
Paralimnadia laharum, Brian V Timms, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4418.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:28B3866D-89EF-4CB6-8152-E622B253CA9E |
|
DOI |
https://doi.org/10.5281/zenodo.5969159 |
|
persistent identifier |
https://treatment.plazi.org/id/B446878B-2F69-592E-28C2-FF6CFD00BAF8 |
|
treatment provided by |
Plazi |
|
scientific name |
Paralimnadia laharum |
| status |
|
Paralimnadia Sars 1896 View in CoL emend Rogers et al., 2012
Paralimnadia laharum sp. nov. (Figs. 1,3,4)
Etymology. The specific epithet is based on the local aboriginal words Lah Arum applied to the large sandstone rock formations of what is known in European times as the Grampians. Specifically Lah Arum means a place of large stones. The derived word ‘laharum’ is used in the sense that this clam shrimp occurs on the rocks of the Grampians, specifically in bare rock pools.
Type locality. Victoria, Grampians ranges, pool (the deepest ‘ Pudding Basin’ shaped pool, on Flat Rock , 36o 53’ 43.6”S, 142o 22’ 37.7”E, GoogleMaps
Type material. Holotype. Male, length 8.6 mm, height 5.5 mm (NMV J73116 View Materials ). Coll. W. Gardner, 2 July 2017,
Allotype. Female, length 9.6 mm, height 5.3 mm, (NMV J73117 View Materials ) collected with holotype.
Paratypes. 2 males, lengths 7.9 and 7.9 mm, 2 females, lengths 8.0 and 8.0mm, Coll. BVT, ( NMV J73118 View Materials ). Victoria, Grampians, pool on Mt Stapylton , 36o 53’ 56.7”S, 142o 23’ 14.5”E, 17 June 2017 GoogleMaps . 2 males, lengths 7.9 and 7.6 mm, 2 females, lengths 7.9 and 7.8 mm, Coll. BVT, (AM P 101152). Victoria, Grampians, pool on Mt Stapylton , 36o 53’ 56.7”S, 142o 23’ 14.5”E, 17 June 2017. GoogleMaps
Other material. Victoria. 2 males, 2 females ( NMV J68585 View Materials ), Grampians, pool on Mt Difficult, 30 May 1973, no collector recorded . 5 males, 5 females Coll. BVT, (NMV J73119 View Materials ), Grampians, pool on Mt Stapylton, 36o 53’ 56.7”S, 142o 23’ 14.5”E, 17 June 2017. 5 males, 5 females Coll. BVT, ( NMV J731210), Grampians, pool on Hollow Mountain , 36o 53’ 31.3”E, 142o 22’ 53.3”E, 18 June, 2017 . 5 males, 5 females Coll. BVT, (NMV J73121 View Materials ), Grampians Visor pool on Flat Rock, 36o 53’ 41.3”E, 142o 22’ 37.2”E, 18 June 2017. 5 males, 5 females, Coll. BVT, (NMV J73122 View Materials ), Grampians, Pudding Basin pool on Flat Rock, 36o 53’ 43.6”S, 142o 22’ 37.7”E, coll W. Gardner 14 August 2017. 5 males, 3 females (NMV J73123 View Materials ) Grampians, Pudding Basin pool on Flat Rock, 36o 53’ 43.6”S, 142o 22’ 37.7”E, coll W. Gardner, 1 October 2017.
Diagnosis. Egg spherical covered with about 58 irregular polygons each with a central groove. First antenna with about 9 lobes in males and about 5 in females. Second antenna with 11–12 antennomeres. Clasper with both long palps biarticulated and inerm. Telson with about 19 spines, and cercopod with 4–5 setae of moderate length and spaced on basal 50%.
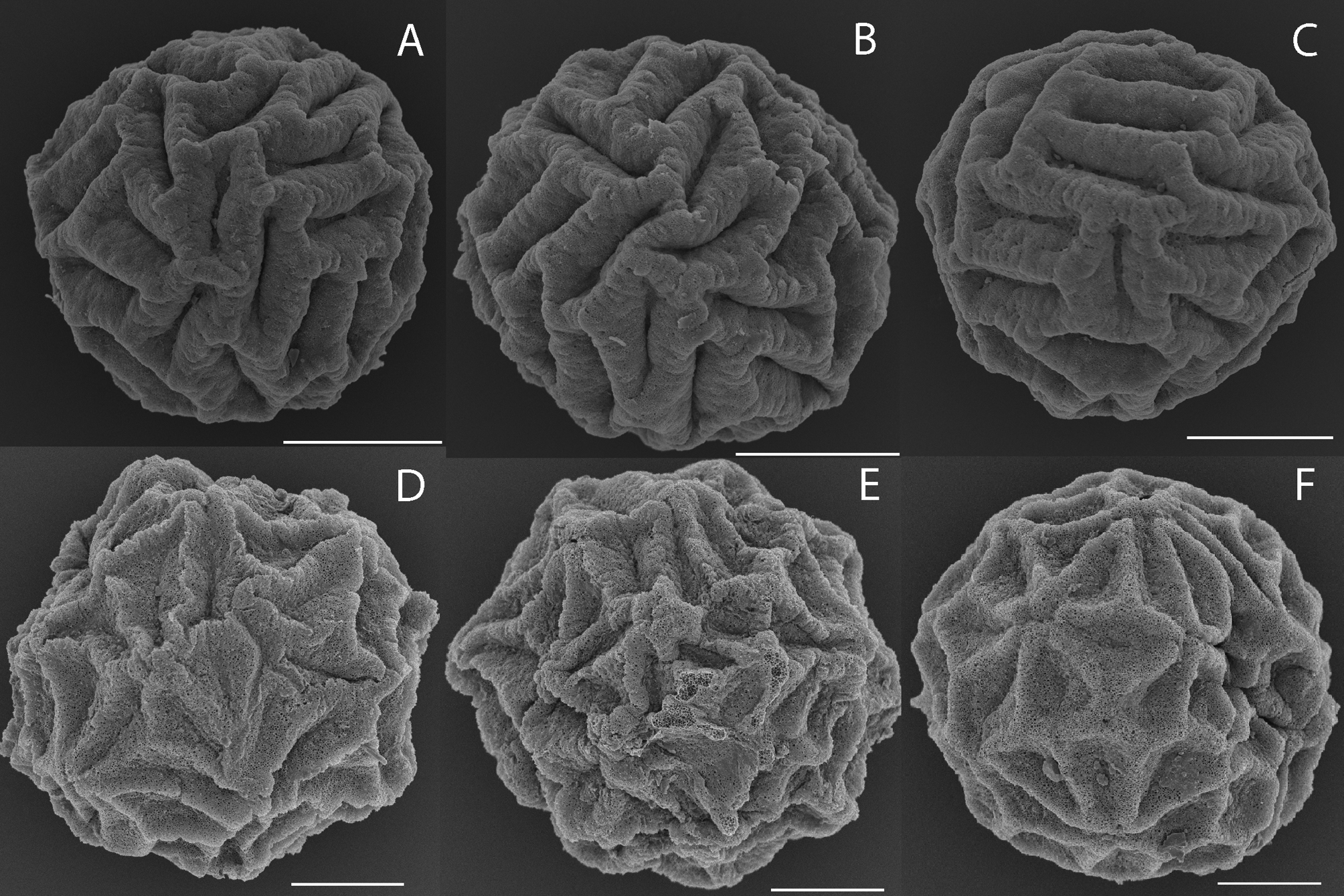
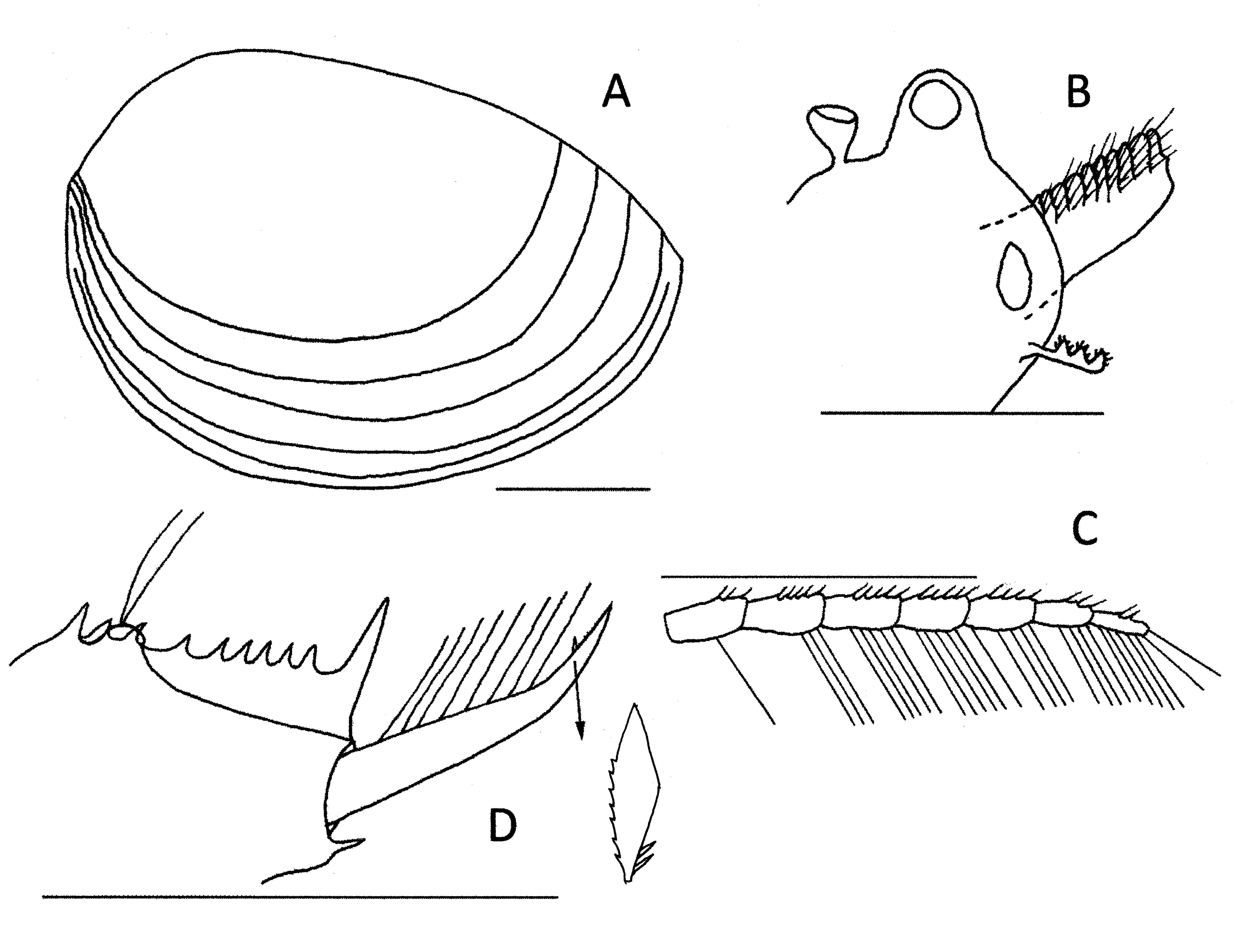
Description. Egg ( Fig. 1D,E,F View FIGURE 1 ). Mean diameter 412 µm, range 388 to 426 µm, n =10. Spherical with surface divided into about 58 (range 52–62) irregular polygons, with thick walls and a deep central groove 12–30 µm long and a sloping shoulder region in between. Perimeter walls generally rounded, but can have a narrow frilly apex. Tertiary layer spongiform.
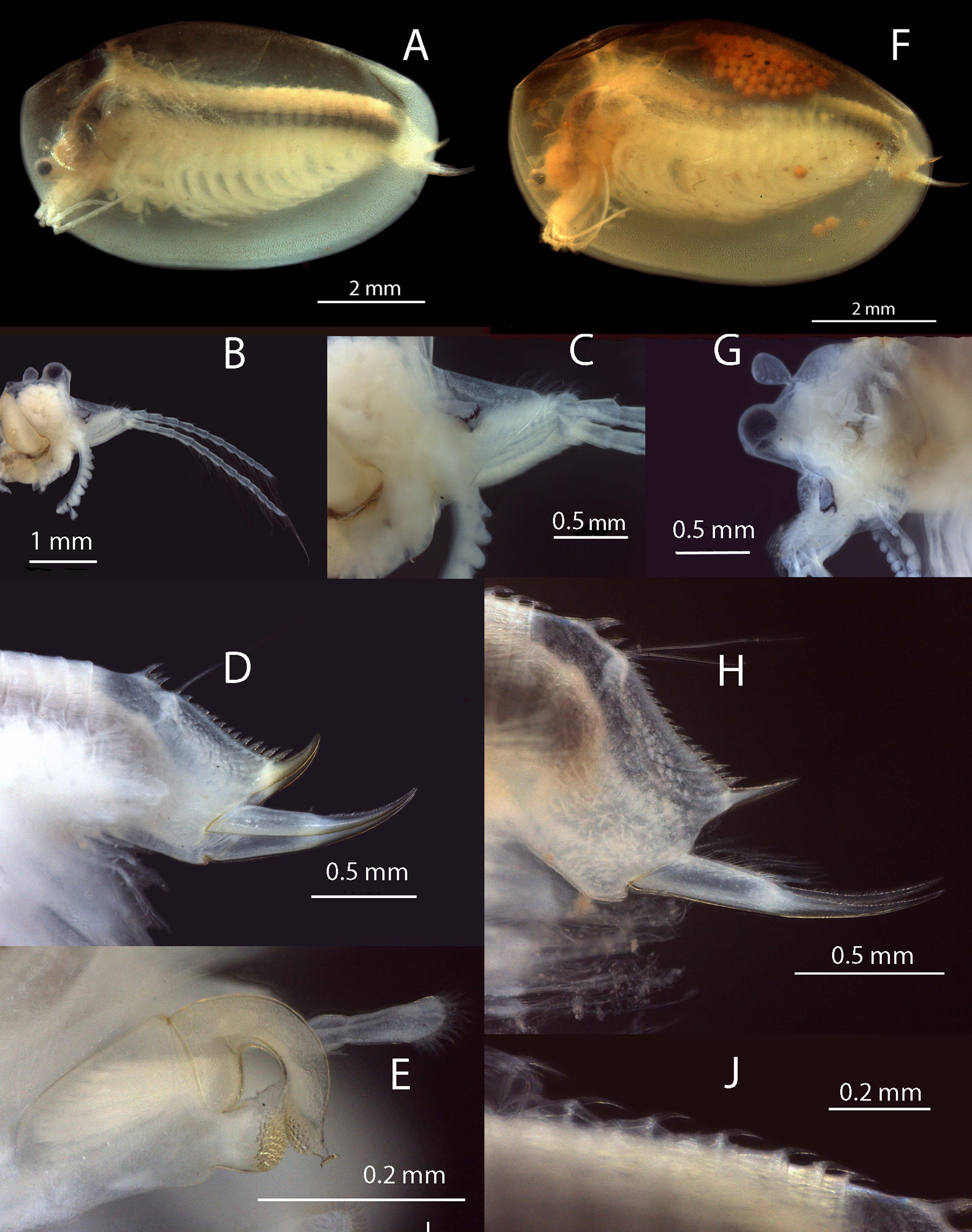
Male. Head ( Fig 3C View FIGURE3 , 4B View FIGURE 4 ) with ocular tubercle prominent, the compound eye occupying most of it. Rostrum protruding a little more than the ocular tubercle, asymmetrically triangular with a rounded apex and angle between it and frons about 80o. Ocellus rounded and a little smaller than compound eye and located dorsobasally in rostrum. Dorsal organ posterior to eye by about one third of its height, pedunculated and almost symmetrical and not quite as high as ocular tubercle.
First antenna ( Fig. 3C View FIGURE3 , 4B View FIGURE 4 ) a little longer than peduncle of second antenna and with about nine lobes, each with numerous sensory setulae. Second antenna ( Fig. 3E View FIGURE3 , 4B View FIGURE 4 ) with spinose peduncle and both flagella usually with 12 antennomeres, dorsally with 1–3 short spines per antennomere and ventrally with 1–5 longer setae (average of 3 spines and 3 setae on middle antennomeres).
Carapace ( Fig. 3A View FIGURE3 , 4A View FIGURE 4 ) elongated oval with a distinct anteriodorsal angle and an indistinct ventrodorsal angle. Dorsal edge evenly convexly curved and usually darkly coloured, otherwise pellucid. A few indistinct growth lines.
Eighteen pairs of thoracopods. First two modified as claspers ( Fig. 3D View FIGURE3 , 4E View FIGURE 4 ), each with palm trapezoidal, with a distinct angle distomedially and subterminal to the apical club bearing many stout spines directed medially. Small palp with many short thin limp spines apically. Finger arcuate with a rounded apex with a suctorial organ distodorsally and many rounded pits distoventrally. Long palps of claspers I and II inserted on apical edge of palm, each with two palpomeres and inerm palpomere junctions. First palp subequal in length to palm, and second palp a little longer. Other thoracopods of typical structure for Paralimnadia (see Fig 2 View FIGURE2 , Timms, 2016b), decreasing in size and complexity posteriorly. Dorsally segments IX or X to XVII with a single posterior spine, but segment XVIII inerm.
Telson ( Fig. 3B View FIGURE3 , 4D View FIGURE 4 ) with spine rows bearing about 18 pairs of similarly sized and evenly spaced spines except the first twice the size of the remainder and the spiniform projection about 5 x the average size. This projection with noticeable spinulae, the spines almost smooth. Telsonic filaments originating from a distinct mound in the dorsal floor of the telson, usually not protruding above the spine row and positioned between the 4th and 5th spine pair. Steep declivity posterior to mound with remainder of floor slightly convex. Cercopod stout basally and thinning markedly at about 50% of its length to a thin sharp apex. About 5 spaced setae, length up to twice cercopod basal diameter, on basal half demarcated by a short spine. Distal half with a cirrus of denticles. Ventroposterior corner of telson with a triangular extrusion not extended into a spine.
Female. Head ( Fig 3H View FIGURE3 , 4G View FIGURE 4 ) with ocular tubercle prominent, the compound eye occupying most of it. Rostrum protruding a little less than the ocular tubercle and symmetrically triangular. Angle between frons and rostrum quite obtuse, about 135o. Ocellus rounded about half the size of the compound eye and located basodorsally in rostrum. Dorsal organ posterior to eye by about one half of its height, pedunculated and almost symmetrical and not quite as high as ocular tubercle.
First antennae ( Fig. 3H View FIGURE3 ) a little shorter than peduncle of second antenna and with about 5 lobes each with numerous sensory setules. Second antenna as in males.
Carapace ( Fig 3F View FIGURE3 , 4F View FIGURE 4 ) as in males though a little more vaulted dorsally. Also dorsal edge often more pigmented, otherwise carapace usually pellucid, but dorsally carapace somewhat darker. No apparent growth lines in most specimens.
Eighteen pairs of thoracopods of typical Paralimnadia structure. Trunk dorsum with segments I to about VIII and XVIII inerm, segments about IX to XII with 5 or so setae, segments XIII to XIV with about 3 setae, segments XV to XVII with 1 seta each ( Fig 4J View FIGURE 4 ) Large egg retaining epipods on thoracopods IX and X.
Telson ( Fig 3G View FIGURE3 , 4H View FIGURE 4 ) largely as in male, though the 19 pairs of spines more uneven in size and placement and telsonic filaments inserted between 5th and 6th spine pairs. Cercopod as in male, with spine mid length and about 4 cercopod setae. Ventroposterior corner triangular.
Variability. Some populations have dark dorsal pigments and otherwise are somewhat opaque. Whether pellucid or opaque, animals easily blend in the background and hence are well camouflaged. Growth lines are usually invisible, but in some specimens 4–5 spaced lines are visible. Antennal spines, setae and lobes as well as telsonic spines and setae can number ± 1 of the numbers recorded for the types.
Differential diagnosis. The egg is most similar to that of Paralimnadia sordida ( Fig. 5F–H View FIGURE5 , Timms, 2016b), but its polygons are cup shaped in profile without a central groove, and the ends of some adjacent polygons tend to protrude as short blunt spines. Moreover there are many differences in morphology between the two species, e.g.: (i) there are more telsonic spines in P. sordida (22) than in P. laharum sp. nov. (19), many more cercopod setae in P. sordida (ca 12) than P. laharum sp. nov. (5), and (iii) the long palps of the claspers are triarticuate with spines at the basal joint in P. sordida but biarticulate and inerm in P. laharum sp. nov..
An analysis of the characteristics of the 15 described species of Paralimnadia (Tables 2 and 3 of Timms, 2016b) reveals significant differences between P. laharum sp. nov. and all of them. Given the number and length of cercopod setae are a key distinguishing character ( Timms, 2016b), there are four possible similar species: P. stanleyana , P. urukhai , P. multispinosa and P. westraliensis ; though all four have 6 or more cercopod setae and P. laharum sp. nov. has 4–5. P. stanleyana and P. urukhai are ruled out as they have <15 telsonic spines and <7 male first antennal lobes whereas P. laharum sp. nov. has ca 19 telsonic spines and 9 male first antennal lobes. Some P. multispinosa and P. westraliensis individuals share these characters with P. laharum sp. nov., but any possible synonymy is rejected based on cercopod setae being short (ca 1 x cercopod diameter) and none near the midway spine whereas in P. laharum sp. nov. the cercopod setae are 1.5 to 2 x cercopod diameter and they are spaced evenly on the basal half of the cercopod. These four species also have very different egg morphologies to that in P. laharum sp. nov. (cf Figs 5A–E, P View FIGURE5 . stanleyana; Fig 20A, P. multispinosa ; Fig 10I –L, P. westralensis with Fig. 1 View FIGURE 1 herewith).
Distribution and ecology. This species is known only from a few rock pools in the higher parts of the Grampians Mountains in Western Victoria. Their eggs hatch after the first major rains of the wet season, usually in May–June, and grow to adulthood over the next few weeks and then persist generally for only a few weeks more, but in the deeper pools (e.g., the one on Mt Staplyton, The Pudding Basin pool on Flat Rock) populations may last months (W. Gardner, pers. comm.). In the Flat Rock Pool the population studied first on 17 June 2017 persisted to at least 1 October 2017 with no significant further increase in size of the individuals after July. Many pools have sparse populations, possibly because strong winds in the dry season blow sediments away so that many pools have bare rocky bottoms. Also it seems there are few other inhabitants (mainly mosquitoes, a few beetles and bugs and very few ostracods and cladocerans) of these pools compared to pan gnammas on Eyre Peninsula to the west ( Timms, 2014) or central Victoria to the east ( Timms, 2017).


| NMV |
Museum Victoria |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |