
Polyonyx sasekumari, Osawa & Naruse & Ng, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4429.2.6 |
|
publication LSID |
lsid:zoobank.org:pub:E42066C8-070D-4742-80FE-726B9B92883A |
|
DOI |
https://doi.org/10.5281/zenodo.5950046 |
|
persistent identifier |
https://treatment.plazi.org/id/B61EB057-FFF0-FFD4-FF06-AD06A58EF391 |
|
treatment provided by |
Plazi |
|
scientific name |
Polyonyx sasekumari |
| status |
sp. nov. |
Polyonyx sasekumari View in CoL n. sp.
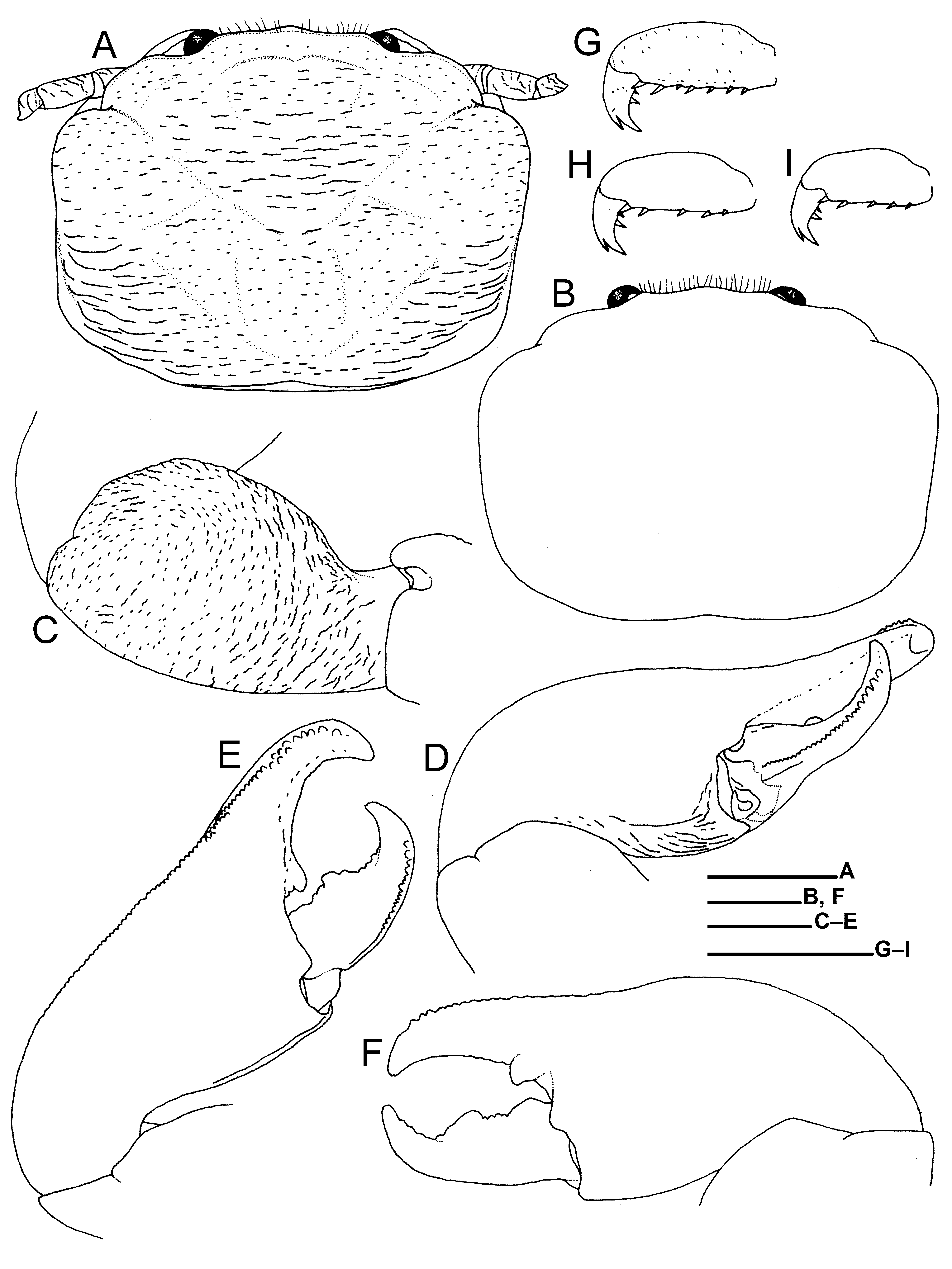
( Figs. 1E, F View FIGURE 1 , 6–8 View FIGURE 6 View FIGURE 7 View FIGURE 8 )
Type material. Holotype: ZRC 2017.200 View Materials , male (cl 3.7 mm), Jeram , Kuala Selangor, Selangor, Malaysia, Jeram Polychaete Reef, sta. C, 3°13'27"N, 101°18'13"E, lower intertidal, mud, July–October 2016, coll. J. J. Eeo. GoogleMaps
Paratypes: ZRC 2016.527 View Materials , 4 males (cl 3.2–3.8 mm), 4 ovigerous females (cl 3.9–4.8 mm), collected with holotype GoogleMaps ; RUMF-ZC-5236, 1 ovigerous female (cl 4.0 mm), collected with holotype GoogleMaps .
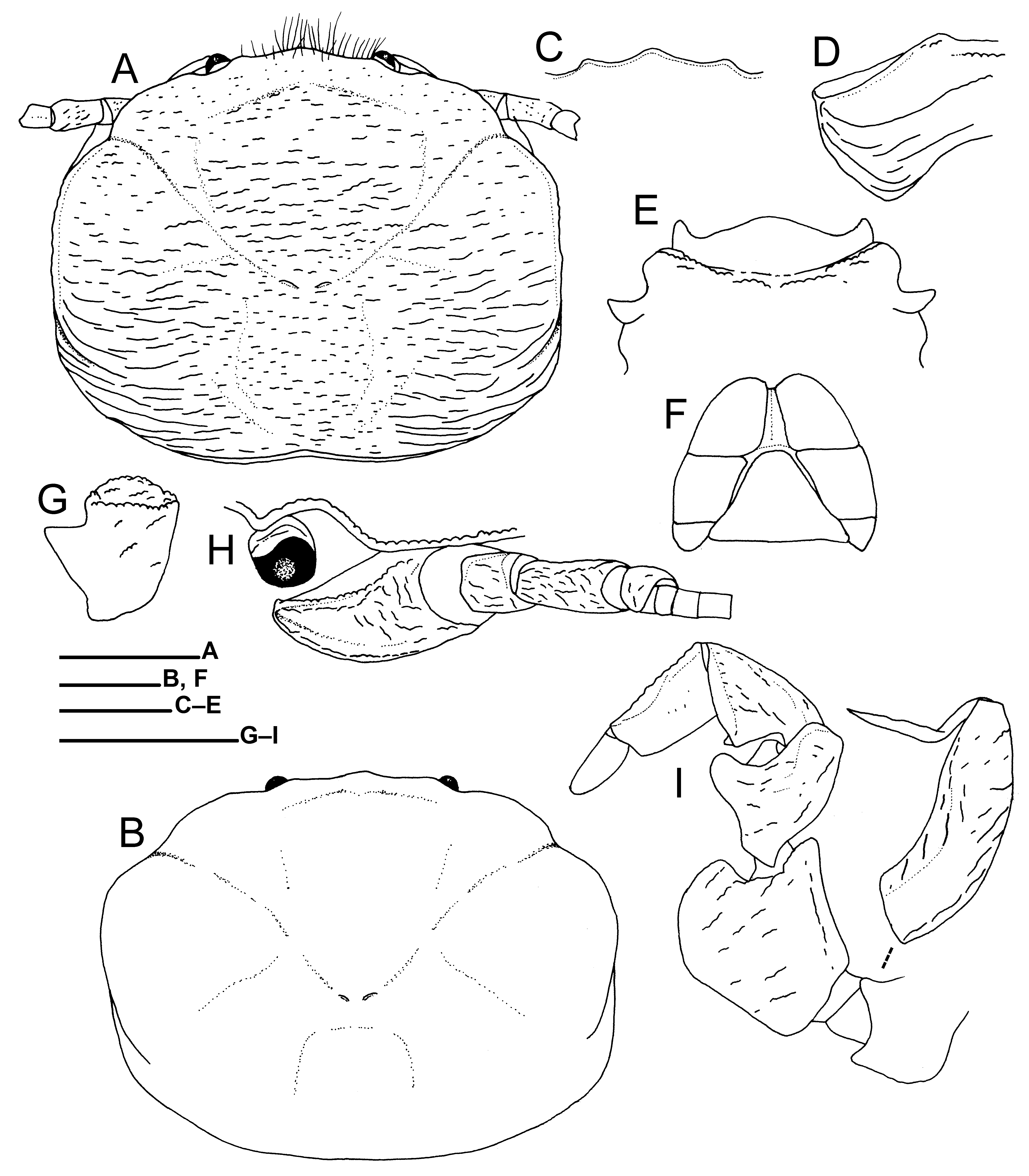
Description. Carapace ( Fig. 6A, B View FIGURE 6 ) rectangular in general outline, 1.2–1.3 (male) and 1.3–1.4 (female) times as broad as long, broadest on median branchial margins. Dorsal surface somewhat convex from side to side, entirely covered with delicate, short and somewhat longer, transverse striae. Protogastric ridges weakly demarcated, obtuse. Cervical grooves moderately demarcated. Hepatic margins moderately convex. Branchial margins moderately convex, subparallel on median parts; anterior half margins thin; posterior margins and adjacent regions with long, transverse ridges. Rostrum ( Fig. 6C View FIGURE 6 ) moderately broad, slightly produced beyond eyes in dorsal view, ventrally bent toward anterior tip; trilobate in frontal view, median lobe bluntly triangular, overreaching rounded lateral lobes, without median longitudinal groove; anterior margin and dorso-anterior median surface with short plumose setae. Orbits shallow; supra-orbital margins nearly straight; lateral orbital angles obtuse.
Pterygostomial flaps entire, with short and long, longitudinal ridges; dorso-anterior margin terminating in narrowly rounded tip ( Fig. 6D View FIGURE 6 ).
Third thoracic sternite ( Fig. 6E View FIGURE 6 ) 3.3–4.0 (male) and 4.3–5.3 (female) times as broad as long measured on midline, with surface depressed medially; anterior margin trilobite, with sparse long setae; median lobe broad, convex on anterior margin; lateral lobes narrow, nearly reaching anterior margin of median lobe, each with rounded apex. Fourth thoracic sternite ( Fig. 6E View FIGURE 6 ) with anterior margin moderately concave; surface depressed medially, with transverse series of short ridges anteriorly.
Telson ( Fig. 6F View FIGURE 6 ) composed of 7 plates; proximolateral plates much smaller than others; distal plates as long as or slightly longer than broad. Ocular peduncles ( Fig. 6A, B, H View FIGURE 6 ) small, short; dorsal extension onto cornea low, rounded.
Basal article of antennular peduncle ( Fig. 6G View FIGURE 6 ) slightly longer than broad; anterior margin convex; ventral surface with transverse ridge anteriorly and short rows of granules medially; ventro-anterior face slightly concave.
Antennal peduncle ( Fig. 6H View FIGURE 6 ) slender, with irregular, short striae on surfaces. First article largest, approximately equal in length to combined second to fourth articles, produced forward in lateral view, broadly in contact with lower orbital margin, with blunt ridge each along dorsal and ventral margins; surface shallowly concave; anterior margin narrowly rounded. Second and third articles roundly rectangular, third article elongate. Fourth article short, rounded.
Third maxilliped ( Fig. 6I View FIGURE 6 ) with coxa bearing blunt projection on ventrodistal margin; distomedian projection with blunt transverse ridge, not articulated. Basis articulating with ischium, rounded subtriangular. Ischium broad, ovate; lateral surface with short ridges and longitudinal ridge along dorsal margin; dorsodistal projection blunt. Merus with laminate, rounded lobe on ventrolateral margin; lateral surface with short transverse ridges and short longitudinal ridges along dorsal margin. Carpus with small subtriangular projection on median part of ventral margin and with longitudinal ridge on dorsal surface. Propodus moderately slender, nearly smooth on lateral surace. Dactylus short, subtriangular. Merus to dactylus with long plumose setae on ventral margins. Exopod with proximal article small, rounded; distal article laminate, robust, reaching midlength of merus, with row of transverse ridges dorsally and ventrally; distal flagellum present.
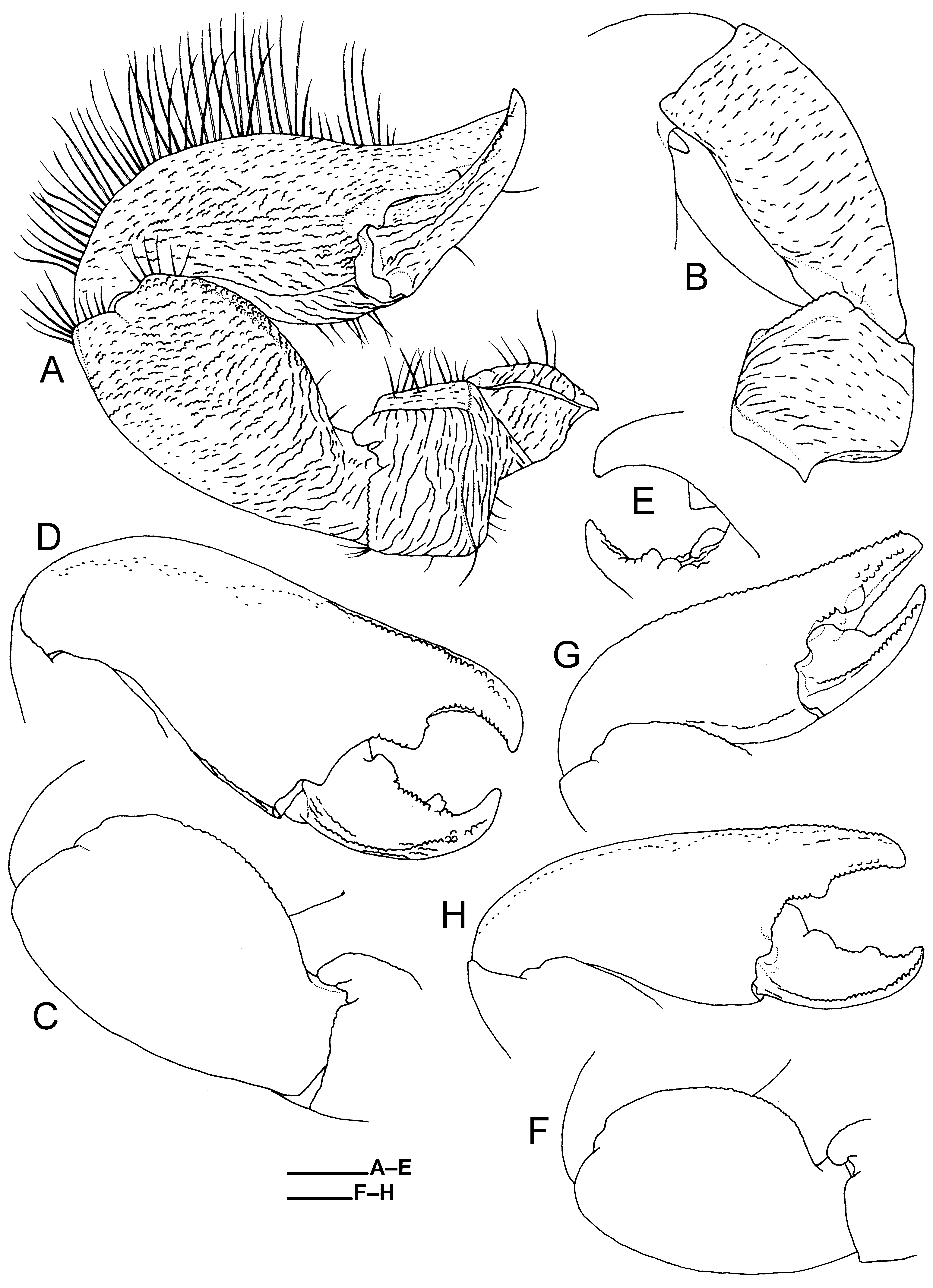
Chelipeds (first pereopods, Figs. 7A–H View FIGURE 7 , 8A–C View FIGURE 8 ) unequal in size; dorsal surfaces with long transverse striae on meri and with numerous, delicate, short and very short striae on carpi and chela (striae on dorsoroximal parts of carpi longer); entire margins of meri and carpi and posterior margins of chelae with sparse, short to moderately long plumose setae; anterior margins of palms with dense, long plumose setae; ventrodistal margins of carpi, ventroposterior parts of palms, and cutting regions of fingers with short to moderately long, plumose setae. Larger cheliped ( Fig. 7A–H View FIGURE 7 ) with ischium bearing no spines on ventro-anterior margin. Merus with rounded transverse crest submedially on dorsal surface; dorso-anterior margin with narrow, rounded lobe distally; ventral surface with short transverse striae; ventro-anterior distal corner rounded. Carpus 1.6 times as long as broad, broadest on median part; dorsal surface slightly convex, posterior margin rounded; dorso-anterior margin with elevated lobe, anterior margin of lobe convex; proximal concavity on dorso-anterior margin short; dorsodistal margin with low, rounded lobe on posterior part; ventral surface with short transverse striae; ventro-anterior margin concave on subdistal part. Chela 1.4–1.5 times as long as carpus, elongate ovate in general outline, 2.3–2.5 times as long as broad, lying on anterior side; anterior margin of distal part of palm and fixed finger subacute, with row of small rounded tubercles. Palm somewhat inflated; dorsal surface convex; dorsoposterior margin with longitudinal blunt ridge; ventral surface convex, with short and very short striae anteriorly. Fixed finger with weakly curved distal claw; dorsal surface with scattered, small pits and blunt ridge along distal part of midline; cutting edge with large, subtriangular tooth proximally followed by small rounded teeth decreasing in size distally. Dactylus 0.4 length of chela, opening at oblique angle, anteriorly curved in dorsal view, with curved distal claw; dorsal surface with longitudinal striae posteriorly; posterior margin distally with short row of small tubercles; cutting edge with proximal large, rounded tooth followed by row of small rounded teeth and with 2 larger blunt teeth (proximal tooth low and smaller than distal tooth) ventrally on median part.
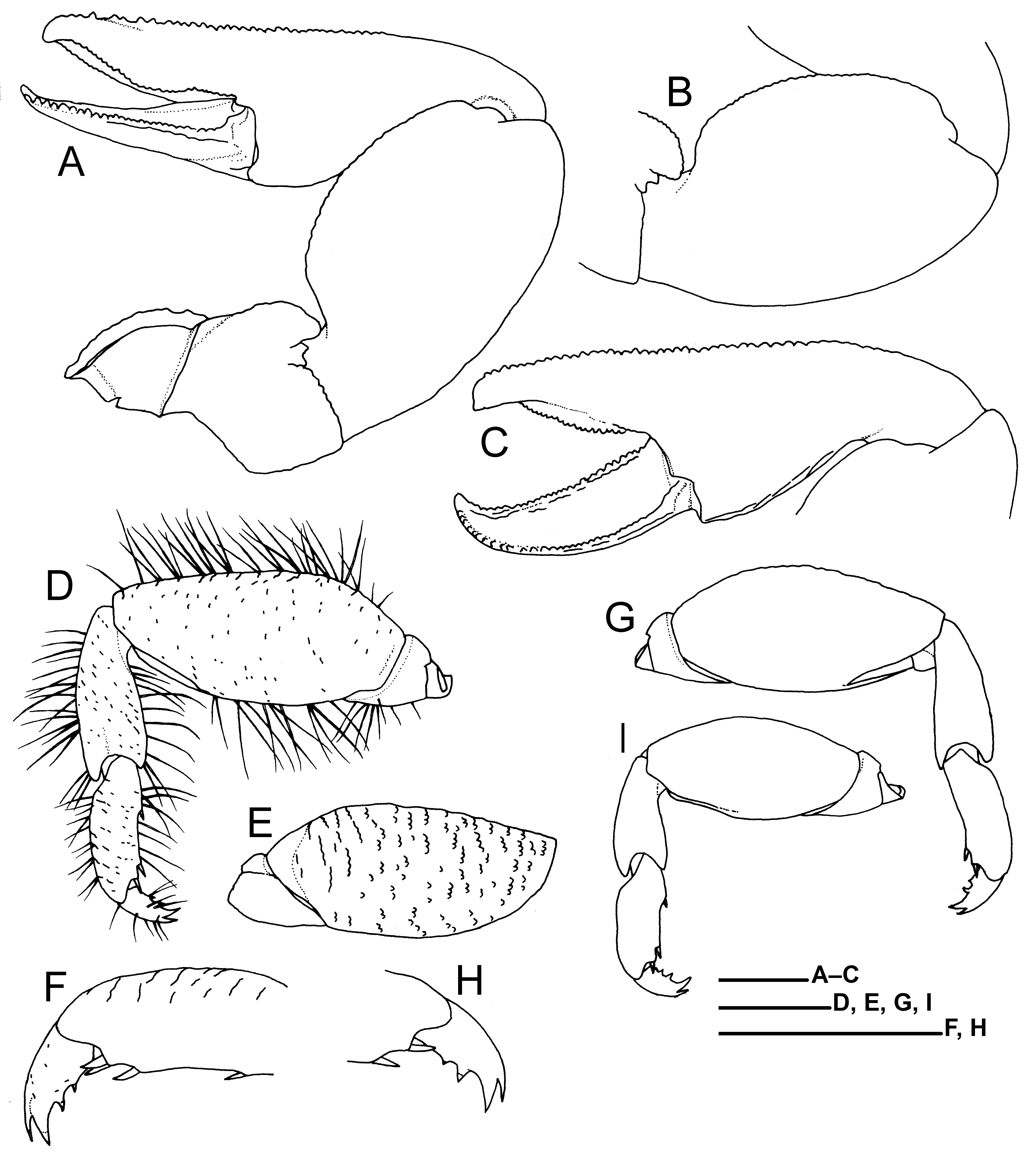
Smaller cheliped ( Fig. 8A–C View FIGURE 8 ) similar to larger cheliped in general, except for: carpus with nearly straight, ventro-anterior margin; chela proportionally narrower, 2.6–2.9 times as long as broad, less convex on dorsal surface, with row of small rounded tubercles on distal 0.7 of anterior margin; plumose setae on anterior margin of palm extending onto fixed finger and anterodorsal surface of chela; denser plumose setae on ventroposterior part of palm, and cutting regions of fingers; cutting edge of fixed finger slightly convex, with row of small rounded teeth; cutting edge of dactylus straight or slightly concave, with row of small rounded teeth; dactylus proportionally longer, 0.4–0.5 length of chela, with posterior margin bluntly elevated.
Ambulatory legs (second to fourth pereopods, Fig. 8D–I View FIGURE 8 ) moderately short, decreasing in size posteriorly (second largest), somewhat compressed laterally, with scattered, short to moderately long plumose setae (lateral setules minute) marginally; lateral surfaces nearly smooth except for short ridges on laterodorsal parts of meri and propodi; mesial surfaces with scattered, short granular ridges on meri, carpi, and propodi, but nearly smooth on dactyli. Ischia each with small, weakly calcified part dorsally along distal margin of mesial surface. Meri 2.2–2.4 (second), 2.0–2.3 (third), and 1.9–2.1 (fourth) times as long as high in lateral view, highest slightly proximal to or just on median part; dorsal margins slightly crenulated (second and third) or smooth (fourth), gently convex; lateral surface of fourth pereopod with scattered, short setae dorsally; mesial surfaces with weakly calcified part proximally, that part broadening dorsally; ventrodistal margins of lateral and mesial surfaces rounded. Carpi moderately slender (second and third) or stout (fourth); dorsodistal and ventrodistal corners rounded. Propodi 2.7– 2.8 (second), 2.6–2.8 (third), and 2.4–2.7 (fourth) times as long as high, highest at base of subproximal ventral corneous spine if present; dorsal margins nearly smooth; ventral margins each with no or 1 corneous spine (usually present; e.g., absent on right third and fourth pereopods of holotype) on proximal part and 1 corneous spine and distal part; distoventral margin with 2 corneous spines subequal in size. Dactyli 0.4–0.5 times as long as propodi, each terminating in curved, sharply pointed, bifurcate claws, ventral claw much larger than dorsal claw; ventral margins each with 2 small corneous spines each on slightly or moderately elevated base, distal spine larger than proximal spine.
Fifth pereopod slender, chelate; propodus with numerous short simple setae and 2 subdistal hooked setae.
Male with pair of gonopods on second abdominal segment; endopod elongate ovate, rounded on distal margin; exopod small, rounded.
Female with no pleopods on third abdominal segment, but with well developed pleopods on fourth and fifth abdominal segments.
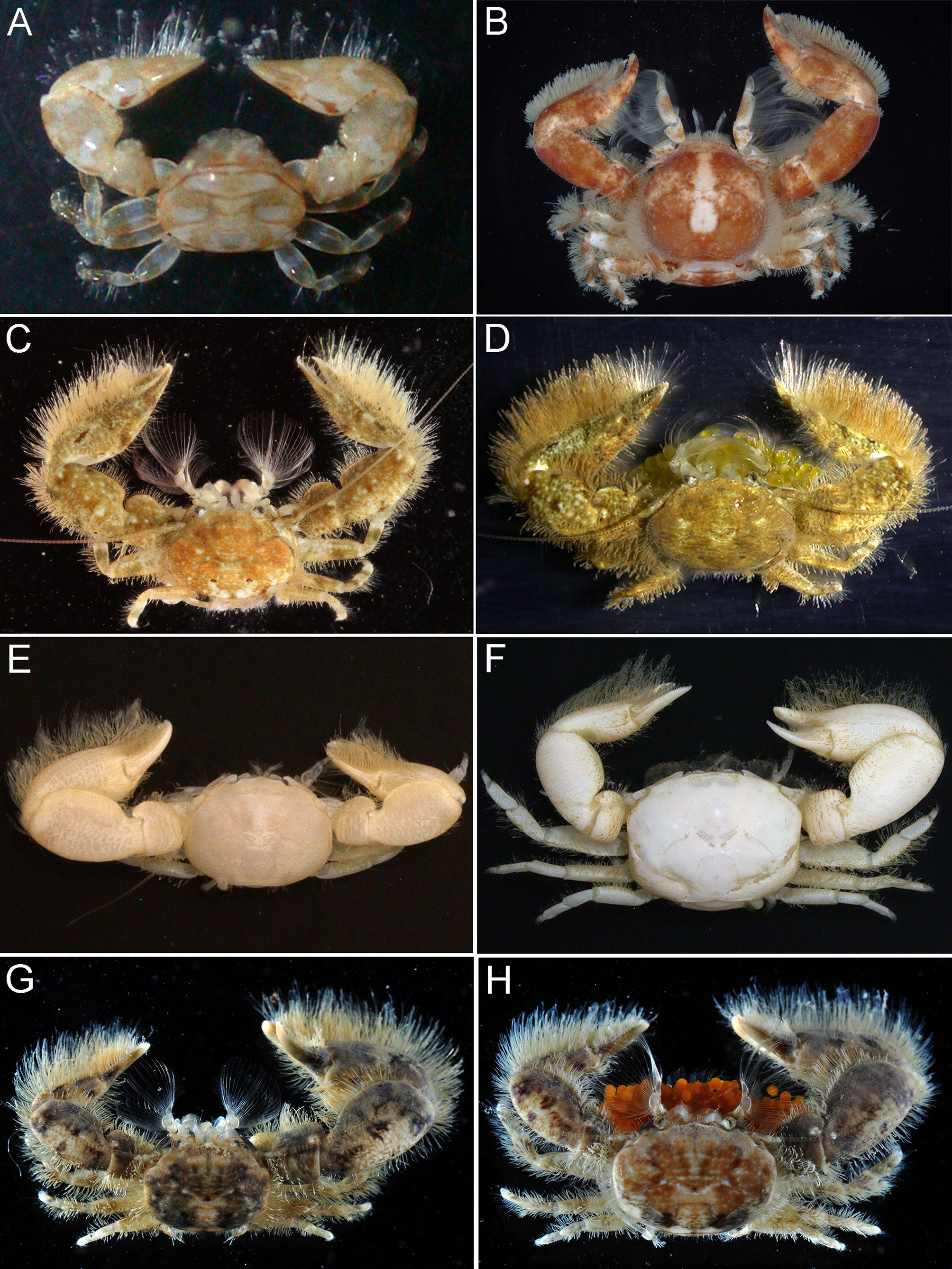
Coloration in life. Not known. Preserved specimens entirely white or pale brown ( Fig. 1E, F View FIGURE 1 ).
Distribution. Presently known only from Jeram, Selangor, western coast of Peninsular Malaysia.
Habitat. The type specimens were collected from the lower intertidal region of muddy shore with intermittent reefs of sabellariid polychaetes. Polgar et al. (2015) reported the environment and community composition of polychaetes at the Jeram beach from December 2010 to April 2013 and the remarkable changes of the beach during the research period. According to the report by Polgar et al. (2015: 408), in April 2013, the whole tidal flat was covered with a superficial layer of mud and heavily colonized by Loimia verrucosa Caullery, 1944 and there were pairs of openings (spaced at ~ 60 cm), possibly corresponding to large U-shaped burrows dug by polychaete worms. Although there are no definite associated host records for the type specimens of Polyonyx sasekumari n. sp., the large U-shaped burrows may be referred to dwelling-tubes unique to species of Chaetopterus (cf. Nishi 2001; Nishi et al. 2009). The candidate for the host of P. sasekumari n. sp. is also supposedly a species of Chaetopteridae as known for its related congeners discussed below in Remarks.
Etymology. The new species is named after Dr. A. Sasekumar of the University of Malaya, who so kindly passed us the specimens for study.
Remarks. Polyonyx sasekumari n. sp. appears closest to P. thai known from Chonburi Province in Thailand and the Johor Straits in Singapore (present study) in having the following characters: the carapace being clearly longer than broad and with only delicate, short transverse striae on the dorso-anterior half and no scattered setae on the entire dorsal surface, the chelipeds without dense coat of setae on the entire dorsal surface, the carpus of each cheliped with a strongly bulging dorso-anterior margin except for a distinct, proximal short concavity, the palm of each cheliped without scattered tubercles on the dorsal surface, and the ambulatory legs without distinct spines on the ventral margin of each merus and usually with at least two spines in addition to a pair of distal spines on the ventral margin of each propodus. However, the new species is distinguished from P. thai by the morphology of the propodi of the ambulatory legs. The segments of P. sasekumari n. sp. are proportionally more slender: 2.7–2.8, 2.6–2.9, and 2.4–2.7 times as long as high on the respective second to fourth pereopods, instead of 2.2–2.5, 2.5– 2.6, and 2.3 times in P. thai; and they usually have only one subproximal and one subdistal spines on each ventral margin, in addition to a pair of distoventral spines, instead of four to nine ventral spines in P. thai (for the characters of P. thai, cf. Werding 2001: fig. 4G–K; present study, usually five or six spines in the Singapore material examined, Fig. 9G–I View FIGURE 9 ). Additionally, the eyes are smaller compared with the size of the carapace and the proximal concavity on the dorso-anterior margin of the carpus of each cheliped is narrower in P. sasekumari n. sp. than in P. thai. For habitats, P. sasekumari n. sp. was collected from the intertidal muddy shore with intermittent reefs of sabellariid polychaetes and had no definite records of the association host, whereas P. thai was noted by Werding (2001) to be obtained from the exposed sand beach and inside of tubes of chaetopterids together with a series of P. sinensis . As shown below, the specimens examined of P. thai from the Johor Straits, Singapore, were collected from the depths of 6.9–24.4 m, but without any host data. In Japan, P. sinensis has been known to be associated with Mesochaetopterus japonicus Fujiwara, 1934 of the Chaetopteridae ( Miyake 1965; Watanabe 2013). The general shape and ornamentation of the carapace and pereopods also links P. sasekumari n. sp. to P. loimicola, P. sinensis , P. tulearis, and P. vermicola , but the detailed outline of the carpus of the larger cheliped distinguishes the new species from the other four species. The segment is proportionally shorter in the new species than in P. loimicola, P. sinensis , and P. vermicola . The dorso-anterior margin of the carpus of the larger cheliped
more strongly bulges in P. sasekumari n. sp. than in P. loimicola and P. tulearis; and it is evenly convex on the almost entire part in the new species instead of strongly convex on the distal half in P. sinensis and P vermicola (for characters of the four known species, cf. Sankolli 1965; Ng & Sasekumar 1993; Werding 2001; Osawa & Chan 2010; personal observation).
Polyonyx vermicola View in CoL was described by Ng & Sasekumar (1993) on the basis of material from Morib (Kuala Langat, Selangor), situated south of the type locality of the present new species. However, the habitat of the respective type localities of P. sasekumari View in CoL n. sp. and P. vermicola View in CoL is different. Unlike mentioned above for the new species, Ng & Sasekumar (1993) noted that P. vermicola View in CoL was collected from sheltered sand beaches and inside of tubes of the chaetopterid Mesochaetopterus selangora ( Rullier, 1976) View in CoL (for the taxonomy and name of the polychaete species, cf. Nishi 1999; Read 2015).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Polyonyx sasekumari
| Osawa, Masayuki, Naruse, Tohru & Ng, Peter K. L. 2018 |
P. sasekumari
| Osawa & Naruse & Ng 2018 |
Polyonyx vermicola
| Ng & Sasekumar 1993 |
P. vermicola
| Ng & Sasekumar 1993 |
P. vermicola
| Ng & Sasekumar 1993 |
Mesochaetopterus selangora (
| Rullier 1976 |