
Potamocarcinus darienensis, Magalhães, Célio, Campos, Martha R. & Türkay, Michael, 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3702.4.2 |
|
publication LSID |
lsid:zoobank.org:pub:C53B7547-58A9-4F2A-9FEC-0904FB34842E |
|
DOI |
https://doi.org/10.5281/zenodo.6158374 |
|
persistent identifier |
https://treatment.plazi.org/id/B7205608-FF8E-6874-A3F7-FC107F62D96A |
|
treatment provided by |
Plazi |
|
scientific name |
Potamocarcinus darienensis |
| status |
sp. nov. |
Potamocarcinus darienensis View in CoL n. sp.
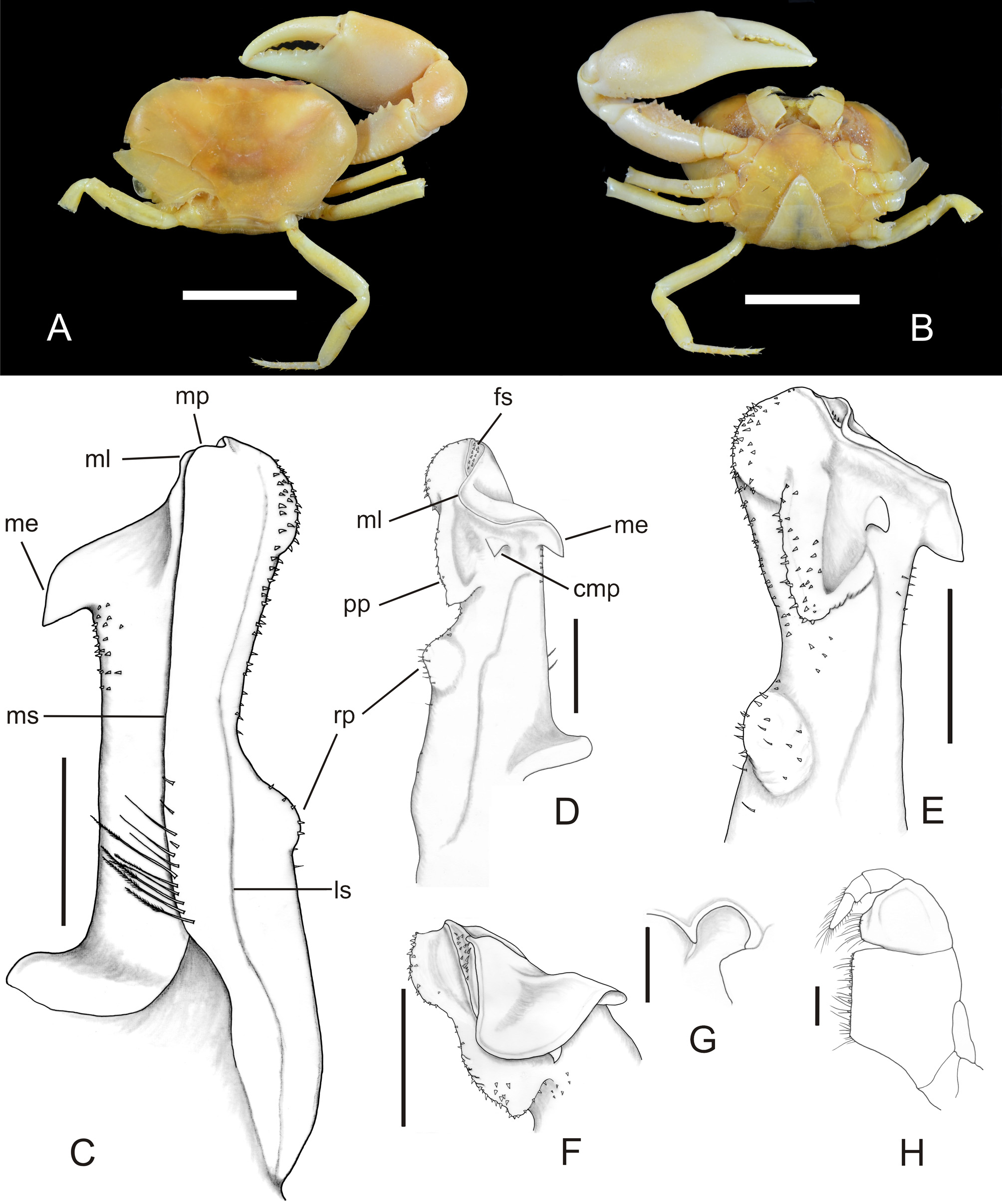
( Fig. 1 View FIGURE 1 A–H)
Holotype. Male (cw 21.0, cl 12.8, ch 8.4, fw 5.7), AMNH 19515 , Panamá, Darién Province, Cerro Malí, Serranía de Darién , 1400 m, on mountain top, 18.Jan.1975, Charles W. Myers leg.
Paratypes. 1 male (cw 18.5, cl 11.4), 1 female (cb 18.5, cl 11.2), AMNH 19516 , same data as holotype; 1 male (cw 20.5, cl 12.7), INPA 1983 , same data as holotype .
Diagnosis. Lateral border of G1 with rounded protuberance near median portion. Apex broadened, caudolateral surface with rounded protuberance, bearing minute spines. Mesial lobe short, oblique to main axis of apex, folded in latero-cephalic direction. Mesial process subtriangular, prominent, recurved cephalically, tip directed downwards; cephalic surface with a small, conical median cephalic process; large, prominent, subtriangular proximal cephalic process, with minute spines, scattered along its rounded margins; median cephalic process positioned between proximal cephalic and mesial processes. Apex cavity narrow, elongated caudo-cephalically, partially covered by folding of mesial lobe.
Description. Carapace outline ellipsoidal, widest in middle (mean cb/cl 1.63; n = 4); dorsal surface smooth, nearly flat, regions poorly defined. Two distinct gastric pits, close to each other on metagastric region. Cervical grooves wide, shallow, nearly straight, distal end failing to reach anterolateral margin. Postfrontal lobules obsolete; median groove indistinct between postfrontal lobules. Carapace surface between front and postfrontal lobules smooth, slightly inclined anteriorly. Upper border of front smooth, rounded; lower border carinate, nearly straight in frontal view. Upper orbital margin smooth, lower orbital margin marked by row of very faint papillae; exorbital angle obtuse. Anterolateral margins of carapace smooth, angled; posterolateral margins smooth, rounded. Epistome narrow; epistomial tooth very low, subtriangular, borders carinate. Suborbital, subhepatic regions of carapace sidewall smooth; pterygostomial regions covered by dense pubescence around mouthparts.
Endopod of third maxilliped ( Fig. 1 View FIGURE 1 H) with outer margin of ischium slightly convex, inner margin straight. Exopod long, 0.52 times length of outer margin of ischium, n = 4. Efferent branchial channel ( Fig. 1 View FIGURE 1 G) opening widely, with pyriform outline.
First pereopods markedly heterochelous (in adult paratype INPA 1983, right cheliped larger than left). Major cheliped with merus subtriangular in cross section; external border rounded, with irregular longitudinal row of tuberculiform teeth along proximal two thirds, smooth distally; internal upper border with row of conical teeth increasing in size distally; internal lower border delimited by row of rounded tubercles, distal border with row of rounded small tubercles. Carpus with row of tubercles, prominent median spine on inner side; outer side rounded, smooth. Palm distinctly swollen (length/breadth 1.47 to 1.51), smooth on both sides, with faint papillae on upper, lower rounded borders. Fingers usually gaping moderately when closed, tips not crossing; both fingers with large triangular teeth, smaller distally, sometimes interspersed with larger ones.
All abdominal segments free. Male telson subtriangular, its lateral margins nearly straight, tip rounded.
