
Kuronezumia endoi, Nakayama, Naohide, 2020
|
publication ID |
https://doi.org/ 10.11646/megataxa.3.1.1 |
|
DOI |
https://doi.org/10.5281/zenodo.6456295 |
|
persistent identifier |
https://treatment.plazi.org/id/B711B23F-FF30-86F8-D99D-C579FAFE7523 |
|
treatment provided by |
Plazi |
|
scientific name |
Kuronezumia endoi |
| status |
sp. nov. |
Kuronezumia endoi sp. nov.
[New Japanese name: Kuro-nezumi-dara]
( Figs. 152 View FIGURE 152 C–D, 153–156; Table 10 View TABLE 10 ; Appendix 3-9D)
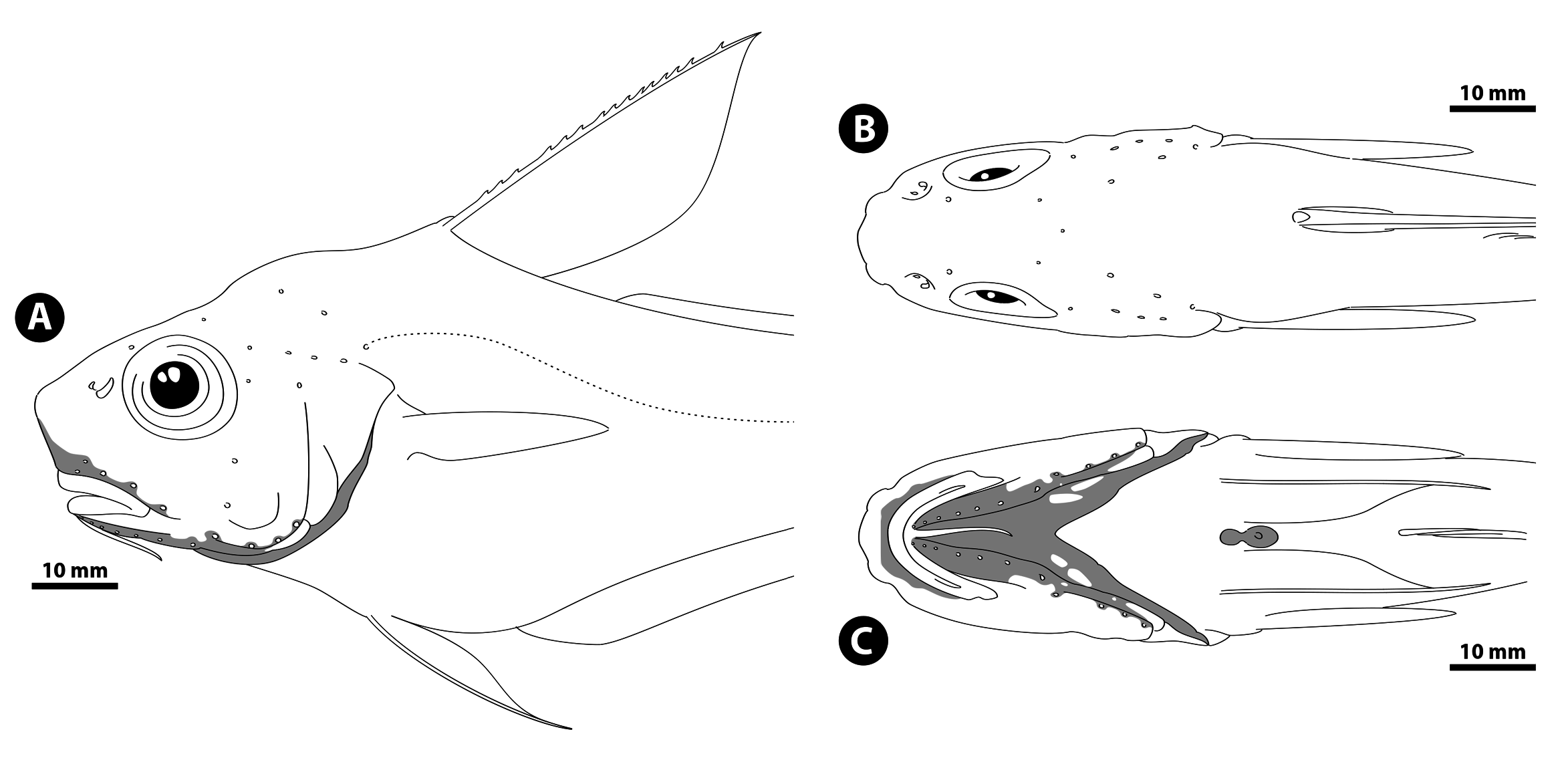
Diagnosis. A species of Kuronezumia without large, scaly, tubercular swelling anterior to anus; pelvic-fin rays 15; no enlarged scutes at snout tip; prominent open pores present along cephalic sensory canals; mandibular rami almost entirely naked except for posterior portions; scaly patches present on lower portions of branchiostegal rays; first dorsal fin blackish; oral cavity pale; body scales covered with long, moderately reclined, needle-like spinules in convergent rows; first dorsal-fin rays II,10–11; pectoralfin rays i22–i27; inner gill rakers on first arch 9; upperjaw length 31–32% HL.
Material examined. 2 specimens. Holotype: MSM 71-945 View Materials (32.7 mm HL, 215+ mm TL), 2 km off Fuji River , Shizuoka Pref., Suruga Bay, Japan, F/ V Asahi-maru, bottom trawl, coll. S. Kishimoto, 23 Dec. 1971 . Paratype: BSKU 44829 View Materials (1, 26.1 mm HL, 159+ mm TL), Tosa Bay , 33.1755ºN, 133.6498ºE, 500 m, FRV Kotaka-maru, 25 Apr. 1988 GoogleMaps .
Description. General features are shown in Figs. 153–155 View FIGURE 153 View FIGURE 154 View FIGURE 155 . Counts and measurements are given in Table 10 View TABLE 10 . The following description based on the holotype, unless otherwise stated; variations found in the paratype are indicated in parentheses. Trunk deep, compressed, width over pectoral-fin bases 2.1 (2.2) in depth at first dorsal-fin origin. Body deepest at first dorsal-fin origin, gradually tapering to long, slender tail. Head moderately large, HL about 6.6 (6.1) in TL. Supraoccipital crest moderately high, giving humpback appearance to dorsal contour of predorsal region (only slightly humped). Snout short, protruding slightly beyond upper jaw, length 0.9 (0.8) times as long as orbit diameter; ventral contour only slightly reclined from vertical when viewed laterally; snout high, its tip situated on horizontal through midorbit. Orbit moderate in size, circular, greatest diameter 1.4 (1.1) in postorbital length. Interorbital space slightly concave in preserved specimens, width 1.3 in orbit diameter. Mouth small, subinferior, upper-jaw length 1.0 (1.2) in orbit diameter; posterior margin of maxilla almost reaching vertical through midorbit (anterior 1/3 of orbit); lateral corner of mouth slightly restricted by lip folds; lips moderately thick, highly papillose near tooth bands. Suborbital region deep, only weakly ridged, lacking stout bony shelf. Preopercle large, posterior margin almost vertical, smoothly curved posteroventrally; preopercular ridge low, poorly marked, forming slight backward extension at angle. Interopercle narrowly exposed beyond preopercle. Gill membranes narrowly connected across, and attached mesially to isthmus, with slight posterior free fold. Gill opening moderately wide, extending forward to below posterior margin of lower jaw. Outermost gill slit moderately restricted by skin folds, length 2.2 (2.3) in orbit diameter. Gill rakers small, tubercular, armed with short, slender spines distally; those on outer side of first arch and inner side of fourth arch much smaller than others. Gill filaments moderately long. Barbel slender, well developed, length 1.4 (1.7) in orbit diameter.
Anus closer to pelvic-fin bases than to anal-fin origin. Periproct broad, teardrop-shaped, connected anteriorly to dermal window of light organ. Light organ small, circular, its anterior margin lying on line connecting outer pelvicfin bases.
Teeth short, slender, sharp, arranged in tapered bands in both jaws. Premaxillary band moderately narrow, with about 4 tooth rows anteriorly; outermost series distinctly enlarged; posterior margin of the tooth band falling far short of corner of mouth. Mandibular band broad but abruptly narrowing posteriorly, with about 5–6 tooth rows at widest point near symphysis; no teeth significantly enlarged.All teeth deeply embedded in thick layer of gum papillae.
Body scales very small, thin, not highly deciduous, covered with long, moderately reclined, needle-like spinules in about 9 convergent rows ( Fig. 152 View FIGURE 152 C–D) (5 subparallel rows); tip of last spinule in each row extending well beyond posterior scale margin; buttresses of spinules absent; reticulate structure broadly developed on posterior parts of unexposed portion. Body fully scaled except for periproct, light organ, and fins, but pelvic fin heavily scaled proximally.
Head ridges not prominently marked, without coarsely modified scales. Tip and lateral angles of snout lacking prominent scutes. Scales on head covered with cluster of short, erect, needle-like spinules. Dorsal and lateral surfaces of head completely and uniformly scaled except for narrow area around anterior nostril; underside of snout with broad, triangular naked area above upper lip ( Fig. 156A View FIGURE 156 ); mandibular rami almost completely naked except for small scaly patches on posterior portions ( Fig. 156C View FIGURE 156 ); gular membranes naked, but branchiostegal membranes with elongate scaly patches on bases of 4th and 5th branchiostegal rays respectively ( Fig. 156C View FIGURE 156 ).
Cephalic sensory pores present, serially arranged along sensory canals ( Fig. 156 View FIGURE 156 ); those on underside of head prominent and easily discernible. Grooved lateral line complete throughout its length.
First dorsal fin originating slightly posterior to vertical through pectoral-fin base; second spinous ray not especially elongate, its tip extending to base of 17th (22th) second dorsal-fin ray when laid back; height of first dorsal fin 3.4 (3.5) times as long as its base length; leading edge of second spinous ray armed with short, reclined, needlelike denticles (denticles much longer and more erect). Second dorsal fin moderately separated from first dorsal, originating above 9th (12th) anal-fin ray; interdorsal space 0.8 times as long as first dorsal-fin base length. Pelvic fin inserted slightly anterior to vertical through pectoral-fin base; outermost ray prolonged as hair-line filament, its tip reaching base of 6th (13th) anal-fin ray when laid back.
Color when fresh ( Fig. 153 View FIGURE 153 ). Uniformly dark; fins intensely blackish.
Color in alcohol ( Figs. 154–155 View FIGURE 154 View FIGURE 155 ). Head and body dark brown overall, with much darker abdomen; gular and branchiostegal membranes, dermal window of light organ, and periproct prominently blackish; lips dark; oral cavity and gum papillae pale; gill cavity paler anteriorly, blackish posteriorly; gill rakers, arches, and filaments yellowish brown; chin barbel generally immaculate, but slightly peppered near base; first dorsal fin dusky basally, but prominently black distally; second dorsal, pectoral, and anal fin uniformly dusky; pelvic fin darker. In paratype, ground color of head and body much darker than that in holotype; chest, abdomen, and gill apparatus distinctly blackish; gill rakers and arches with small melanophores densely scattered; first dorsal and pelvic fins uniformly black; pelvic fin blackish basally; anal fin darker anteriorly.
Size. To about 22 cm TL ( MSM 71-945 View Materials , holotype, 215+ mm TL, Suruga Bay , Japan) .
Distribution. So far known only from Suruga Bay and Tosa Bay at depths of about 500 m (Appendix 3-9D). Very rare.
Etymology. The new species is named in honor of Dr. Hiromitsu Endo of the Laboratory of Marine Biology, Faculty of Science and Technology, Kochi University (BSKU), for his contributions to our knowledge of gadiform systematics. The Japanese name, Kuro-nezumidara, is derived from a combination of the generic name (= Kuronezumia ) and a Japanese name for gadiform fishes (= dara).
Remarks, relationships, and comparisons. Data used in the following comparisons are based on Shcherbachev et al. (1992) and this study. Kuronezumia endoi sp. nov. is poorly represented in museum collections, with only two specimens being available. The new species is readily diagnosed within the genus by its higher counts of pelvic-fin rays (15 vs. ± 13 in other species, rarely 14 in K. bubonis Iwamoto, 1974 ). It further differs from all in that the mandibular rami are completely naked except for their very posterior portions ( Fig. 156C View FIGURE 156 vs. mostly or fully scaled). In addition, K. endoi differs notably from K. leonis ( Barnard, 1925) in the absence of a large spiny scute at the tip of the snout (vs. present); from K. dara ( Gilbert & Hubbs, 1916) and K. pallida Sazonov & Iwamoto, 1992 in the presence of the cephalic sensory pores ( Fig. 156 View FIGURE 156 vs. absent); from K. bubonis and an undescribed species from the South China Sea (sensu Iwamoto 1974; Shcherbachev et al. 1992) in the absence of a large tubercular swelling anterior to the anus (vs. present); and from all except K. leonis , K. pallida , and the above undescribed species in the presence of scaly patches on lower portions of the branchiostegal membranes ( Fig. 156C View FIGURE 156 vs. absent).
TABLE 10. (Continued)
It is putatively most similar to K. paepkei Shcherbachev, Sazonov & Iwamoto, 1992 known only from the holotype collected from the western Indian Ocean off Kenya. In addition to the above-mentioned differences, K endoi further differs from K. paepkei in having fewer gill rakers (9 vs. 12 in the inner side of the first arch; 8–9 vs. 11 on the outer side of the second arch), and smaller body scales (longitudinal scales 56–58 vs. 34–37; transverse scale rows below first dorsal-fin origin 19 vs. 13; below first dorsal-fin midbase 13–15.5 vs. 7; below second dorsal-fin origin 15–16 vs. 8).




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |