
Cervonema indet
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3368.1.14 |
|
persistent identifier |
https://treatment.plazi.org/id/B420BD55-FFD5-FF97-3CA7-119FFD6166B2 |
|
treatment provided by |
Felipe |
|
scientific name |
Cervonema indet |
| status |
|
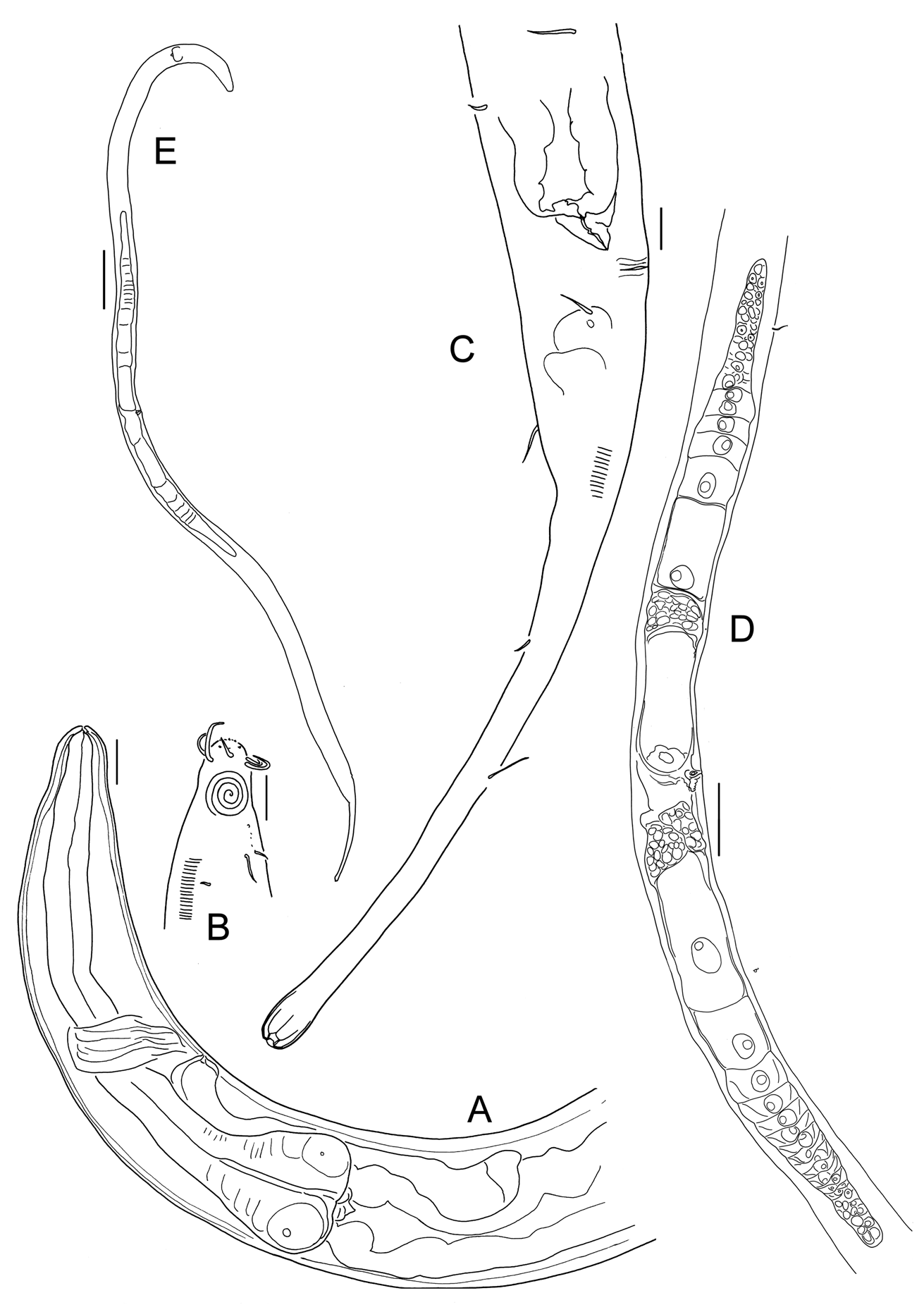
Cervonema indet . B
( Fig. 8 View FIGURE 8 )
Material. One female, NIBRIV0000245890. Collected by Jinwook Back in May 2007 from Research Vessel Tam
Gu 18.
Sampling locality. Subtidal, 25 m, in silty sand off the Korean west coast (36.251º N 126.198º E), within the chain of islands from Sapsi-do to Eoncheong-do.
Measurements. See Tables 7 and 8 for detailed measurements and calculations.
3 138 298 792 295 1534 1727
female 1 10 33 40 31
Description. Female ( Fig. 8 View FIGURE 8 ). Habitus tubular, tapering from cloaca to conico-cylindrical tail posteriorly, anterior narrowing more gently from nerve ring to lips. Cuticle with faint, relatively fine striation throughout body, cylindrical section of tail appears smooth.
Six small lips indistinct, labial papillae minute. Six anterior cephalic setae (0.7 hd long) located about 0.3 hd from anterior extremity, 4 posterior cephalic seta. (0.9 hd long) in distinctly separate crown. Amphid, 0.5 hd from anterior, multi-spiralled with 4 turns (diameter 0.8 cbd). Cervical setae in two sub-lateral rows, 7 µm long, somatic setae scarce, 9 µm long. Setae scattered around lateral field in cloacal region, last setae observed 3.7 abd (60 % of tail length) from the cloaca in cylindrical portion of tail.
Anterior section of buccal cavity minute (2 µm deep), posterior section not observed. Oesophageal musculature surrounds buccal cavity, sclerotization not observed. Posterior oesophageal bulb (20 % of oesophagus length), distinct rounded triangular shape; flattened posterior base. Cardia short (3 µm long). Nerve-ring at 54 % oesophageal length from anterior, ventral pore posterior to nerve ring (65 % of oesophageal length). Ventral gland small, round, behind cardia, distance from pore to base of gland 85 µm (62 % of oesophageal length).
Vulva at 46 % of body length, as lateral slit without epiptygmata. Vagina short and straight without sclerotizations. Didelphic, outstretched ovaries, anterior 298 µm, posterior 295 µm long. Supplements absent. Tail conico-cylindrical 6.3 abd long, with cylindrical portion 74 % of total, three terminal setae (obscured).
Affinities. Cervonema sp. B is similar to C. chilensis and C. hermani in having the amphids positioned almost directly behind the cephalic setae. It is additionally similar to H. hermani owing to the relative lengths of the cephalic setae (as hd) and the tail (as abd). It differs from C. hermani and D. chilensis , however, with regard to the de Man's ratio a (43 vs 60 and 35, respectively) and de Man's ratio b (12.5 vs 9.9 and 8.5, respectively). It is additionally different from C. hermani and C. chilensis with regard to the R2:R3 sensilla length ratio (0.8 vs 1 and 1, respectively), and in having fewer turns of the amphid (4 vs 5.5 and 5.5, respectively).
Concluding remarks on Cervonema . Like Laimella , species of Cervonema are recorded from the intertidal and estuarine habitats to the deep sea, and mostly recorded in fine sand and muddy sediments. It has also been noted that they tend to be restricted to marine or near-marine salinities ( Soetaert et al. 1995). Forster (1998) noted that an upper-shore species of Cervonema was able to osmoregulate under osmotic stress conditions (immersion in hypo- or hyper-tonic solutions), though of course this will reflect habitat-specific adaptation across the Nematoda rather than a species or genus specific characteristic.
Cervonema have been described from Africa, America, Asia and Europe, and have been recorded in, but not described to, the Arctic and Southern Oceans ( Vanreusel et al. 2000; Lee et al. 2001; respectively). They may be (co-)dominant in some habitats (eg. upper and mid-slope; see Muthumbi et al. 2004; Ingels et al. 2011) and deepsea manganese nodule provinces ( Miljutina et al. 2010; Vopel & Thiel 2001) and have been observed to increase in abundance in disturbed sediments ( Lee et al. 2001).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.