G1 ( Fig. 1 View FIGURE 1 C-F) robust, straight, wider at base; mesial border straight, with small patch of minute, scattered spines distally; lateral border convex basally in caudal view, with distinct rounded protuberance bearing scattered minute spines near midpoint; marginal suture situated on mesial side, straight along the whole stem, with proximal row of long (some plumose), short setae. Marginal process rudimentary, straight, slightly shorter than lateral border of apex. Lateral suture incomplete, marked by distinct sulcus on the proximal half of caudal side, by faint one to the distal half. Mesial lobe short, oblique to the main axis of apex, folded in latero-cephalic direction. Mesial process subtriangular, prominent, recurved cephalically, tip directed downward. Apex broadened, caudo-lateral surface with rounded protuberance bearing minute spines; apex cavity narrow, elongated caudo-cephalically, borders crested, mesial border slightly lower than lateral one. Field of apical spines weakly developed as elongated patch of few minute spines, partially covered by folding of mesial lobe. Distal portion of cephalic surface with small, conical median process, large, strong, subtriangular proximal process, with minute spines, scattered along its rounded margins; median process positioned between proximal cephalic, mesial processes. G2 as long as G1, very slender, tip tapering.
Etymology. The species is named after the region in which the type specimens were collected, Serranía de Darién, in the homonymous province.
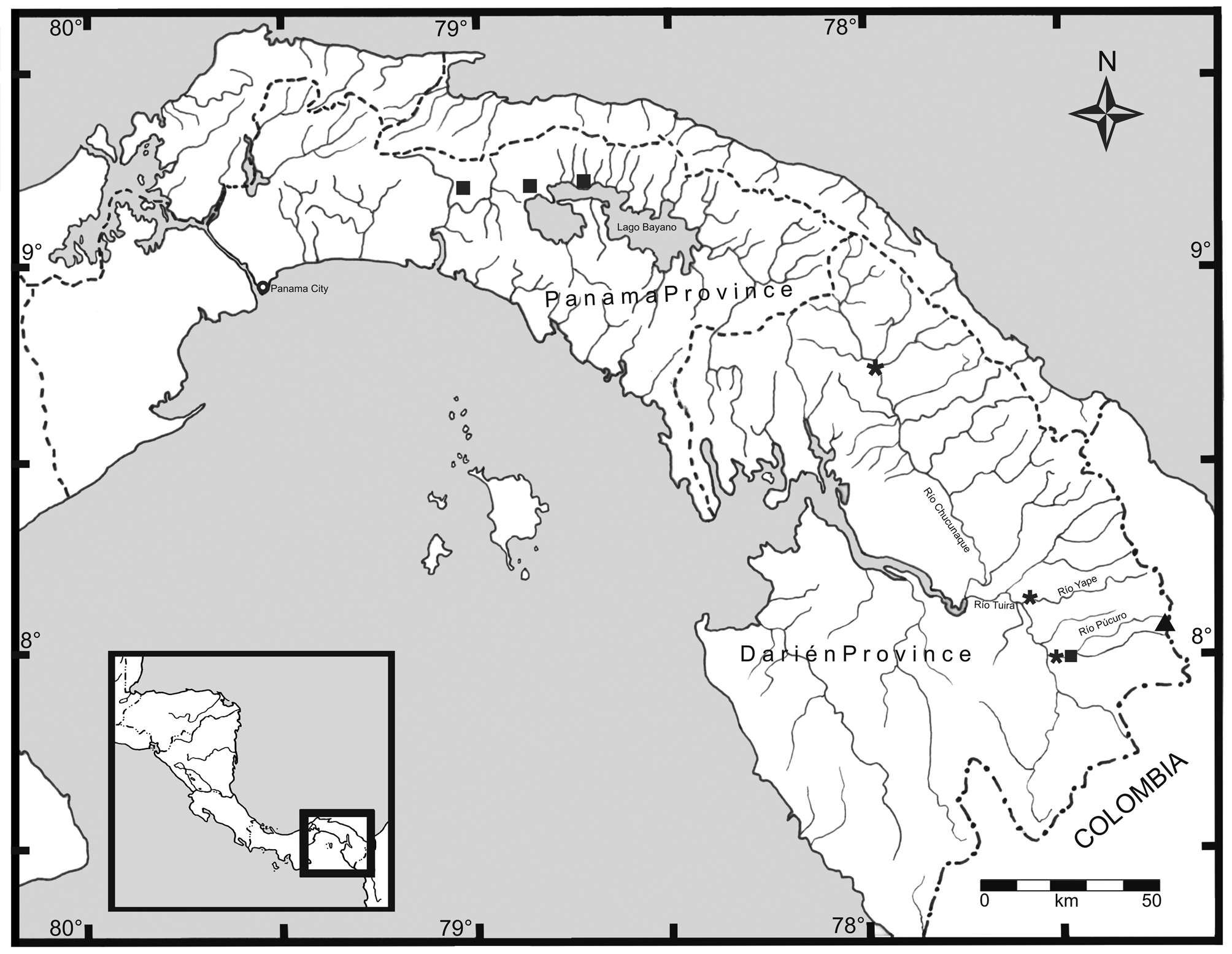
Type locality. Panamá, Darién Province, Serranía de Darién, Cerro Malí (08°07'N 77°14'W; Fairchild & Handley 1966) (see Fig. 3 View FIGURE 3 ).
Ecological notes. The specimens were "common under rocks—in the area of large rock outcrops" (C.W. Myers field notes), and they were collected near the top of Cerro Malí, a "ridge-top wet forest transitional to cloud forest". Myers (1969: 24-25) provided a detailed description of the physiography of the region and of the environment at Cerro Malí, and remarked that "Cerro Malí is high (about 1410 meters) and wet, but is affected by the dry season and much of its forest is possibly no more than transition to cloud forest". It was also noticed that there was no standing water in the immediate vicinity of the collection site (C.W. Myers, e-mail to first author, 1 March 2013).
Remarks. All specimens were disarticulated to some degree and some had a partially damaged carapace. The larger male specimen was chosen to be the holotype due to the size of the G1, although it only had the right cheliped (apparently the larger one) and the right P5 intact ( Fig. 1 View FIGURE 1 A, B); it also had a cleft along the left branchial region. The larger male paratype (INPA 1983) had both chelipeds, but the larger one (right) was detached from the body.
This new species closely resembles Potamocarcinus pinzoni Campos, 2003 . The main distinguishing feature between these two species is characters of G1. The apex of G1 of P. darienensis sp. n. is broadened by a rounded protuberance on its caudo-lateral surface (vs. apex without a rounded protuberance in P. pinzoni ); the lateral border of G1 has a rounded protuberance near the mid portion (vs. regularly rounded and devoid of a protuberance in P. pinzoni ); the cephalic surface of G1 has a small, conical median cephalic process, and a large, prominent, subtriangular proximal cephalic process (vs. a basally wide spine-shaped median cephalic process with median external notch, and the lack of a proximal cephalic process in P. pinzoni ).
The biogeographic importance of the Darién region in Panamá with respect to its fish fauna, was studied by Meek & Hildebrand (1916) and Hildebrandt (1938), who pointed out the similarity between the fish fauna of the Pacific slope of Panamá and that of the Atlantic slope of Colombia, particularly regarding the fishes of Tuira and Atrato rivers. Bermingham & Martin (1998), studying the phylogeography of representatives of three genera of primary freshwater fishes using mitochondrial genetic markers, also suggested a close relationship between the lower Central American fish fauna with that from northwestern Colombia. The morphological resemblance of P. darienensis sp. n. and P. pinzoni , a species from northwestern Colombia (Antioquia, Antioquia Department, Gulf of Urabá drainage on the Atlantic slope of Colombia) ( Campos 2003), rather than with the other species of Potamocarcinus that are distributed from central Panamá to southern Mexico, might be interpreted as further evidence of the faunal similarities between these regions.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.