
Homo luzonensis, Détroit & Mijares & Corny & Daver & Zanolli & Dizon & Robles & Grün & Piper, 2019
|
publication ID |
https://doi.org/10.1038/s41586-019-1067-9 |
|
DOI |
https://doi.org/10.5281/zenodo.5931871 |
|
persistent identifier |
https://treatment.plazi.org/id/B551AE7B-841A-F701-FE9A-3713FEE4CD1D |
|
treatment provided by |
Admin |
|
scientific name |
Homo luzonensis |
| status |
sp. nov. |
Homo luzonensis sp. nov.
Etymology. The species name is derived from the island of Luzon, where the specimens were discovered.
Holotype. CCH6 (CCH6-a to CCH6-e), maxillary right postcanine dentition of a single individual discovered on 24 August 2011. The repository is the National Museum of the Philippines, Manila. Homo luzonensis has been deposited in the ZooBank database (http://zoobank.org/) with Life Science Identifier urn:lsid:zoobank.org:act:.
Paratypes. Recovered in 2007, 2011 and 2015 from the same excavation area and layer as the holotype: CCH1, a right third metatarsal 1; CCH2 and CCH5, two manual phalanges; CCH3 and CCH4, two pedal phalanges; CCH8, a left upper third or fourth premolar (P 3/4); and CCH9, a right M 3 (all specimens are housed at the National Museum of the Philippines, Manila) .
Referred material. CCH7, a femoral shaft that belonged to a juvenile individual (housed at the National Museum of the Philippines, Manila).
Locality. The type locality is Callao Cave, in the Callao Limestone formation in the Peñablanca region of northern Luzon , the Philippines, at coordinates 17° 42′ 11.7″ N, 121° 49′ 25.5″ E. GoogleMaps
Diagnosis. Postcanine maxillary teeth of small size that are mesiodistally compressed, with a premolar:molar crown size ratio that is high compared to other species in the genus Homo . Upper premolars with two or three roots, a mesio-distally expanded lingual crown, strong buccal grooves, partial or continuous transverse crest, and an enameldentine junction (EDJ) shape that is distinct from that of H. sapiens, Homo neanderthalensis and Asian Homo erectus . Very small upper molars, with a M 1 > M 2 > M 3 crown size pattern, a simplified occlusal morphology with reduced metacone and hypocone, no crenulation on the EDJ, and EDJ shape affinities with that of H. sapiens and Asian H. erectus. Intermediate manual phalanx (rays 2–4) that is long and narrow (unlike all hominins except H. sapiens), with a longitudinally curved and dorso-palmarly compressed shaft, well-developed flexor sheath attachments and a strongly developed dorsal beak; it shares shape affinities with Australopithecus , H. floresiensis and—to a lesser extent— H. sapiens . Distal hand phalanx with proportions unlike
those of early Homo / Paranthropus , Homo naledi, H. floresiensis and H. neanderthalensis. The third metatarsal has a base that is very small relative to bone length, dorso-plantarly short and with a marked dorsoplantar convexity of the proximal articular facet. The proximal pedal phalanx (rays 2–4) shares shape affinities with Australopithecus : marked longitudinal curvature, parallel medial and lateral sides in dorsal view, a small bicondylar head and weak dorsal canting of the proximal articular surface (unlike African and European early Homo , H. naledi, H. neanderthalensis, H. floresiensis and H. sapiens).
Description and comparison of the fossil elements
Of the seven posterior maxillary teeth, six are from the right side (CCH6-a to CCH6-e and CCH9) and one (CCH8) is from the left ( Fig. 2a, f, g View Fig , Extended Data Fig. 1 View Fig and Supplementary Information). The M 2 (CCH6-b) and one of the M 3 s (CCH9) are complete; the other teeth have well-preserved crowns, and at least one—but sometimes all—roots are partly broken. Micro-fissures are present in the enamel, dentine and cementum, and the pulp cavity is generally filled by calcium carbonate; however, these post-depositional modifications do not affect the size, shape or morphological attributes of the teeth. In comparison to Australopithecus , Paranthropus and other species of the genus Homo , the maxillary postcanine teeth of H. luzonensis are small and mesio-distally compressed, and there is a marked contrast between the relative size and shape of the premolars and molars ( Figs. 2–4 View Fig View Fig View Fig and Extended Data Figs. 1–4 View Fig View Fig View Fig View Fig ).
The upper molars of H. luzonensis have small crown sizes with a M 1 > M 2 > M 3 pattern and simplified occlusal surface morphology with a reduced number of cusps and an absence of marked crenulations at the EDJ ( Fig. 3 View Fig and Extended Data Fig. 1 View Fig ). These derived features are found in other Late Pleistocene species of the genus Homo (that is, H. neanderthalensis, H. floresiensis and H. sapiens), but in this respect H. luzonensis molars more closely resemble those of H. sapiens 4, 5. However, the M 1 and M 2 of H. luzonensis have smaller crown sizes than those of H. sapiens ( Fig. 3 View Fig and Extended Data Fig. 2d, e View Fig ). Compared to the molars of H. floresiensis, H. luzonensis molars are smaller and the M 1 is not more mesio-distally compressed than the other postcanine teeth 6, 7 (Extended Data Figs. 2–4 View Fig View Fig View Fig ). For their diminutive size and simplified crown morphology, H. luzonensis molars also differ considerably from Asian H. erectus 4, 5, 8, 9 and Denisovans 10, although the EDJ of H. luzonensis M 1 shows some shape affinities with several Indonesian H. erectus specimens (Extended Data Fig. 4f, h View Fig ).
H. luzonensis premolars are noticeably large relative to the molarseven more so than seen in H. floresiensis and unlike the pattern seen in other hominins, except Paranthropus ( Fig. 3 View Fig )—and also display several primitive features. For instance, the P 3 and P 4 crowns are asymmetric with a large and mesially displaced lingual cusp, as in early Homo 11, and complete or partial mesial transverse ridges are present on the upper premolars, similar to those of H. floresiensis 6, 7. Developed mesial and distal vertical grooves are also evident on the buccal aspects of the P 3 and P 4; features that are more frequent in early Homo than in H. sapiens 12 and that are absent in H. floresiensis 6, 7. Moreover, H. luzonensis premolars have multiple roots (P 3 has three, P 4 has two) that are robust and highly divergent, an archaic condition typically found in Australopithecus , Paranthropus and early Homo 13. Multiple robust and divergent premolar roots also occur in Asian H. erectus (Early Pleistocene Indonesian and Middle Pleistocene Chinese specimens), but are rare in H. sapiens and absent in H. floresiensis 4, 6, 9, 12 – 17. CCH8 ( Fig. 2f View Fig and Extended Data Fig. 1f View Fig ), an isolated left upper premolar, exhibits three roots (typically a P 3 feature) with a relatively symmetric crown (typically a P 4 feature). If CCH8 is a P 4, a similar three-rooted condition has not been reported for any Pleistocene Homo from Asia and is rarely found in modern humans (1 to 3%) 13, 14.
In terms of absolute tooth size and premolar–molar proportions, H. luzonensis shows a pattern that is not seen elsewhere in the genus Homo ( Fig. 3 View Fig and Extended Data Fig. 2 View Fig ). Although Paranthropus shows similarly large premolars relative to molars ( Fig. 3 View Fig ), the maxillary postcanine teeth of H. luzonensis differ markedly from those of Paranthropus in absolute size and shape (Extended Data Fig. 2 View Fig ). Multivariate statistical analysis of P 3 –M 2 dental diameters results in H. luzonensis clustering with Asian H. erectus. Both species are characterized by almost similarly sized first and second molars, large premolars relative to molars and postcanine teeth that are mesio-distally compressed (Extended Data Fig. 2g, h View Fig ). The mesio-distally compressed shape of the M 1 of H. luzonensis is also visible in the results of the elliptic Fourier analysis of the crown contour, in which H. luzonensis plots at the margins of the range of variation for H. sapiens (which is characterized by more squared or rhomboidal M 1 s) (Extended Data Fig. 3 View Fig ). H. luzonensis differs from H. floresiensis, which displays an M 1 crown contour shape that is even more mesio-distally compressed and has a more developed protocone (Extended Data Fig. 3c, f View Fig ). Threedimensional geometric morphometric analyses of the P 3 and P 4 EDJs discriminate CCH6 and CCH8 from fossil and extant H. sapiens, H. neanderthalensis and Asian H. erectus, with H. luzonensis occupying its own area of the morphospace or sharing it with H. floresiensis when the latter is included ( Fig. 4 View Fig and Extended Data Fig. 4d, e View Fig ). The 3D geometric morphometric analyses of M 1 and M 2 EDJs are less clear, with H. luzonensis clustering with extant H. sapiens and Asian H. erectus (Extended Data Fig. 4f View Fig ), with extant H. sapiens only (Extended Data Fig. 4g View Fig ), with Asian H. erectus only (Extended Data Fig. 4h View Fig ), or by itself (Extended Data Fig. 4i View Fig ). However, the overall pattern of morphology seen in the dental remains of H. luzonensis is easily distinguishable from all previously described hominin species.
CCH2 is an intermediate manual phalanx with a total length of 32.5 mm (interarticular length, 31 mm; Fig. 2b View Fig , Supplementary Information and Supplementary Table 2). The bone is complete and well-preserved, but it was recovered broken just distal of the midshaft in two refitting parts (Extended Data Fig. 5a View Fig ). On the basis of the asymmetry of the proximal and distal ends, it belongs to rays 2, 3 or 4 of the left hand. CCH2 displays primitive features such as an elongated and dorso-palmarly compressed shaft and well-developed flexor sheath attachments (Extended Data Fig. 5b View Fig ). All these features are observed in Australopithecus 18 – 20 and to a lesser extent in the Homo habilis holotype OH7 21 (but see a previous publication 22), but not in other Homo species that show more derived morphologies 23 – 26 (Extended Data Fig. 5c View Fig ), although a primitive morphology is reported for H. naledi 27. Metric (Extended Data Fig. 5 View Fig d–f) and 3D geometric morphometric analyses (Extended Data Fig. 6 View Extended ) show that the intermediate manual phalanx of H. luzonensis is long, narrow medio-laterally (base, shaft and head) and markedly curved. These shape characteristics are shared with Australopithecus and H. floresiensis, but not with H. naledi (which shares shape affinities with Paranthropus /early Homo specimens from Swartkrans), and are seen occasionally in H. sapiens (Extended Data Fig. 6a, d, e View Extended ). H. floresiensis differs from H. luzonensis and most other hominins by the shape of the phalangeal head, which—in lateral or medial view—has a very small diameter and is only slightly palmarly deviated relative to the proximo-distal axis of the shaft (Extended Data Fig. 6b, c View Extended ). An idiosyncratic feature that differentiates H. luzonensis from all other species of Homo is the strong development and proximal projection of the dorsal beak and this may have limited extension at the interphalangeal joint.
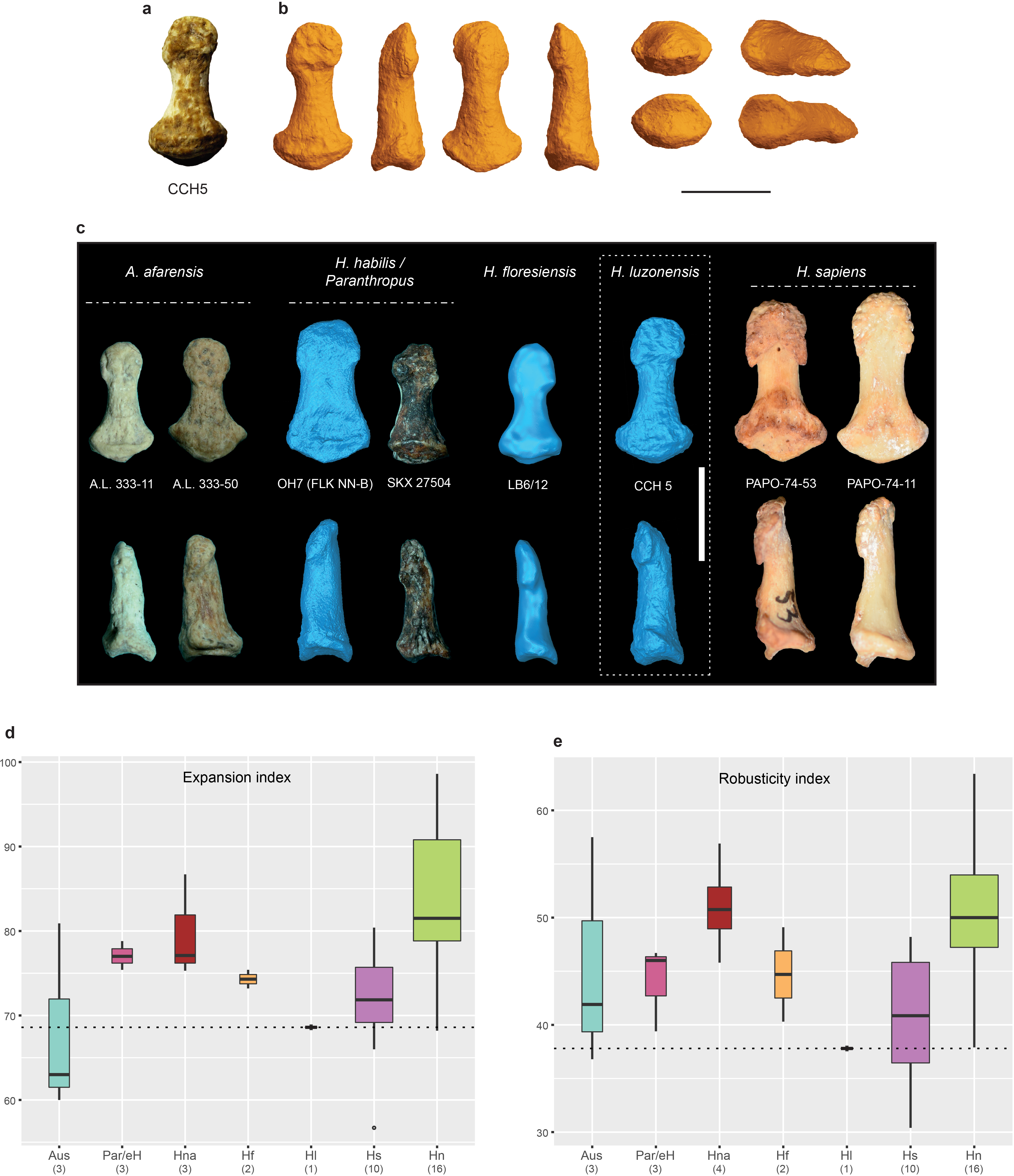
CCH5 is a complete and well-preserved distal manual phalanx 15.9 mm in total length (interarticular length, 15.6 mm; Fig. 2c View Fig , Extended Data Fig. 7 View Extended , Supplementary Information and Supplementary Table 2). Its laterality and ray cannot be determined with certainty, but its proximal articular surface is too small to articulate with CCH2. The proportions of CCH5 (apical tuft expansion and robusticity indices) are within the ranges of variation for H. sapiens and Australopithecus and outside those of H. neanderthalensis and H. floresiensis 23, 28 (Extended Data Fig. 7d, e View Extended ).
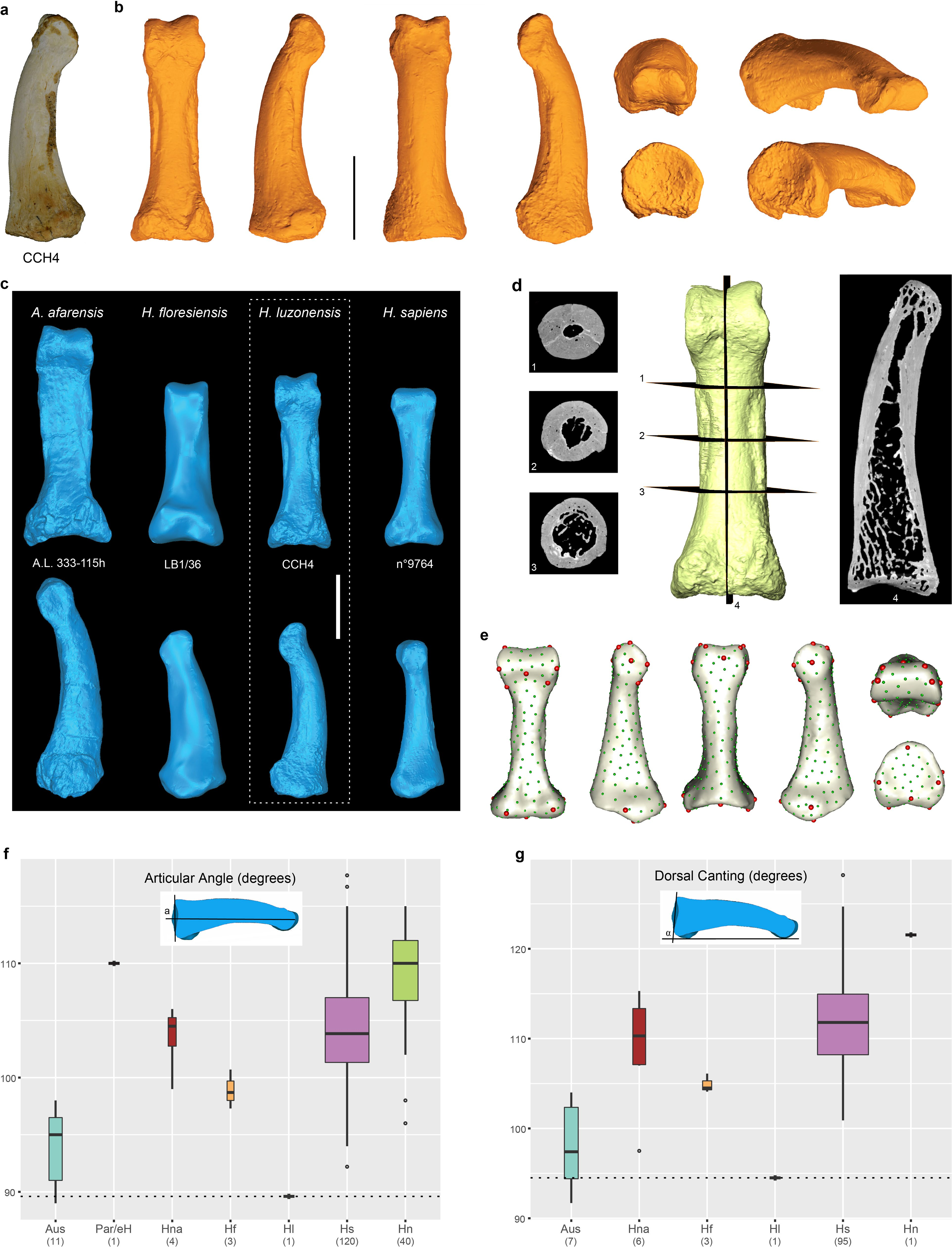
CCH4 is a complete and well-preserved right proximal pedal phalanx of rays 2, 3 or 4 ( Fig. 2d View Fig , Extended Data Fig. 8 View Extended , Supplementary Information and Supplementary Table 2) that is 27.4 mm long (interarticular length, 26.5 mm). It shows primitive— Australopithecus -likefeatures, including pronounced longitudinal curvature of the shaft in lateral view, parallel medial and lateral diaphyseal margins in dorsal view and well-developed flexor sheath attachments in the distal two-thirds of the shaft, bordered by marked plantar crests (Extended Data Fig. 8b, c View Extended ). The base is small, with a circular proximal articular surface that displays dorsal canting and articular angles of 94.5° and 89.6°, respectively. These angular values are outside of the ranges of variation for H. sapiens, H. neanderthalensis, H. naledi and H. floresiensis, but within those of Australopithecus afarensis and Australopithecus africanus 20, 29 – 31 (Extended Data Fig. 8f, g View Extended ). The head is relatively small, bicondylar and proximo-dorsally to disto-plantarly compressed, with a plantar breadth that is larger than its dorsal one. All of these features are typically found in combination in Australopithecus 19, 20, 29, 31. The morphology of the proximal pedal phalanges of H. floresiensis has also been described as Australopithecus -like in some respects (for example, absence of an hourglass shape) 32, 33; however, the combination of features found in CCH4 is essentially indistinguishable from the features of A. afarensis and A. africanus as shown by a 3D geometric morphometric shape analysis ( Fig. 5 View Fig ). Although proximal pedal phalanges are missing from the Asian H. erectus fossil record, those known for African and European species (for example, H. naledi, Homo antecessor and H. neanderthalensis) suggest that the morphology of H. luzonensis is unique among the genus Homo 26, 31, 34.
CCH3 is a complete and well-preserved intermediate pedal phalanx with a total length of 16.3 mm (interarticular length, 14.9 mm; Fig. 2e View Fig , Extended Data Fig. 9 View Extended , Supplementary Information and Supplementary Table 2). Because the shape, size and morphology of this element is highly variable in H. sapiens and other hominins, little can be said at this time about its comparative morphology; however, it visually more closely resembles the intermediate pedal phalanges of H. sapiens and H. floresiensis than it does those of A. afarensis (Extended Data Fig. 9c View Extended ).
The previously published third metatarsal (CCH1) 1, 2 has a proximal base that is particularly small compared to the total length of the bone. It is further characterized by a pronounced triangular shape with a short dorso-plantar height and a medio-laterally expanded dorsal aspect, and a marked dorso-plantar convexity of the proximal articular facet for the lateral cuneiform. The latter feature, which is very uncommon in hominins, has been reported (since the description of CCH1) in a fourth metatarsal of Australopithecus sediba 35.
The referred specimen, CCH7, is a partial left femur from which both the proximal and distal ends are missing ( Fig. 2h View Fig , Extended Data Fig. 10 View Extended and Supplementary Information). Transverse slices of micro-computed tomography (micro-CT) scans show a high density of Haversian canals in the cortical bone that is characteristic of a growing juvenile individual (Extended Data Fig. 10c View Extended ).
Implications of H. luzonensis for hominin evolution
The Late Pleistocene hominin fossils from Callao Cave show a combination of dental and postcranial (hand and foot) features that is distinct from currently known species of the genus Homo (Supplementary Table 3). The premolars of H. luzonensis combine size and shape features seen in other Late Pleistocene species of the genus Homo (for example, H. neanderthalensis, H. floresiensis and H. sapiens) with primitive morphological features of the crown, EDJ and roots, which are typically found in earlier hominins, including Australopithecus and Paranthropus (Supplementary Table 3). The molars are extremely small and their external morphology resembles those of H. sapiens and—to a lesser extent—those of H. floresiensis, whereas some of their EDJ shapes show affinities with those of Asian H. erectus. Notable similarities are observed between H. luzonensis and Australopithecus in the anatomy of their manual and pedal elements. The morphologies of the hands and feet of Australopithecus , which are generally described as an intermediate between the morphologies of great apes and modern humans, are typically interpreted either as indicating adaptations to various degrees of bipedalism and climbing and/or suspension 36 or as reflecting the retention of plesiomorphic features in obligatory bipeds 35, 37. However, the partial and fragmentary nature of the H. luzonensis postcranial elements presently limits further interpretation of its locomotor and manipulative abilities.
The origin of H. luzonensis , as well as its phylogenetic relationships with other hominins present in eastern Asia at around the same timeincluding H. sapiens 38, H. floresiensis 39 and Denisovans 10, and hominins recently discovered in China 40 —remains to be determined. As is the case for most hominins recovered from tropical Asia 41, attempted DNA extraction from H. luzonensis fossils has been unsuccessful. Direct dates on H. luzonensis tooth and bone samples indicate it was present on Luzon before 50 kyr ago 1, 3 and the recent discovery 42 of stone tools and a butchered rhinoceros in the nearby Cagayan Valley indicates that hominins, perhaps H. luzonensis , have been present on Luzon since more than 700 kyr ago. Clearly, both H. luzonensis and H. floresiensis were present east of the Wallace Line on Luzon and Flores, respectively, at the same time 39 and perhaps even over a similar temporal interval 17, 42. The skeletons of both species present anatomical traits that are either rare or absent elsewhere in the genus Homo but have similarities with those of Australopithecus 6, 7, 23, 32, 33, 43 – 45. As is the case with the island of Flores, a substantial sea crossing has always been required to reach Luzon from any mainland, even during the lowest sea level periods of the Quaternary period 46 ( Fig. 1 View Fig ). The insular nature of Luzon and the Philippine archipelago east of Palawan more generally is reflected in the high rates of endemism recognized in vertebrate faunas 47 – 49. Given the highly endemic nature of the Luzon vertebrate faunas, evolutionary convergences and/or reversals under the effects of insular evolution 50 may explain the distinct anatomy of H. luzonensis ; however, further discoveries and more definitive evidence are needed. The discovery of H. luzonensis underscores the complexity of the evolution, dispersal and diversity of the genus Homo outside of Africa, and particularly in the islands of Southeast Asia, during the Pleistocene.








No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.