
Lepidocyrtus (Setogaster) nashi Mateos & Greenslade, 2021
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4981.2.9 |
|
publication LSID |
lsid:zoobank.org:pub:4BD9A224-FB20-4752-B489-1F8E50E1143D |
|
DOI |
https://doi.org/10.5281/zenodo.5046929 |
|
persistent identifier |
https://treatment.plazi.org/id/B8062D31-0B0B-4F0B-971F-EB5EC32ECEF8 |
|
taxon LSID |
lsid:zoobank.org:act:B8062D31-0B0B-4F0B-971F-EB5EC32ECEF8 |
|
treatment provided by |
Plazi |
|
scientific name |
Lepidocyrtus (Setogaster) nashi Mateos & Greenslade |
| status |
sp. nov. |
Lepidocyrtus (Setogaster) nashi Mateos & Greenslade sp. nov.
Figs 23, 24 View FIGURES 23–25 , 35–47 View FIGURES 35–40 View FIGURES 41–43 View FIGURES 44–47 , Tab 1 View TABLE 1
ZooBank: urn:lsid:zoobank.org:act:B8062D31-0B0B-4F0B-971F-EB5EC32ECEF8
Type material. Holotype: female on two slides (head and body separated, code T 22517), Australia, Victoria State, Tagels, Mt Bogong , 1868 m above sea level, S36°41’21” E147°18’24”, in pitfalls under snow in alpine grassland, 14.i.2012, M. Nash leg. GoogleMaps Paratypes: one male on slide code T 22518 , 4 specimens of unknown sex on slide codes LP307-1 , LP307-2 , LP307-4 , LP307-5 , and 2 specimens on absolute alcohol (sample code LP307 ), same data as holotype GoogleMaps . Holotype and paratype slide T 22518 deposited at the Museum of Victoria ( Melbourne , Australia); other paratype deposited at the E. Mateos collection (University of Barcelona, Barcelona, Spain) .
Etymology. The new species was named after the collector, Michael Nash who has been assiduously collecting Collembola for the junior author for many years.
Diagnosis. With blue colour present on Ant.I–IV, dorsal and ventral head, Th.II to Abd.IV and coxae, trochanters and femora of three pair of legs; antenna, legs and dorsal side of manubrium unscaled; Ant.IV tip with T-chaeta; eyes G and H strongly reduced; labial chaetotaxy M 1 M 2 rEL 1 L 2;dorsal macrochaetae formula A0A2aA2/00/01*00+3 (*short ciliated macrochaeta); without dorsal macrochaetae from Th.II to Abd.I; Abd.I with chaeta a6; Abd.II m3 short ciliated macrochaeta, m5 smooth microchaeta; Abd.III with lateral tuft of 20–25 long ciliated filaments; Abd. IV trichobothria T2 and T4 close to each other, chaetae C1p, T3 and D1p in triangular pattern, chaeta a ciliated and bilobed, chaetae D1 and m ciliated and paddle-like, chaetae F2 and F3 ciliated macrochaete, with two psp on lateral position; unguis with a basal pair of teeth and two small inner teeth; unguiculus lanceolate and with serrated outer margin; spatulate tibiotarsal tenent hair; dentes with small, rounded basal tubercle; mucronal spine with spinelet.
Description. Adult body length (without head and furca) 1.0– 1.3 mm (Holotype 1.3). Mesothorax not projecting overhead. Body dorsoventrally compressed. Blue colour present on Ant.I–IV, dispersed spots on dorsal and ventral head, on Th.II to Abd.III, mid-dorsal band on Abd.IV, and coxae, trochanters and femora of three pair of legs; densely black pigmented ocular areas ( Fig. 35 View FIGURES 35–40 ).
Antenna without scales. Antennal length to head diagonal length ratio (head diagonal measured from cervical edge to apex of mouth part) ≈ 1.4. Relation of antennal joints I–IV as 1:1.6:1.7:2.7. Dorsal Ant.I-organ present. Ant. III organ composed of two subcylindrical sensory rods partially covered by an integumentary fold. Ant.IV with subapical mushroom-shaped chaeta (T-chaeta), without apical bulb.
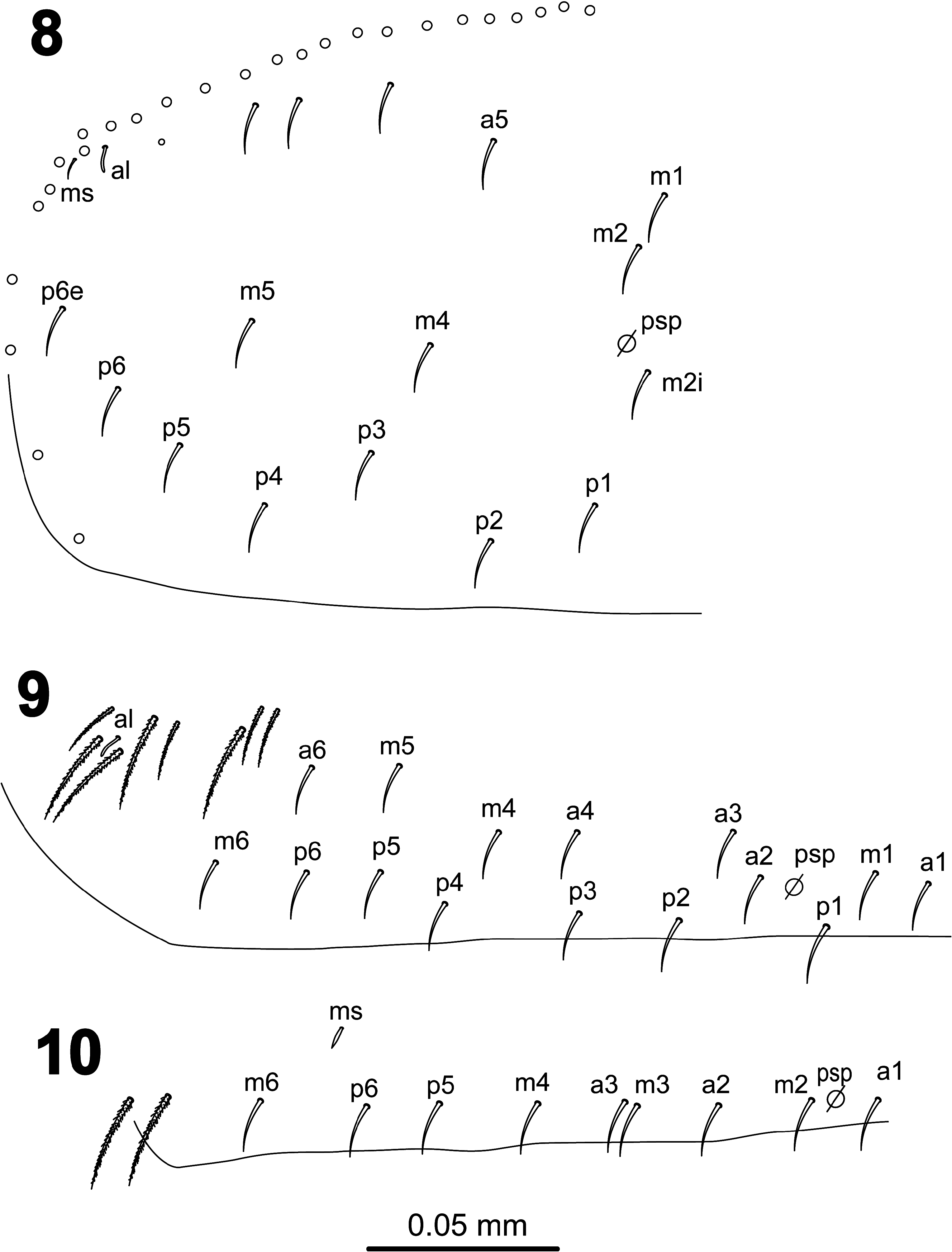
Clypeus and labrum as L. agricolus sp. nov. ( Fig. 3 View FIGURES 1–7 ). Clypeus with four lateral chaetae (2 L1 and 2 L2), four facial chaetae in two rows, and three prefrontal chaetae (1 pf0 and 2 pf1), all these chaetae ciliated. Labrum with ciliated prelabral chaetae and smooth labral chaetae in typical number 4/5,5,4; chaetae of apical row thicker than those in other rows. Closed inverted V-shaped labral apical intrusion; labral papillae smooth. Maxillary palp outer lobe with two subequal smooth chaetae and four smooth sublobal appendages, the tiny distal process that is present in the other two species described above (see Fig. 5 View FIGURES 1–7 ) has not been observed ( Fig. 36 View FIGURES 35–40 ). Lateral process of outer labial papilla short, finger-shape, tip not reaching apex of papilla ( Fig. 37 View FIGURES 35–40 ). Labium ( Figs 38–39 View FIGURES 35–40 ) with five smooth proximal chaetae at the base of labial palp; labial anterior row formed by five smooth chaetae (a1–a5); posterior row formed by ciliated chaetae with formula M 1 M 2 rEL 1 L 2; r vestigial smooth microchaeta, ratio M 2 /r = 9; postlabial chaetotaxy as in L. agricolus sp. nov. ( Fig. 6 View FIGURES 1–7 ), with all chaetae ciliated, without spines, 3 chaetae in row I (one paratype asymmetric, with 3+4 ciliated chaetae on this position), 1 chaeta in row C, 3 chaetae in row E, 2 chaetae in row L, and 4 chaetae in row O.
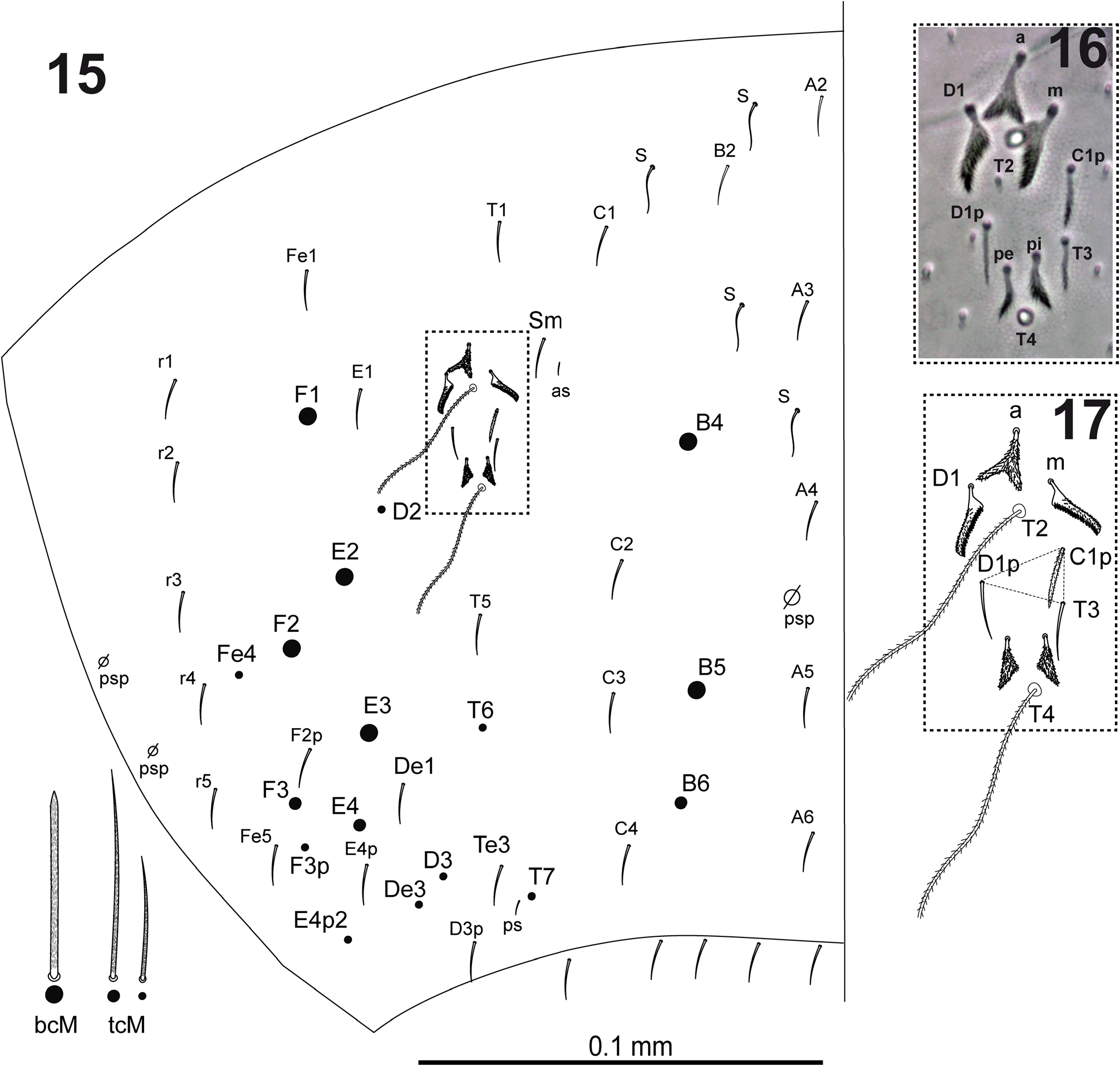
Dorsal head ( Fig. 40 View FIGURES 35–40 ) with macrochaetae A0, A2a and A2; A2a length equal or longer than A2; chaeta Pa5 absent.Interocular chaetotaxy with ciliated chaetae s, t, p, and 3 scales; eyes G and H small and difficult to see on the slides. Th.II–III dorsal chaetotaxy as L. agricolus sp. nov. ( Figs 8–9 View FIGURES 8–10 ). Th.II with 2 lateral S-chaetae (al and ms) and without macrochaetae in dorsal position. Th.III with a lateral sensillum (al) close to several ciliated chaetae. Abd.I dorsal chaetotaxy as in Fig. 41 View FIGURES 41–43 , with chaeta m5 and a lateral S-microchaeta (ms) external to a6. Abd.II chaetotaxy as in Fig. 42 View FIGURES 41–43 ; chaetae ml and a2p absent; chaetae mi, a2, ml and ll fan-shaped; chaeta m3 short ciliated macrochaeta, ratio m3/m3e = 1.2; chaeta a6 present; m5 smooth microchaeta. Abd.III chaetotaxy as in Fig. 43 View FIGURES 41–43 ; with S-chaetae as and ms; chaeta d3 present on one specimen, absent on the rest; chaetae mi, ml, a2, li, lm, ll, a6, im, em and am6 fan-shaped, of which a6 and li larger (paddle-like); p6 and pm6 broad ciliated macrochaetae; a8 and p8 thin ciliated macrochaetae; with lateral tuft of 20–25 long ciliated filaments. Abd.IV chaetotaxy as in Fig. 44 View FIGURES 44–47 ; Sm smooth microchaeta; B4 and B5 broad ciliated macrochaetae, B6 thin ciliated macrochaeta with socket of minor diameter than macrochaetae B4 and B5; F2 broad ciliated macrochaeta; F3 thin ciliated macrochaeta; Fe4 thin ciliated macrochaeta; with 4+4 dorsal long S-chaetae on anterior region; with two lateral psp located external to chaeta r4. In the Abd.IV anterior bothriotrichal complex the ratio of distances between T2–T4/C1p ≈ 3; bothriotrichum T2 without accessory chaeta s; chaetae C1p, D1p and T3 forming a triangle (as in L. agricolus sp. nov., Fig 17 View FIGURES 15–17 ); chaeta C1p strongly ciliated (Holotype) or finelly cilated (Paratypes), chaetae m, D1, pi and pe fan-shaped, from which m and D1 larger (paddle-like); chaeta a bilobed and cliated. Abd.V ( Fig. 45 View FIGURES 44–47 ) with dorsal S-chaetae as, acc.p4 and acc.p5.
Ventral tube without scales, with 10+10 ciliated chaetae on anterior side and 9 ciliated chaetae on posterior side (as in L. agricolus sp. nov., Fig 19 View FIGURES 18–22 ); all chaetae on posterior side ciliated; each lateral flap with maximum of 6 ciliated chaetae and 6 smooth chaetae.
Legs without scales. V shaped trochanteral organ (leg III) with maximum of 15 smooth straight chaetae arranged in triangular shape ( Fig. 46 View FIGURES 44–47 ). Unguis ( Fig. 47 View FIGURES 44–47 ) with well-developed basal pair of teeth at 42% from base of the inner edge, and with two small inner teeth at 67% and 84% from base of inner edge, respectively (apical tooth smaller); unguiculus lanceolate and with serrated outer margin; spatulate tibiotarsal tenent hair and acuminate supra-empodial chaeta; ratio tenent hair/supra-empodial chaeta ≈ 2; ratio unguis inner edge/tenent hair ≈ 1.
Manubrium without scales on dorsal surface; ventro-apical end with 2+2 ciliated chaetae; manubrial plate with two psp, 3 inner chaetae, and 3 outer chaetae. Dentes with small, rounded basal tubercle (difficult to see in several specimens). Mucronal basal spine with spinelet (as in L. agricolus sp. nov., Fig. 22 View FIGURES 18–22 ).
Pseudopores distribution as in L. (S.) agricolus sp. nov. ( Figs 23–24 View FIGURES 23–25 ).
Discussion. Lepidocyrtus (Setogaster) nashi sp. nov. is close to the other two new species described, L. (S.) agricolus sp. nov. and L. (S.) coorongensis sp. nov., and to the other species listed in Table 1 View TABLE 1 . Body with blue pigment and labium with smooth vestigial microchaeta r are good diagnostic characters differentiating L. (S.) nashi sp. nov. from species L. (S.) agricolus sp. nov., L. (S.) coorongensis sp. nov. and L. (S.) kuakea ( Table 1 View TABLE 1 ). Acuminate unguiculus in L. (S.) nashi sp. nov. clearly separates this species to the other pigmented species listed in Table 1 View TABLE 1 .
The new species appears to be locally endemic and restricted to alpine grassland habitat on Mt Bogong where it is active under snow. The plant species present in the area were Celmisia costiniana (70%), Poa fawcettiae (20%), Luzula acutifolia (5%), and assorted herbs (5%) including Trachymene humilis and Aciphylla glacialis . A number of other invertebrates have been described as restricted to this habitat and locality ( Greenslade & Slatyer 2017, Braby & Wurtz 2018). Rocky crevices here are an aestivation site for the Bogong Moth, an important food source for the Mountain Pygmy Possum. This fauna is being impacted adversely by climate change resulting in reduced snow cover as well as by increasing tourism. There is anecdotal evidence of visitors interfering with aestivating moths.






| T |
Tavera, Department of Geology and Geophysics |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |