
Eulimnadia kimberleyensis, Brian V Timms, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4418.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:28B3866D-89EF-4CB6-8152-E622B253CA9E |
|
DOI |
https://doi.org/10.5281/zenodo.5969155 |
|
persistent identifier |
https://treatment.plazi.org/id/B446878B-2F6C-592A-28C2-FC85FD11B876 |
|
treatment provided by |
Plazi |
|
scientific name |
Eulimnadia kimberleyensis |
| status |
sp. nov. |
Eulimnadia kimberleyensis View in CoL sp. nov. ( Figs 1 View FIGURE 1 , 2 View FIGURE2 ).
Etymology. This species is named for the area of Australia in which it occurs.
Type material. Holotype. Female. Rock pool. Western Australia, Kimberley , Gardner Plateau, from 14o 47’ 2.1”S, 126o 31’ 18.9”E. Length 4 mm, height 3.1 mm ( WAM C72086), coll A. Cross, March 2012. GoogleMaps
Paratypes. Two females. Lengths 4.0 and 3.9 mm ( WAM C72087) from the same locality.
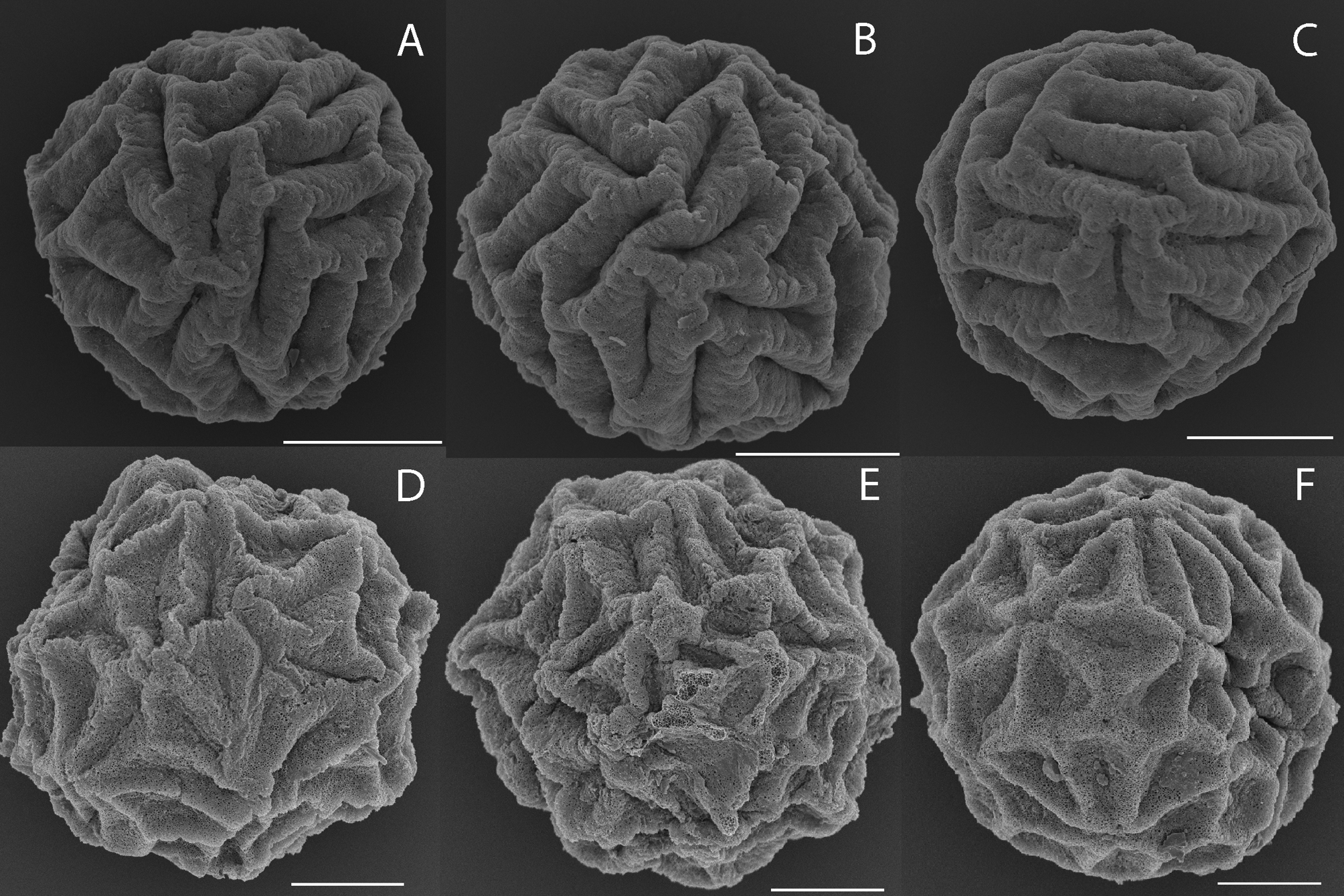
Diagnosis. Egg spherical with about 36 grooves arranged randomly and about 30 µm long; walls of grooves thick and rounded with surface usually crenulated at right angles to groove axis. Female first antennae with about three lobes; second antennae with 7 antennomeres and generally 4–5 spines dorsally. Telson with about 19 dorsal spines and cercopod with about 8 long setae.
Description. Egg ( Fig 1A, B, C View FIGURE 1 ). Spherical, mean diameter 128 µm, range 122 to 134 µm, n = 10. Surface with about 36 (range 33–42, n = 10) grooves each about 30 µm long and arranged randomly. Tertiary layer represented by the apparent groove walls spongiform and thick and rounded, though surface usually crenulated at right angles to the major axis; sometimes a minor elevated area where walls of adjacent grooves confluent. In cross section, each groove—wall system consists of 80–90% wall and only 10–20% flat-floored groove with a distinct demarcation between the two.
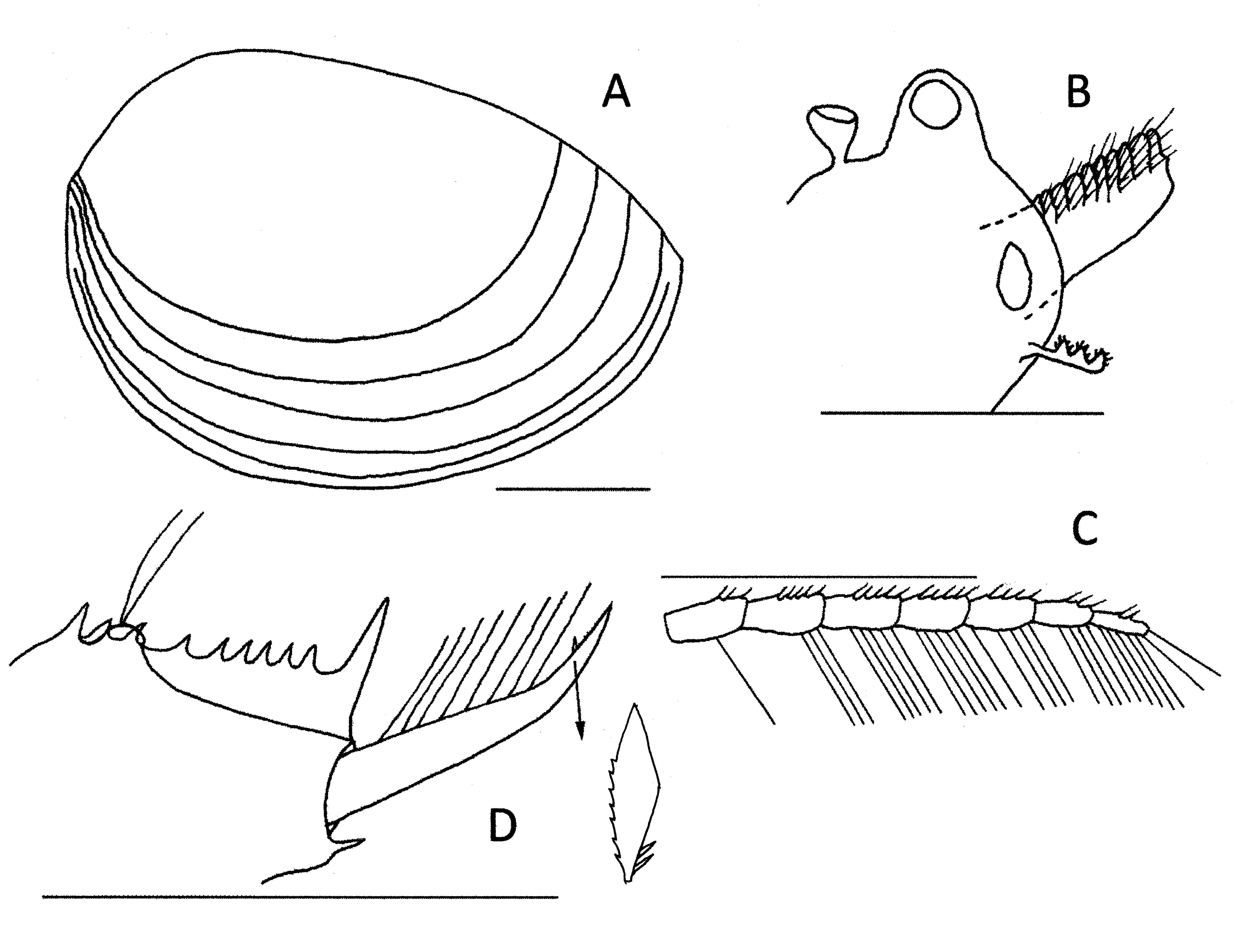
Female. Head ( Fig. 2B View FIGURE2 ) with ocular tubercle prominent, the compound eye occupying most (ca 80%) of it. Rostrum broadly rounded and protruding a little less than the ocular tubercle. Ocellus about as large as the compound eye and located in the middle base of the rostrum. Frons-rostrum angle about 150o. Dorsal organ posterior to the eye by about its height, pedunculated, asymmetrical and about half as high as the ocular tubercle.
First antenna ( Fig 2B View FIGURE2 ) short, about 40% the length of the peduncle of the second antenna, and with three lobes, each with short sensory fine setae.
Second antenna ( Fig 2C View FIGURE2 ) with 7 antennomeres, most (II to VI) with 4–5 short spines dorsally and 3–5 long setae ventrally. Basal antennomere with just 3 spines and a seta while distal antennomere with 2 spines and about seven setae.
Carapace ( Fig 2A View FIGURE2 ) elongated oval with a vaulted dorsal and ventral edge and weak anterodorsal and posterodorsal angles. Abductor muscle scar at about 45o to carapace long axis. Carapace pellucid and without markings except for five growth lines, generally well spaced except near the ventral edge.
Trunk of 18 segments. Dorsal surface of posteriormost 9–10 segments with many long two-segmented setae and sometimes shorter spines. Thoracopods of trunk segments IX and X with long epipods for retaining eggs.
Telson ( Fig 2D View FIGURE2 ) with 9 dorsal spines followed by a large spiniform projection, the first spine about 20% larger than the next few followed by some variation in size and spacing along the row. Spines smooth. Telsonic filaments inserted on a mound between 2nd and 3rd spine. A moderate declivity posterior to the mound, remainder of telsonic floor sloping evenly and gently to cercopod base. Cercopod a little shorter than telson length and with a short spine about 80% along its length separating a thicker basal area supporting setae from a thinning distal area with a weak cirrus of small denticles. Short laminate spine with about 7 serrations on anterior surface and two small spines near the base. (see Fig. 2D View FIGURE2 insert). Eight long cercopod setae, their length about twice the basal diameter of the cercopod. Prominent spiniform projection on ventroposterior corner of telson.
Variability. There are only three specimens available, so little can be added on variability of the above characters. Sometimes there are only six antennomeres though whether this is natural or damaged is not certain. Telsonic spines may number 8–10 and and cercopod setae 7–10.
Differential diagnosis. There is no doubt this species belongs to the genus Eulimnadia . The diagnostic character is a spiniform projection on the ventroposterior corner of the telson ( Rogers et al., 2012). In addition most Eulimnadia , including this species, have a spine on the cercopod at about 4/5ths its length and the antenna has about 7–8 antennomeres on each flagellum ( Timms, 2016a).
However, because of the limited number of specimens and lack of males the description of this species is minimal. Nevertheless species specific egg morphology in this genus ( Belk, 1989; Rabet, 2010; Timms, 2016a) provides characters to separate species and in some cases is about the only distinguishing character, e.g. E. graniticola Rogers et al. 2010 ). However, for Australian species species specific egg morphology has been supported by features of the cercopods and sometimes a few other characters as well from the telson, claspers, antenna and head ( Timms, 2016a). This dictum applies to this species.
In this case the egg morphology is certainly the most distinguishing feature, though a combination of less important characters (a cercopod with 8 uniform setae, just nine unevenly spaced telsonic spines and a first antenna of just three lobes) help to define it. The most similar eggs are of E. beverleyae Timms, 2016 and E. uluruensis Timms, 2016 . Eggs of E. beverleyae ( Fig. 3F View FIGURE3 , Timms 2016a) have about 30 grooves, each about 40 µm long, and of the same tertiary material as their walls, whereas in E. kimberleyensis sp. nov. there are but 40 grooves, 40 µm long and of different tertiary material to that of the walls. In addition E. beverleyae has 11–13 cercopod setae whereas E. kimberleyensis sp. nov. has only about 8 cercopod setae. In E. uluruensis (Fig.9 H. Timms, 2016a) the grooves number about 28, lie within distinct polygons with low lateral ridges and a knobbly surface, a situation quite different to that in E. kimberleyensis sp. nov. (see above). Though E. uluruensis has telsonic spines reminiscent of E. kimberleyenesis sp. nov., the cercopod setae of E. uluruensis are shorter than those of E. kimberleyensis sp. nov.
Given E. kimberleyensis sp. nov. lies within the distribution of the widespread and variable E. dahli Sars 1896 , it is appropriate to detail their distinctiveness. Firstly, habitat choice is not distinctive— E. dahli occurs in gnammas in limestone in northwest Queensland ( Timms, 2016a) and also in granite in the Pilbara (author unpublished data) while E. kimberleyensis sp. nov. occurs in sandstone gnammas in between these areas. However, their eggs are entirely different, those of E. dahli have ca. 20 surface polygons, while those of E. kimberleyensis sp. nov. have ca.36 grooves. Eulimnadia dahli typically (but not always) has 20 trunk segments, and E. kimberleyensis sp. nov. the standard 18. Furthermore E. dahli has ca. 15 telsonic spines and ca 18 cercopod setae, whereas E. kimberleyensis sp. nov. has ca 9 and 8 respectively, a difference wide enough to be certain of identity.
Distribution. Known only from gnammas on the Gardner Plateau, Kimberley. These pools fill during the wet season, November to April, and the deeper pools (averaging 46 mm) support aquatic plants ( Cross et al., 2015a, 2015b). No information is available on their fauna, except that a few pools support the new species of clam shrimp described here plus Limnadopsis multilineata Timms 2009 .

| WAM |
Western Australian Museum |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |