
Scelidosaurus (NHMUK R, 1111)
|
publication ID |
https://doi.org/10.1093/zoolinnean/zlaa061 |
|
DOI |
https://doi.org/10.5281/zenodo.10541467 |
|
persistent identifier |
https://treatment.plazi.org/id/B66BDD2A-082B-FF9E-E367-74C5FEB6E16B |
|
treatment provided by |
Felipe |
|
scientific name |
Scelidosaurus |
| status |
|
SCELIDOSAURUS : CRANIAL BIOLOGY
EXTERNAL FEATURES
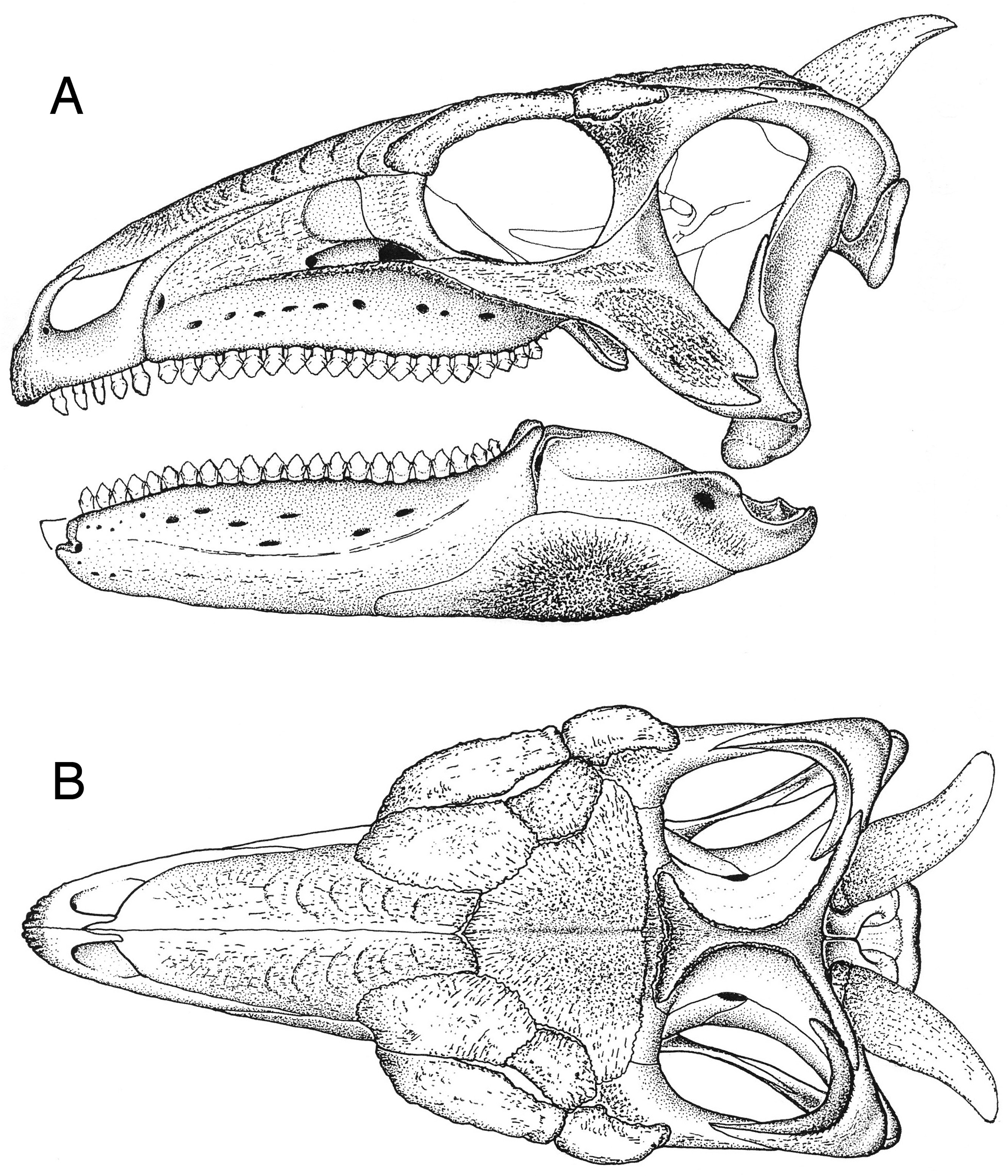
The surfaces of the skull and mandible ( Fig. 4 View Figure 4 ) are coated by widespread exostoses and a small number of discrete osteoderms. This suggests that the scelidosaur skull was encased by an array of keratinous epidermal scutes or plates.
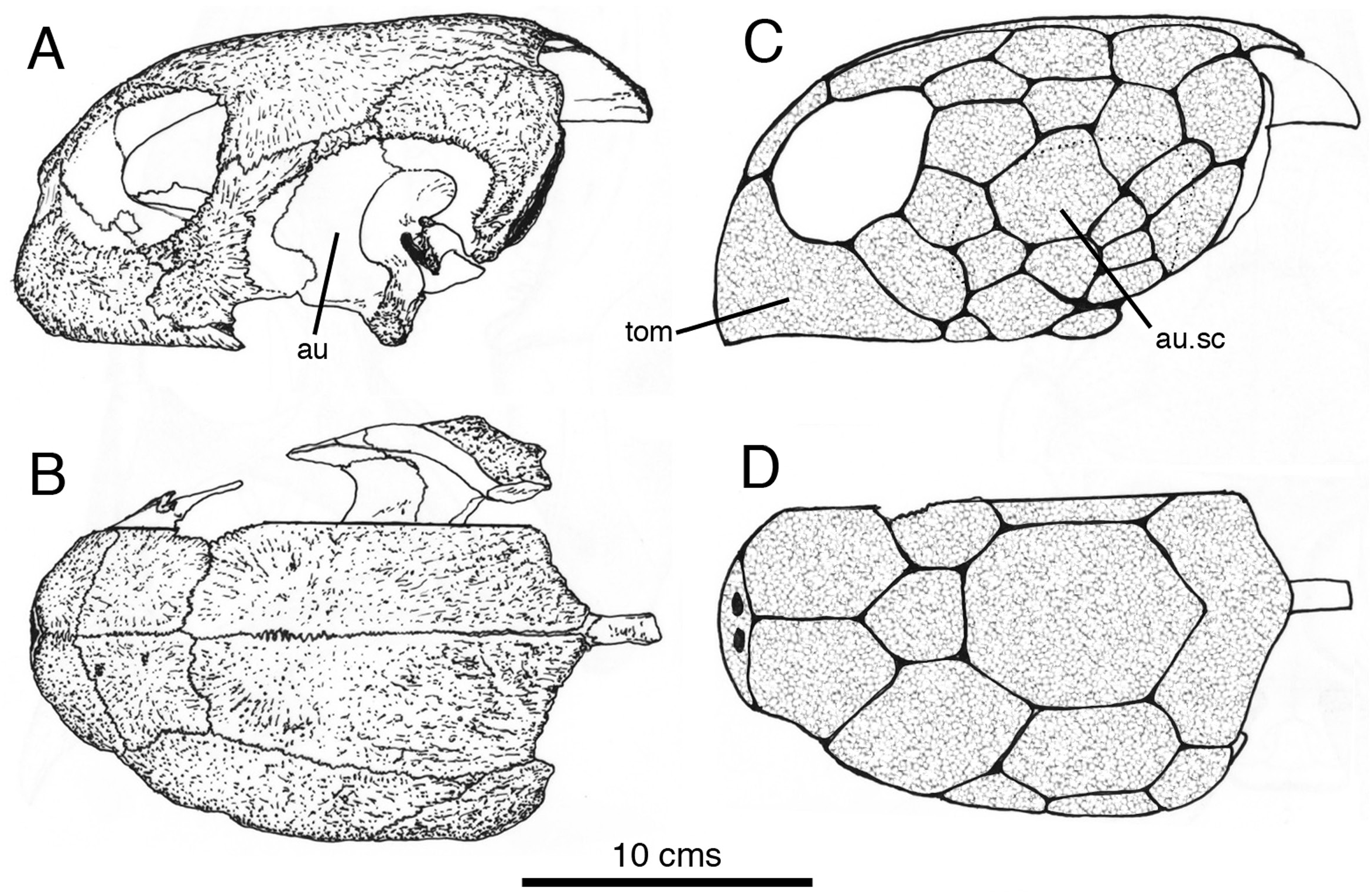
For comparison, an osteological preparation of the skull of a similarly sized subadult green turtle [ Chelonia mydas ( Linnaeus, 1758) – Norman, personal collection – Fig. 5A, B View Figure 5 ] – a taxon belonging to the clade Testudinata with putative (sister-taxon) archosaur affinities ( Crawford et al., 2012) – reveals similarly heavily textured skull roofing bones that, in life, are known to anchor a tessellate covering of keratinous scutes ( Fig. 5C, D View Figure 5 ). Unfortunately, this particular specimen does not show the pattern of smooth, shallow grooves that mark the edges of some of the principal cranial scutes (Penkalski, pers. comm. March 2020).
The premaxillary beak
In Scelidosaurus a rhamphotheca was, without much doubt, an externally smooth casque-like structure referred to as a tomium in Chelonia ( Fig. 5C View Figure 5 , tom) that enveloped the premaxillae ( Fig. 6 View Figure 6 , rsc). The external surface of the premaxilla (not including the narial fossa) is slightly rugose and pitted with small foramina that in combination would have supported, anchored and indicate the presence of a vascular supply that provided nutrients for the growth of an overlying rhamphotheca; closely comparable osteological features are seen underlying the tomium of the turtle. The scelidosaur rhamphotheca formed a short cuttingedge along the edentulous margin of the premaxilla ( Figs 4 View Figure 4 , 6 View Figure 6 ). The slightly rugose and vascularized lateral wall to the dentulous portion of the premaxilla is likely to have been similarly encased by a posterior extension of the rhamphotheca, in conformity with the structure seen in Chelonia . It is probable that the premaxillary dentition was ensheathed (and supported) by this portion of the keratinous beak; the crowns of the teeth are envisioned projecting from behind the rhamphothecal parapet ( Fig. 6A View Figure 6 ), even though there is no modern analogue for such a composite arrangement. The dorsal portion of the rhamphotheca coated the external surface of the premaxillae and would have extended as far dorsally as the base of the dorsomedian premaxillary process, but would have been cut back so that it skirted the ventrolateral portion of the external naris. Its posterodorsal edge would have merged with the rhamphothecal margin near the posterior end of the premaxilla on either side of the snout ( Fig. 6A View Figure 6 ).
The snout
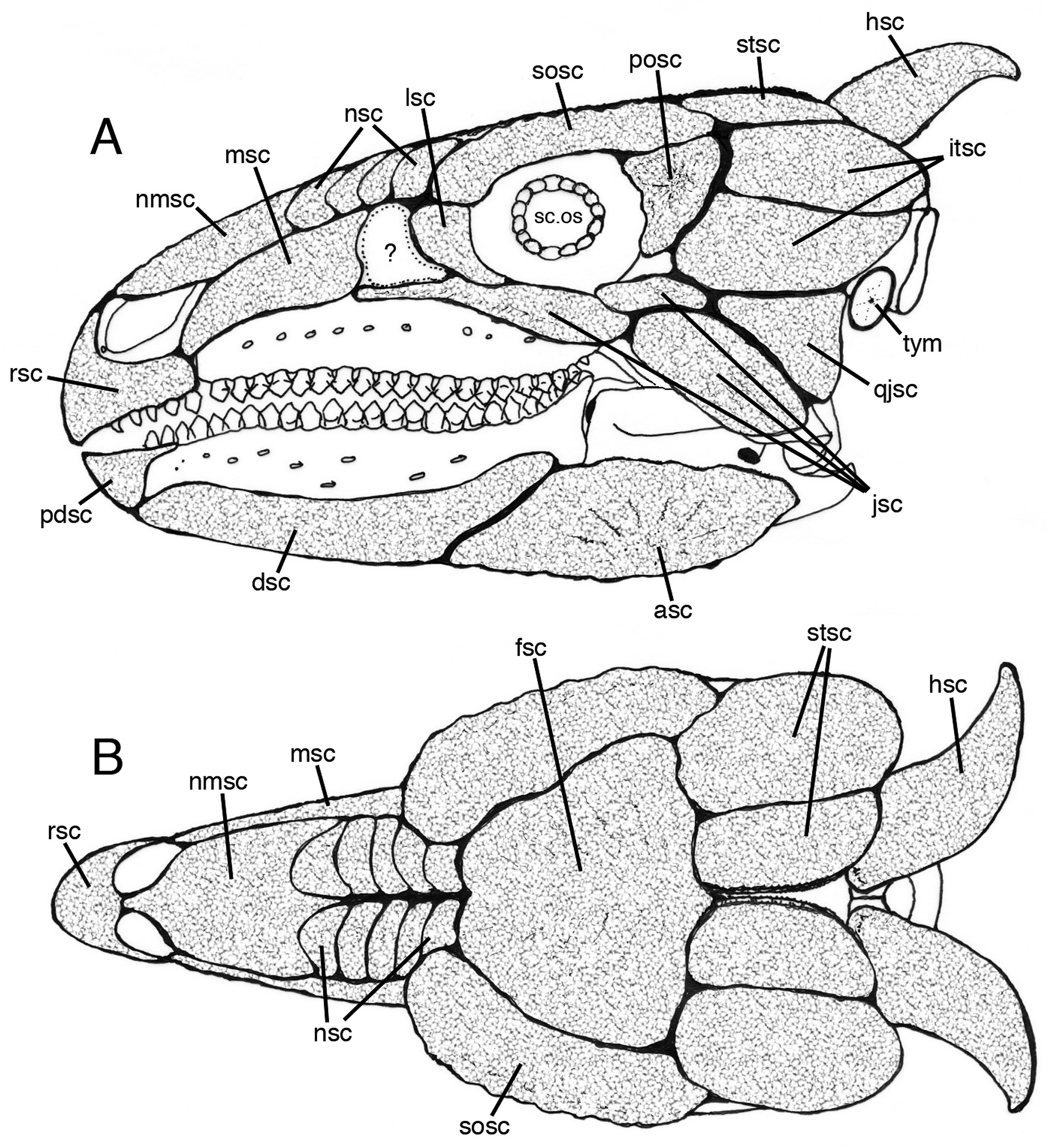
Anteriorly, the dorsal surface of the nasals ( Fig. 4B View Figure 4 ) is characterized by a radiating pattern of strands of bony tissue that may have anchored a midline scute ( Fig. 6A, B View Figure 6 , nmsc); this underlying bony pattern corresponds to that which supports similar midline scutes on the chelonian skull ( Fig. 5B, D View Figure 5 ). Farther posteriorly, the surface of these bones develops a thicker knobbly texture that is overprinted by a series of repeated curved ridges. The ridges are oriented (more or less) transversely across the roof of the snout and extend down the sides of the snout where it is walled by the maxilla and prefrontal. The repetition of the curved ridges is suggestive of attachment sites for successive (possibly overlapping) scutes that encased the snout above the buccal emargination and extended posteriorly as far as the nasofrontal suture ( Fig. 6 View Figure 6 , nsc). The lateral wall of the snout formed by the maxilla and premaxilla is reconstructed here covered by a large maxillary scute ( Fig. 6 View Figure 6 , msc), but there are indications ( Fig. 4A View Figure 4 ) of faint attachment ridges, so it is possible that a series of overlapping scutes were found here as well. Posterior to the reconstructed lateral maxillary scute there is a smooth patch of bone ( Fig. 6 View Figure 6 ,?) behind which the lacrimal bears irregular exostotic growth that would have supported an overlying scute ( Fig. 6 View Figure 6 , lsc).
There is no equivalent patterning of successive curved exostotic ridges on the chelonian skulls that I have examined, so the scute pattern in the scelidosaur skull probably differs from the mosaic-like tessellate pattern of scutes seen in these living, albeit distantly related, diapsid taxa ( Fig. 5 View Figure 5 ).
The skull roof and occiput
The frontals are dominated by a dense pattern of strand-like superficial bone that radiates from a midline groove ( Fig. 4B View Figure 4 ). As with the anterior portion of the nasal, this morphology is suggestive of the presence of a large, shield-like scute ( Fig. 6B View Figure 6 , fsc) that extended across to the adjacent surfaces of the prefrontal, middle supraorbital and postorbital.
Lateral to the frontal plate, the palpebral (= anterior supraorbital) and posterior supraorbital osteoderms form a shallowly arched bar of bone (= brow ridge) that flanks the dorsal orbital margin of the skull roof. The rugose external surfaces of these bones are structurally distinct from the frontal plate, and are likely to have anchored their own substantial keratinous sheaths ( Fig. 6 View Figure 6 , sosc) that served to shield the orbit and its associated soft tissues. It is possible that these keratinous scutes were subdivided into smaller units than illustrated here, and may have been superficially ornate for behaviourally related reasons; enlarged, sculpted and colourful circumorbital scutes are seen in many living squamates.
A rugose, double-ridged sagittal crest dominates the posterior part of the skull roof. The ridges are flanked by large ovoid supratemporal fenestrae and behind these latter is a divergent pair of prominent, curved, horn-like occipital osteoderms. The temporal arches and occipital margin show some evidence of irregular exostotic growth that may well reflect the attachment of overlying scutes ( Fig. 6 View Figure 6 , stsc). It is, of course, possible that this posterior part of the skull table was less extensively scute-covered. The fenestrae themselves would have been spanned by skin that (although scaly) retained a degree of flexibility to allow movement of the underlying temporal musculature. The same consideration should also apply to the adjacent infratemporal fenestrae (itsc). However, there are areas of the skull of Chelonia mydas where exostotic bone is absent and the bone surface is, instead, smooth-surfaced ( Fig. 5A View Figure 5 , au); this area (the margins of which are dotted in Fig. 5C View Figure 5 ) is covered in life by several scutes ( Fig. 5C View Figure 5 , ausc). Therefore, it is possible that in Scelidosaurus tessellated scutes enveloped the posterior skull roof, as well as its lateral flanks. The restoration ( Fig. 6A, B View Figure 6 ) includes an imaginary array of rather large scutes in these areas.
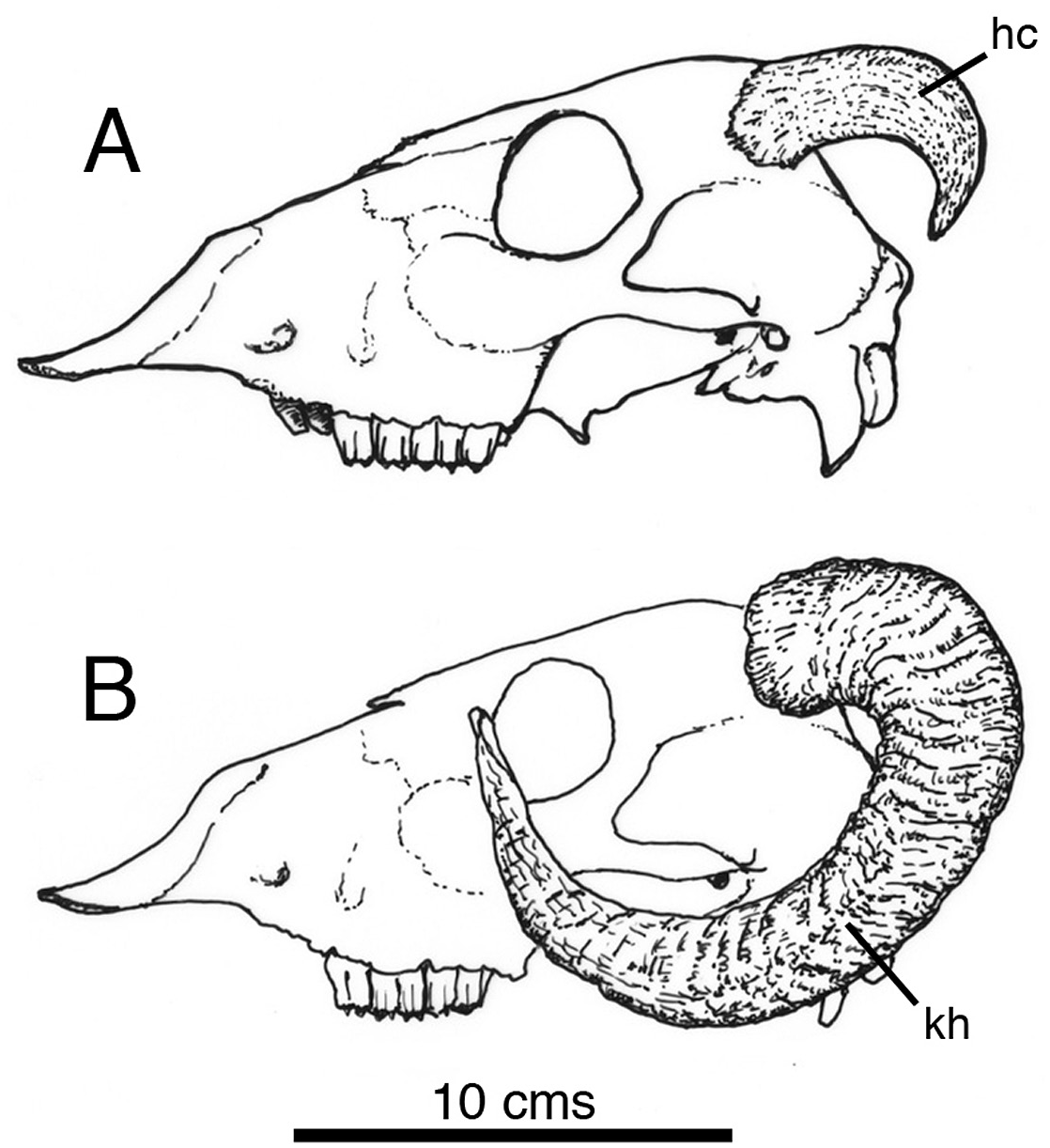
The occipital osteoderm ‘horns’ have comparatively smooth, finely grooved surfaces pock-marked by many tiny foramina; these features are interpreted as a combination of points for the connective tissue that tethered an overlying keratinous horn ( Fig. 6 View Figure 6 , hsc), and the vascular supply for its continued growth. Similar textures are visible on the horn-cores of living bovid mammals (e.g. Ovis aries – Norman, pers. colln – Fig. 7A View Figure 7 , hc). As can be seen in this example, the shape of the horn core may not necessarily have a direct bearing on that of the overlying keratinous sheath ( Fig. 7B View Figure 7 , kh); the same may be true in the case of the scelidosaur, but in the absence of new discoveries of scelidosaur material in the Lias exhibiting exceptional preservation (e.g. Arbour & Evans, 2017; Brown, 2017), there is no way of judging on the matter, so a conservative restoration has been illustrated.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.