
Lepidocyrtus (Setogaster) agricolus Mateos & Greenslade, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4981.2.9 |
|
publication LSID |
lsid:zoobank.org:pub:4BD9A224-FB20-4752-B489-1F8E50E1143D |
|
DOI |
https://doi.org/10.5281/zenodo.5046925 |
|
persistent identifier |
https://treatment.plazi.org/id/46D86A23-03E3-4F82-B03F-6D14F0BFA0E0 |
|
taxon LSID |
lsid:zoobank.org:act:46D86A23-03E3-4F82-B03F-6D14F0BFA0E0 |
|
treatment provided by |
Plazi |
|
scientific name |
Lepidocyrtus (Setogaster) agricolus Mateos & Greenslade |
| status |
sp. nov. |
Lepidocyrtus (Setogaster) agricolus Mateos & Greenslade sp. nov.
Figs 1–25 View FIGURES 1–7 View FIGURES 8–10 View FIGURES 11–12 View FIGURES 13–14 View FIGURES 15–17 View FIGURES 18–22 View FIGURES 23–25 , Tab 1 View TABLE 1
ZooBank: urn:lsid:zoobank.org:act:46D86A23-03E3-4F82-B03F-6D14F0BFA0E0
Type material. Holotype: female on slide ( SAMA 01-1450 About SAMA ), Australia, New South Wales, Harden , wheat fields, 416 m above sea level, S34º34’ E148º17’, pitfall traps, 1994/5, leg. M. Nash. 12 paratypes on slides: 4 females on the same slide as holotype (with separate codes SAMA 01-1451 About SAMA – 54 About SAMA ), 1 female and 2 of unknown sex on same slide ( SAMA 01-1455 About SAMA ), 2 females and 3 of unknown sex on same slide ( SAMA 01-1456 About SAMA ), all specimens with same data as holotype. GoogleMaps
Other material: 2 females and 13 of unknown sex on the slide SAMA 01-1457 About SAMA , 1 male and 14 of unknown sex on the slide SAMA 01-1458 About SAMA , 15 specimens of unknown sex on the slide SAMA 01-1459 About SAMA , 1 male and 7 of unknown sex on the slide SAMA 01-1460 About SAMA , 2 males and 3 of unknown sex on the slide SAM-028, Australia, New South Wales, Macquarie Valley, Auscott Warren cotton fields, 198 m above sea level, S31º47’ E147º46’, pitfall traps, 1994/5, leg. J. Lytton-Hitchins. All material deposited at the South Australian Museum (Adelaide, Australia) except slide SAM-028 deposited at the E. Mateos collection ( University of Barcelona , Barcelona, Spain) GoogleMaps .
Etymology. The specific name refers to the habitat of the new species being exclusively found so far only on broad acre arable agricultural land.
Diagnosis. Body without pigment; antenna, legs and dorsal side of manubrium unscaled; Ant.IV tip with T-chaeta; eyes G and H strongly reduced; labial chaetotaxy M 1 M 2 REL 1 L 2; dorsal macrochaetae formula A0A2aA2M2/00/01*00+3 (*short ciliated macrochaeta); without dorsal macrochaetae from Th.II to Abd.I; Abd.I without chaeta a6; Abd.II m3 short ciliated macrochaeta, m5 smooth microchaeta; Abd.IV trichobothria T2 and T4 close to each other, chaetae C1p, T3 and D1p in triangular pattern, chaeta a ciliated and bilobed, chaetae D1 and m ciliated and paddle-like, chaetae F2 and F3 ciliated macrochaete, with two psp on lateral position; unguis with a basal pair of teeth and two small inner teeth (apical tooth smaller); unguiculus lanceolate and with serrated outer margin; spatulate tibiotarsal tenent hair; dentes without basal tubercle; mucronal spine with spinelet.
Description. Adult body length (without head and furca) 0.8–1.3 mm (Holotype 0.93 mm). Th.II not projecting overhead ( Fig. 1 View FIGURES 1–7 ). Body dorsoventrally compressed; alcohol-preserved specimens with general body pigmentation pale fawn/yellow (no photograph available); densely black pigmented ocular areas.
Antenna without scales. Antennal length to head diagonal length ratio (head diagonal measured from cervical edge to apex of mouth part) 1.3–1.6. Relation of antennal joints I–IV as 1:1.8:1.9:3. Dorsal Ant.I-organ (three microchaetae arranged in triangle) present. Ant.III organ composed of two sub-cylindrical sensory rods partially covered by integumentary fold. Ant.IV with subapical mushroom-shaped chaeta (T-chaeta, Fig. 2 View FIGURES 1–7 ), without apical bulb.
Clypeus with four lateral chaetae (2 L1 and 2 L2), four facial chaetae (4f) in two rows, and three prefrontal chaetae (1 pf0 and 2 pf1), all these chaetae ciliated ( Fig. 3 View FIGURES 1–7 ). Labrum with ciliated prelabral chaetae and smooth labral chaetae in typical number 4/5,5,4; chaetae of apical row thicker than those in other rows; closed inverted V-shaped labral apical intrusion; labral papillae rounded and smooth ( Fig. 3 View FIGURES 1–7 ). Maxillary palp outer lobe with two subequal smooth chaetae, three smooth sublobal appendages, and a minute distal process ( Fig. 4 View FIGURES 1–7 ). Lateral process of outer labial papilla short, finger-shape, tip not reaching apex of papilla ( Fig. 5 View FIGURES 1–7 ). Labial and postlabial chaetotaxy as in Fig. 6 View FIGURES 1–7 ; with five smooth proximal chaetae at the base of labial palp; labial anterior row formed by five smooth chaetae (a1–a5); posterior row formed by ciliated chaetae with formula M 1 M 2 REL 1 L 2; chaeta R shorter, ratio M 2 /R = 2; postlabial chaetotaxy with all chaetae ciliated, without spines, 3 chaetae in row I (along ventral cephalic groove), 1 chaeta in row C, 3 chaetae in row E, 2 chaetae in row L, and 4 chaetae in row O.
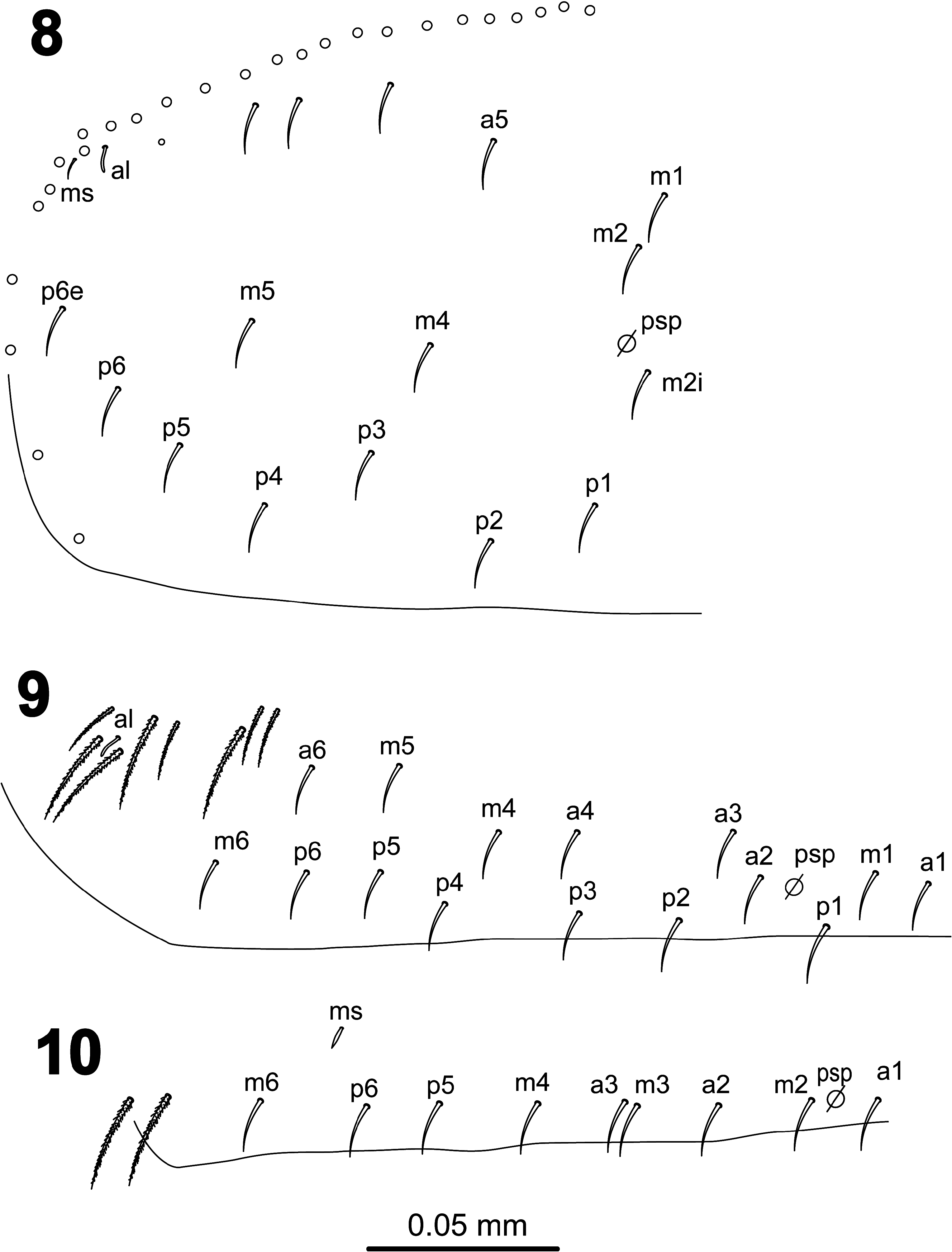
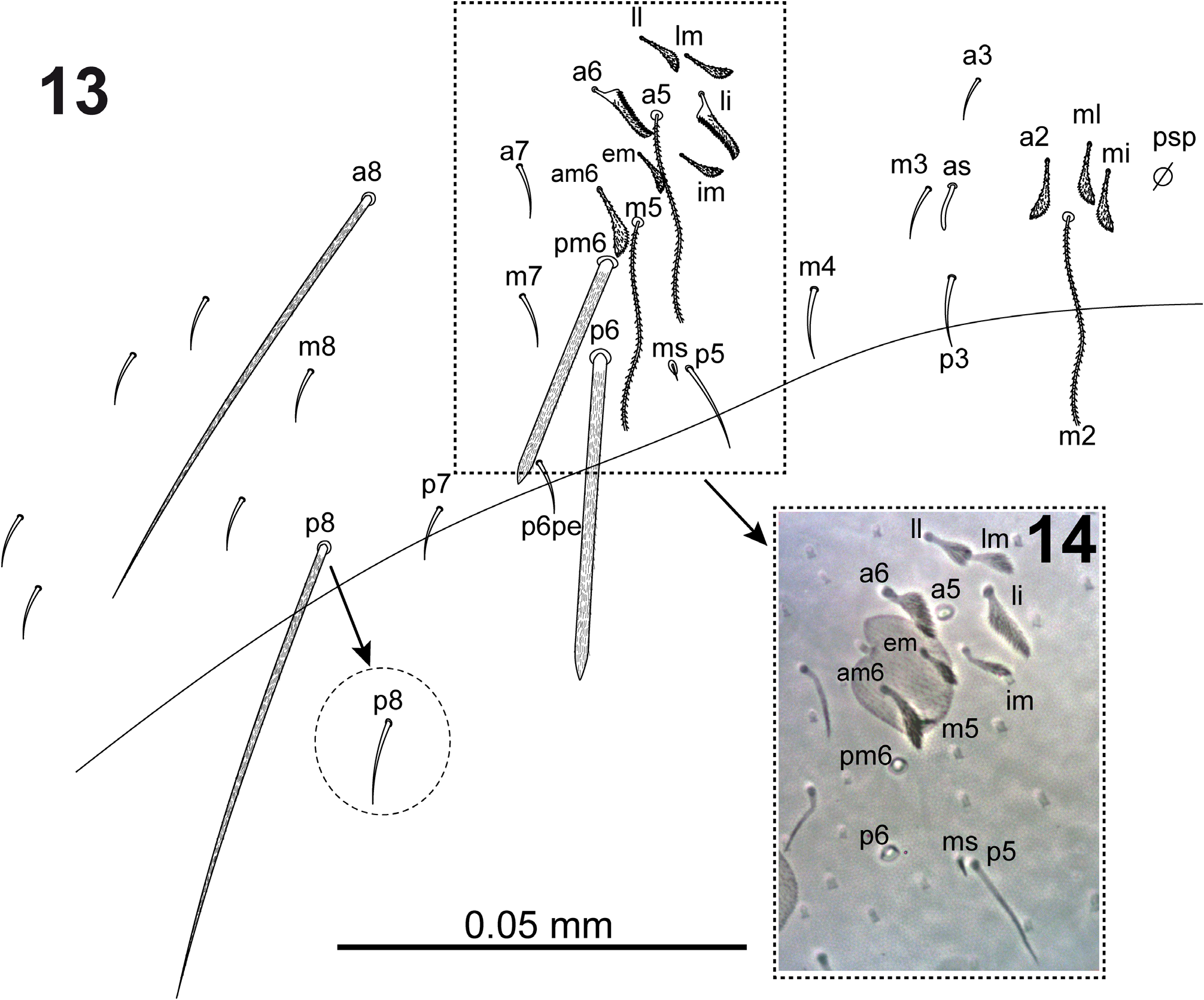
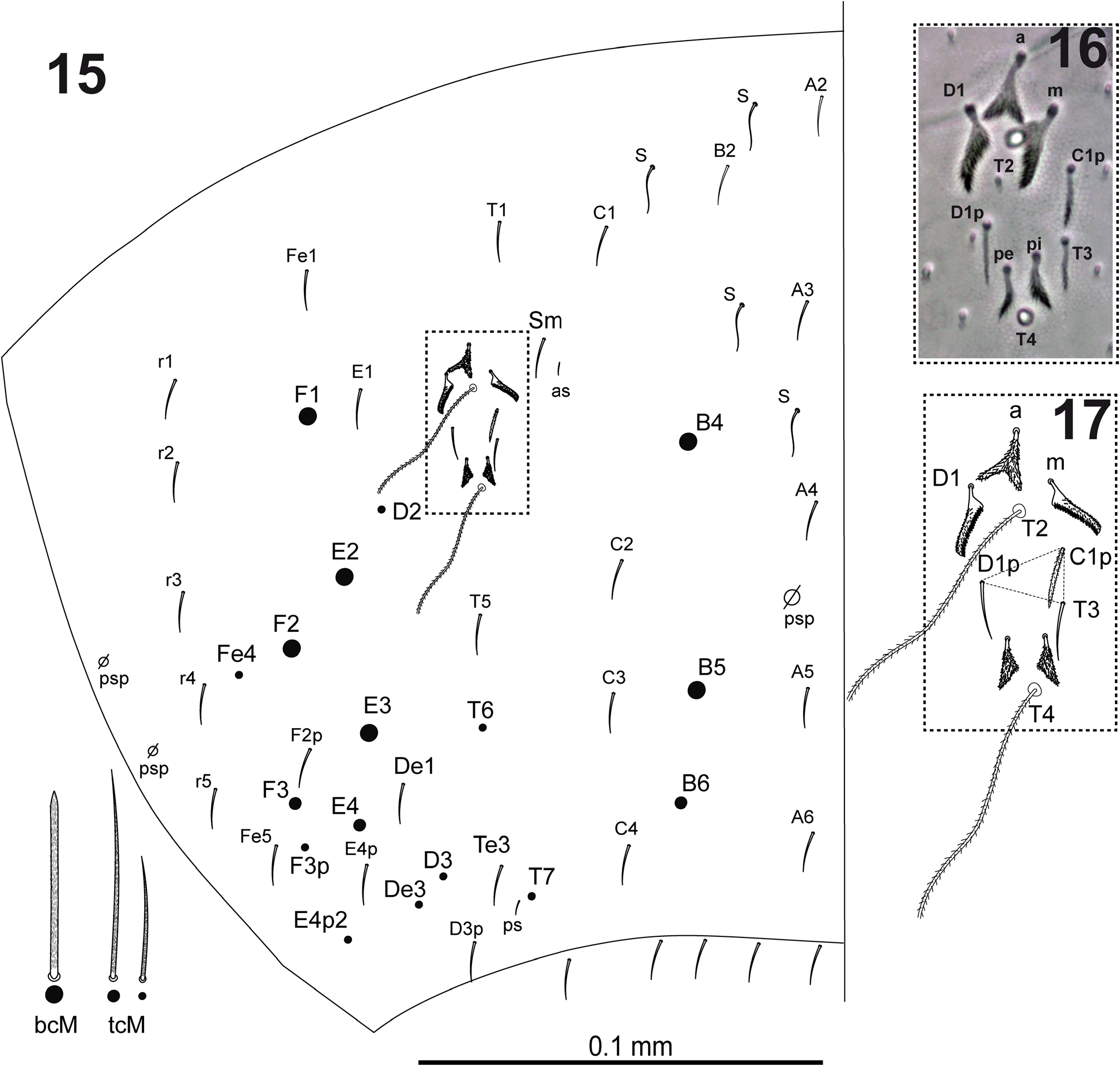
Dorsal head ( Fig. 7 View FIGURES 1–7 ) with macrochaetae A0, A2a, A2 and M2; A2a shorter than A2; one specimen (from slide SAMA 01-1456) with the two cephalic chaetae A2 as smooth microchaeta; chaeta pa5 absent. Interocular chaetotaxy with ciliated chaetae s, t, p, and 2 scales. Eyes G and H small and difficult to see on slides. Th.II–III and Abd.I dorsal chaetotaxy as in Figs 8–10 View FIGURES 8–10 . Th.II with 2 lateral S-chaetae (al and ms) and without macrochaetae in dorsal position. Th.III with a lateral sensillum (al) close to several ciliated chaetae. Abd.I with a lateral S-microchaeta (ms), chaetae a6 and m5 absent. Abd.II chaetotaxy as in Figs 11–12 View FIGURES 11–12 ; chaetae ml and a2p absent; chaetae mi, a2, lm and ll fan-shaped; chaeta m3 short ciliated macrochaeta; ratio m3/m3e = 1.7; m5 smooth microchaeta. Abd.III chaetotaxy as in Figs 13–14 View FIGURES 13–14 ; with S-chaetae as and ms; chaeta d3 absent; chaetae mi, ml, a2, li, lm, ll, a6, im, and em fanshaped, of which a6 and li larger (paddle-like); am6 fan-shaped chaeta; p6 and pm6 broad ciliated macrochaetae; a8 thin ciliated macrochaeta; p8 thin ciliated macrochaeta or smooth microchaeta; without lateral tuft of ciliated filaments. Abd.IV chaetotaxy as in Fig. 15 View FIGURES 15–17 ; Sm smooth microchaeta; B4 and B5 broad ciliated macrochaetae, B6 thin ciliated macrochaeta with socket of smaller diameter than macrochaetae B4 and B5; F2 broad ciliated macrochaeta; F3 thin ciliated macrochaeta; Fe4 thin ciliated macrochaeta or smooth microchaeta; with 4+4 dorsal long S-chaetae on anterior region; with two lateral psp located external to chaeta r4. In the Abd.IV anterior bothriotrichal complex ( Figs 16–17 View FIGURES 15–17 ) the ratio of distances between T2–T4/C1p ≈ 3; bothriotrichum T2 without accessory chaeta s (one specimen with a supplementary ciliated chaeta between chaetae a and m); chaetae C1p, D1p and T3 forming a triangle; chaeta C1p finelly cilated; chaetae m, D1, pi and pe fan-shaped, from which m and D1 larger (paddle-like); chaeta a bilobed and cliated. Abd.V ( Fig. 18 View FIGURES 18–22 ) with dorsal S-chaetae as, acc.p4 and acc.p5.
Ventral tube without scales, with 10+10 ciliated chaetae on anterior side ( Fig 19 View FIGURES 18–22 ) and 9 ciliated chaetae on posterior side; each lateral flap with maximum of 6 ciliated chaetae and 3 smooth chaetae.
Legs without scales. V-shaped trochanteral organ (leg III) with maximum of 12 smooth straight chaetae arranged in triangular shape ( Fig. 20 View FIGURES 18–22 ). Unguis ( Fig. 21 View FIGURES 18–22 ) with well-developed basal pair of teeth at 55% from base of the inner edge, and with two small inner teeth at 73% and 87% from base of inner edge, respectively (apical tooth smaller); unguiculus lanceolate and with serrated outer margin; spatulate tibiotarsal tenent hair and acuminate supra-empodial chaeta (smooth chaeta on tibiotarsus III opposite to tenent hair); ratio tenent hair/supra-empodial chaeta ≈ 1.7; ratio unguis inner edge/tenent hair ≈ 1.1.
Manubrium without scales on dorsal surface; ventro-apical end with 2+2 ciliated chaetae; manubrial plate with two psp, 2–3 inner chaetae, and 2 outer chaetae. Dentes without basal tubercle. Mucronal basal spine with spinelet ( Fig. 22 View FIGURES 18–22 ).
Pseudopores distribution ( Figs 23–24 View FIGURES 23–25 ) as below:
- Antenna: 1 on ventral side of Ant.II and Ant.III, located distally on the membranous area between the chaetae of the apical row and the tip of the antennal joints.
- Head: 1+1 located anteriorly to each eye patch (dorsoexternally to Ant.I), and 2+2 located between antennal bases (dorsointernally to each Ant.I).
- Dorsal body: 1+1 on each segment form Th.II to Abd.IV (near the central axis of the segments).
- Lateral body: 2+2 laterally on Abd.IV ( Fig. 25 View FIGURES 23–25 ).
- Ventral body: 1+1 on each thoracic segment (near the base of coxae), 1+1 anteriorly to the base of the ventral tube and 1+1 posteriorly to the base of the ventral tube, a group of 2 between the base of ventral tube and retinaculum, 2 in a line posterior to retinaculum, and 1 anterior to genital plate on Abd.V.
- Legs: 2 on Cx.I; 3–4 on Cx.Cx.II; 2 on Cx.III.
- Furca: 1+1 on dorsal manubrial base; 2 on each dorsal manubrial plate; 1 on each dorsal dens base (within the first basal crenulations).
Discussion. The subgenus Setogaster is characterised by the dorso-basal region of the dentes with a rounded tubercle and lacking spines, absence of apical bulb on Ant.IV, and presence of spinelet on the basal mucronal spine ( Cipola et al. 2019). The new species lacks dental tubercle, as well as L. (S.) fasciatus ( Salmon, 1951) , but presents the other characteristics that define Setogaster , which is why it has been assigned to this subgenus (see Mateos & Greenslade 2015). Lepidocyrtus (Setogaster) agricolus sp. nov. is close to species Lepidocyrtus (Setogaster) coorongensis sp. nov., Lepidocyrtus (Setogaster) fasciatus ( Salmon, 1951) sensu Mateos & Greenslade, 2015 , Lepidocyrtus (Setogaster) kuakea Christiansen & Bellinger, 1992 , Lepidocyrtus (Setogaster) nashi sp. nov., Lepidocyrtus (Setogaster) nigrofasciatus Womersley, 1934 , Lepidocyrtus (Setogaster) praecisus Schött, 1917 and Acrocyrtus zhujiensis Xu, Pan & Zhang, 2013 . Body pigmentation, dorsal body chaetotaxy, labial chaetotaxy, foot complex morphology (unguis and unguiculus) and dental tubercle shape and development are useful characters that allow differentiating these species ( Table 1 View TABLE 1 ). Body without pigment in L. (S.) agricolus sp. nov. clearly separates this species to the pigmented L. (S.) fasciatus , L. (S.) nashi sp. nov., L. (S.) nigrosetosus , L. (S.) praecisus , and A. zhujiensis . Abd.IV F2 and F3 as ciliated macrochaetae in L. (S.) agricolus sp. nov. separates this species to L. (S.) coorongensis sp. nov. and L. (S.) kuakea (with Abd.IV F2 and F3 as smooth microchaetae).
The new species was trapped in large numbers, up to 75 individuals per pitfall trap day at times, in cotton fields in northern New South Wales. On one site, it was virtually the only species trapped and was most abundant under cotton trash in furrows between plants ( Lytton-Hitchins et al. 2015) and survived several applications of pesticide to the crop. Numbers plummeted after irrigation. It was recorded by Lytton-Hitchins et al. (2015) as Setogaster spp. In wheat fields, nearly 600 km further south, it was also only collected in pitfalls and not from soil samples but it was much less abundant and only found in numbers in October 1994 when immatures were also present. More than twice as many individuals were trapped on direct drilled wheat plots than on conventionally tilled plots.




No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |