 m, 14 Apr 1974, J GoogleMaps . Child H2591 , F1034966 ; ibid, J . Child H2598 , F1034987 ; Fiordland, head of Lake McKerrow , 24 Jan 1955, R. E . Hatcher 828, F1106022 ; Lake Manapouri , south arm, 20 Apr 1970, J . Child H251 , F1034645 ; Otago Province, Beans Burn, tributary of Dart River , 44°35’S 168°15’E, 400 m, 23 Aug 1972, J GoogleMaps . Child s.n., F1033346 ; Turret Peak, Earnslaw , 44°38’S 168°23’E, c. 600 m, 19 Aug 1973, J GoogleMaps . Child H2469 c, F1033545 ; tributary of Siberia Stream, opposite and downstream from Siberia Hut , 44°13’S 169°3’E, c. 800 m, 17 Jan 1976, J GoogleMaps . Child H2892 , F1034636 GoogleMaps .



| M |
Botanische Staatssammlung München |
| A |
Harvard University - Arnold Arboretum |
| AK |
Auckland War Memorial Museum |
| NSW |
Royal Botanic Gardens, National Herbarium of New South Wales |
| F |
Field Museum of Natural History, Botany Department |
| N |
Nanjing University |
| J |
University of the Witwatersrand |
| BM |
Bristol Museum |
| E |
Royal Botanic Garden Edinburgh |
| C |
University of Copenhagen |
| B |
Botanischer Garten und Botanisches Museum Berlin-Dahlem, Zentraleinrichtung der Freien Universitaet |
| R |
Departamento de Geologia, Universidad de Chile |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |