
Cyclops ankyrae Mann, 1940
|
publication ID |
https://doi.org/ 10.1080/00222930802140160 |
|
persistent identifier |
https://treatment.plazi.org/id/B95087A2-0D74-FFB4-FE53-FAF2FB91222C |
|
treatment provided by |
Felipe |
|
scientific name |
Cyclops ankyrae Mann, 1940 |
| status |
|
Cyclops ankyrae Mann 1940: p.65 –67, Figure 18; Einsle 1992: p.183, Figures 5,6; Einsle 1996a: p.14 –16, Figure 7 View Figure 7 .
Cyclops furcifer Claus : Kiefer 1955: p.108, p.114–117, Figures 9 View Figure 9 –13.
Material examined
Cyclops ankyrae : Turkey, Emirgân, 20 February 1939, two ♀♀, ten „„ (two „„ dissected and eight „„ undissected), one CV ♀ ( SMNK, alcohol material ‘‘Glas’’: 1391) ; Italy, Latium, Wood of Foglino , 41 ° 289100N 12 ° 439470E, temporary puddle, F. Stoch & D. Vagaggini 28 November 2004, five ♀♀ ( MIZ: 157864-68), two „„ ( MIZ: 157869-70)
Comparative material
Cyclops furcifer . Germany, Eichener See (Baden), seven ♀♀, three „„, leg. F. Kiefer 13 April 1935 ( SMNK: 2708-2709, 2716-2717). ‘‘ Cyclops lacunae ’’ Lowndes : United Kingdom, Marlborough District , A. G. Lowndes 25 May 1926, three ♀♀ (syntypes, NHM) .
Cyclops vicinus . Hungary, Szarvas, Fisheries Research Institute: fishpond B1, leg.
M. Hołyńska 9 May 1990, one ♀ ( MIZ: 157871); fishpond B6, 46 ° 529N 20 ° 339E, leg. M. Hołyńska 29 May 1992, one ♀ ( MIZ: 157872). Poland Wojewódzkie Mazowieckie, Nowa Wieś ca. 20 km southwest of Warsaw , [3], 52 ° 089N 20 ° 469E, leg. R. & M. Hołyńska 3 April 2006, one ♀ (undissected) ( MIZ) . South Korea, Gyeongsan, Muncheon Reservoir, 35 ° 539490N 128 ° 509430E, leg. C. Y. Chang 24 December 2003, one ♀ ( MIZ: 157873) .
Cyclops kikuchii . Japan, Hokkaido, Yoichi, pond, 43 ° 119N 140 ° 479E, leg. C. Y.
Chang 4 April 2005, two ♀♀ ( MIZ: 157874-75); South Korea, Changnyeong, Shinjeon Reservoir, 35 ° 249430N 128 ° 279530E, leg. C. Y. Chang 2 February 2005, two ♀♀ ( MIZ: 157876-77) .
Cyclops glacialis . Mongolia, Turgen-Charchiraa Mountains, Aimak Uvs, Nogoon Nuur , 49 ° 499N 91 ° 169E, 14 August 1999, two ♀♀ , two „„ one CV ♀ (paratypes, ZMB: 27426) .
Cyclops insignis . Norway, Frognertjern, 60 ° 489N 11 ° 109E, leg. S. B. Waervågen 20 February 1995, two ♀♀ ( MIZ: 157878-79) .
Description
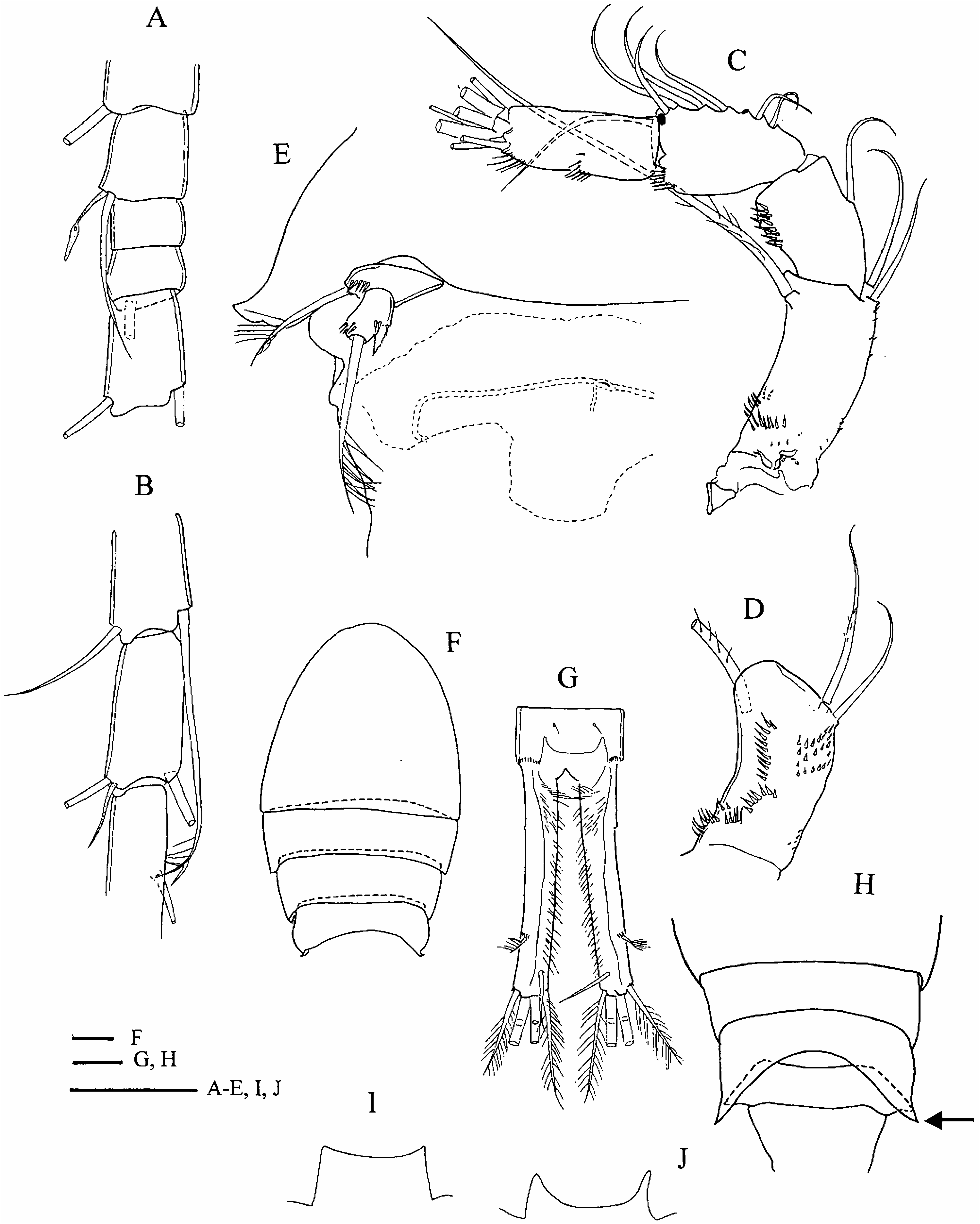
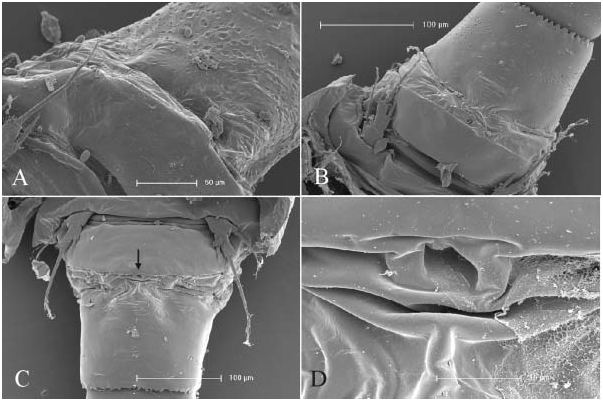
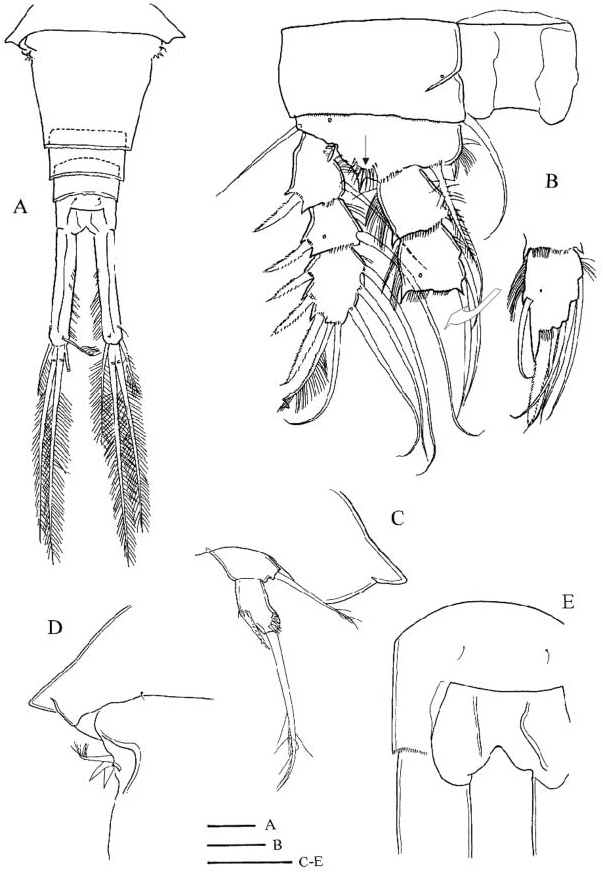
Female ( Turkey and Italy). Morphometric data are given in Table 1. Posterior angle of pediger 4 ( Figure 1F View Figure 1 ) not prolonged (no ‘‘wings’’) (cf. Figure 1H View Figure 1 , ‘‘wings’’ arrowed). Pediger 5 ( Figure 1E View Figure 1 ) pointed outwards but without lateral ‘‘wings’’, segment emarginated and only slightly wider than genital double-somite (for pediger 5 with ‘wings’ see Figure 1H View Figure 1 ), lateroventral lobes absent ( Figure 1E View Figure 1 , cf. C. abyssorum divergens in Figure 6A View Figure 6 ). Genital double-somite gradually or abruptly narrowing posterior to seminal receptacle. No cuticular ridges near copulatory pore on ventral surface of genital double-somite ( Figure 1E View Figure 1 , cf. C. abyssorum divergens in Figure 7 View Figure 7 ). Anal operculum ( Figure 1G,J View Figure 1 ), in comparison to congeners, large and rounded. Proctodeum without hair ornamentation. Posterior margin of anal somite with continuous row of spinules. Caudal ramus ( Figure 1G View Figure 1 ) with distinct longitudinal keel on dorsal surface. Medial margin pilose; at anterior end tuft of hairs set perpendicular to the ramus axis. Lateral caudal seta inserted at distal quarter. Spinules present at implantation of lateral and lateralmost terminal caudal setae.
Antennule 17-segmented, reaching beyond cephalothorax, but not reaching posterior margin of pediger 2. Last three terminal segments with hyaline membrane. Armature formula: 8, 4, 2, 6, 4, 1+spine, 2, 1, 1, 0, 1, 1+aesthetasc, 0, 1, 2, 2+aesthetasc, 7+aesthetasc. Aesthetasc on segment 12 ( Figure 1A View Figure 1 ) (29–36 Mm; seven ♀♀ measured) reaching about middle of segment 14, aesthetasc on segment 16 ( Figure 1B View Figure 1 ) (25–35 Mm; seven ♀♀ measured) not reaching insertion of medial seta of segment 17. Antennule adorned with spinules on anterior surface of first segment, and cuticular pits on posterior surface of more proximal segments. Antenna ( Figure 1C View Figure 1 ) bearing 3 setae on coxobasis, and one, nine and seven setae on endopodal segments 1, 2 and 3, respectively. Lateral seta of coxobasis, remnant of exopodite, reaching beyond distal margin of third endopodal segment. Medial setae of coxobasis without long setules. Caudal ornamentation of coxobasis ( Figure 1D View Figure 1 ) composed of long spinules on lateral margin near base, longitudinal row of elongate spinules, double oblique row proximal to longitudinal row, oblique field of small spinules below insertion of medial setae, and tiny spinules on medial margin near base. Frontal surface of coxobasis ( Figure 1C View Figure 1 ) ornamented with few spinules near lateral margin, at 4/10 of segment, and transverse rows of long and small spinules medioproximally to former group.
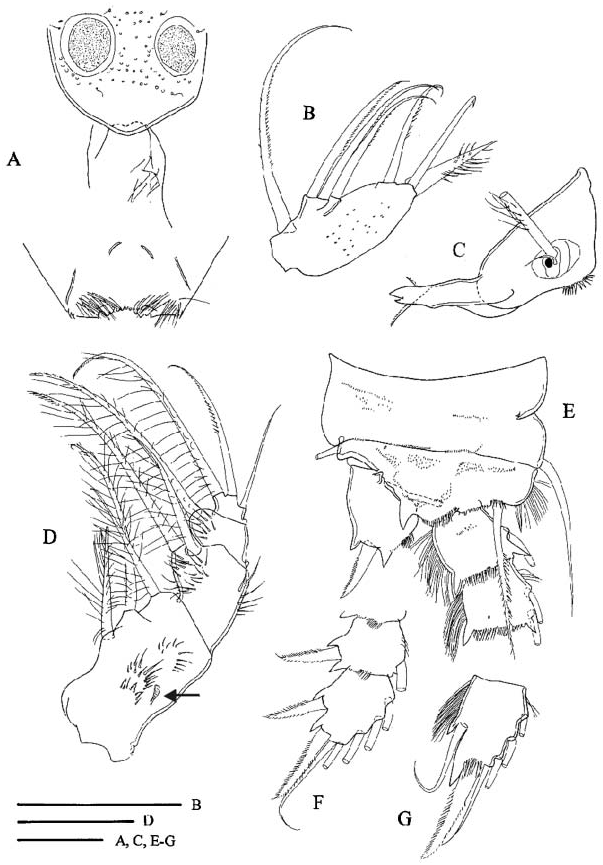
Labrum ( Figure 2A View Figure 2 ) has small medial teeth on distal margin, lateral protuberances with two or three small spinules. In one female (Emirgân, ♀ -2) long hairs observed on left side of epistoma, but epistoma bare in another specimen (Foglino, MIZ: 157867), where this feature could be verified. Palp of mandible ( Figure 2C View Figure 2 ) with two long and one short setae, large spinules present in longitudinal row near palp on frontal surface of coxopodite. Maxillula with seta armature typical of family, palp ( Figure 2B View Figure 2 ) ornamented with small spinules, palp setae without long setules. Maxilla composed of syncoxopodite with three endites (two, one and two setae, respectively), basipodite (two setae), and two-segmented endopodite (two and three setae). Maxilliped ( Figure 2D View Figure 2 ) with syncoxopodite (three setae), basipodite (two setae) and two-segmented endopodite (one and three setae). Groups of large spinules present at levels between insertion of distalmost and median setae, and insertion of proximalmost seta on frontal surface of syncoxopodite. In single female (Emirgân, ♀ - 1), one of the maxillipeds bearing third group of spinules proximally to groups mentioned above. Membranous element ( Figure 2D View Figure 2 , arrowed) located lateral to groups of spinules.
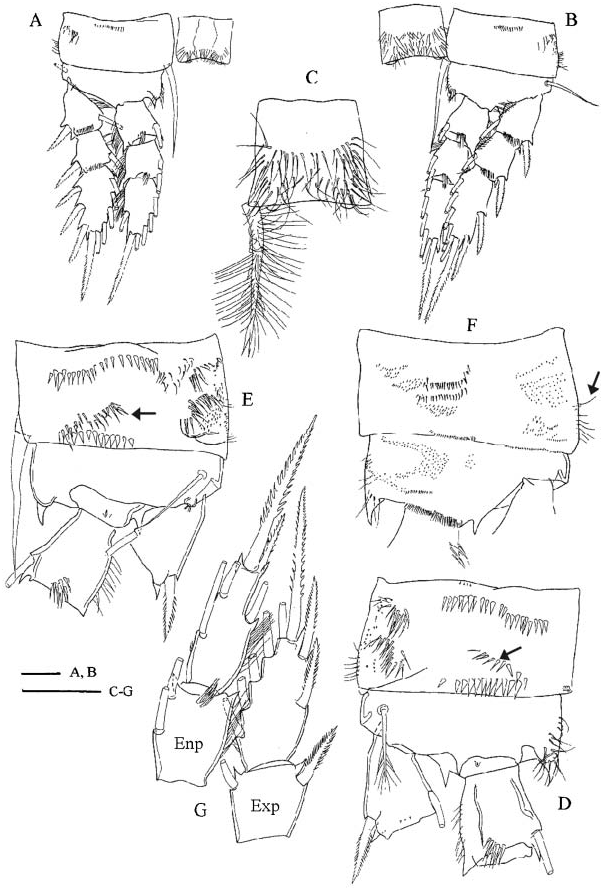
Legs ( Figures 2E–G View Figure 2 ; 3A–G View Figure 3 ). Armature formula as in Table 2. Terminal exopodal segments of P1–P4 bearing two, four, three and three spines, respectively. Medial spine of P1 basipodite ( Figure 2E View Figure 2 ) bearing only small spinules, no long setules in proximal half. Spine reaching beyond distal margin of second endopodal segment. Large spinules arranged in arc between implantation of endo- and exopodites, characteristic of genus (cf. C. abyssorum divergens in Figure 9B View Figure 9 ), absent on frontal surface of P1 basipodite ( Figure 2E View Figure 2 ). Medial expansion of basipodite of P1–P4 pilose ( Figures 2E View Figure 2 , 3A,B,D,F View Figure 3 ), in single specimen (Emirgân, ♀ -1) hairs appear asymmetrically on left basipodite of P4 and absent on right side ( Figure 3E View Figure 3 ). P1 coupler bare in six of seven specimens examined, and caudally pilose in one female (Emirgân, ♀ -1). P2 coupler caudally pilose in specimens from Turkey ( Figure 3A View Figure 3 ) and bare in females from Italy. Couplers of P3 and P4 pilose in all specimens ( Figure 3B,C View Figure 3 ). Obtuse distal protuberances of P4 coupler ( Figure 3C View Figure 3 ) emerging only slightly beyond distal margin. P4 coxopodite seta ( Figure 3C View Figure 3 ) reaching well beyond medial expansion of basipodite. Caudal surface of P4 coxopodite ( Figure 3D,E View Figure 3 ) ornamented with 8–15 robust spinules of similar size along distal margin, group of long spinules at laterodistal angle and small spinules lateral to this group, long row of robust spinules nearer to proximal margin, long spinules at lateroproximal angle, and hairlike spinules on the lateral margin. In both females from Turkey an additional oblique row of thin spinules ( Figure 3D,E View Figure 3 , arrowed) added to the pattern described above. Frontal surface of coxopodite and basipodite of P4 (verified in specimens from Turkey) also adorned with small spinules ( Figure 3F View Figure 3 ). Lateral hairs on first endopodal segment shorter than those on second and third endopodites ( Figure 3D,G View Figure 3 ). P5 ( Figure 1E View Figure 1 ) setation typical of genus. Medial spine inserted in distal half (in five of six females verified) or in middle of segment. Small spinules present at implantation of apical and lateral setae and medial spine.
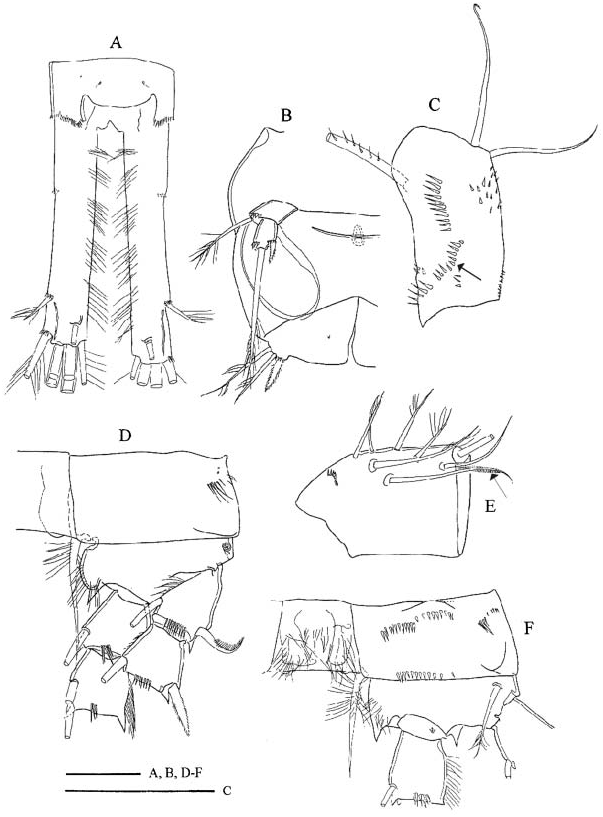
Male ( Turkey and Italy). Body length 1100–1300 Mm. Pediger 5 not pointed outwards ( Figure 4B View Figure 4 , cf. female in Figure 1E View Figure 1 ). Large pore present on ventral anterior part of genital somite. Except for medial cuticular pore, no ornamentation of P6 flap ( Figure 4B View Figure 4 ). Caudal ramus ( Figure 4A View Figure 4 ) 6.6–8.3 times as long as wide, no longitudinal keel on dorsal surface. Lateral caudal seta 0.18–0.21 times as long as ramus. Dorsal seta 1.1–1.2 times longer than lateralmost terminal seta. Relative length of terminal caudal setae from medialmost to lateralmost: 1.3–1.5, 5.0–6.8, 3.8– 4.9, 1.0 (seta length/ramus length: 0.56–0.73, 2.3–3.2, 1.6–2.2, 0.45–0.53). Longest terminal caudal seta about 0.8–1.1 times as long as urosome, tip slightly curved.
Antennule 16-segmented, with compound (segments 16 and 17 anteriorly fused) terminal segment, armature formula: 8+1 aesthetascs ( Figure 4E View Figure 4 , aesthestasc arrowed), 4, 2, 2+aesthetasc, 2, 2, 2, 2, 1+spine+aesthetasc, 2, 2, 2, 2+aesthetasc, 2, 1+aesthetasc, (4+aesthetasc, 7+aesthetasc). Plate-like structures (one large on segment 14, and two smaller on segment 15) and short conical elements (one each on segments 14 and 15) at distal geniculation. Second endopodal segment of antenna with eight setae. Triple ( Turkey) ( Figure 4C View Figure 4 , arrowed) or double ( Italy) row present proximally to longitudinal row on caudal surface of antennal coxobasis. Syncoxopodite of maxilliped frontally adorned with one or two groups of spinules.
In one ( Turkey) of the 12 males verified, spine formula was 2333, in all the other specimens the spine formula was as in female, 2433. Medial spine of P1 basipodite reaching only little beyond distal margin of enp2. Spine on P2 exp1 ( Figure 4D View Figure 4 ) more slender and curved, spinules on spine longer than those on other spines of swimming legs. P1–P2 couplers without ornamentation, P3 bare or caudally pilose, P4 couplers with hairs on caudal surface. Medial expansion of basipodite of P1–P4 pilose. Caudal spinule ornamentation of P4 coxopodite ( Figure 4F View Figure 4 ) reduced in comparison to that of female: oblique row next to row along distal margin, absent or present (Emirgân, „ -2); lateral section, except for lateroproximal group of spinules, devoid of ornamentation. P4 enp 3 2.3–2.8 times as long as wide, medial apical spine 1.1–1.3 times longer than segment, and 2.2–2.8 times as long as lateral spine. Medial spine, and apical and lateral setae of P5 0.5–0.8, 4.2–4.8 and 2.4–3.4 times as long as segment 2. Proportional lengths of P6 setae from medialmost to lateralmost in males from Turkey and Italy 1.0, 1.4–1.7, 2.2 and 1.0, 2.1, 3.4–4.1, respectively.
Copepodid V, female. Body length 1330 Mm; prosome/urosome 1.6. Caudal ramus 5.0 times as long as wide, no keel on dorsal surface. Lateral caudal seta inserted at distal third, 0.19 times as long as ramus. Dorsal seta 0.8 times as long as lateralmost terminal seta. Relative length of terminal caudal setae from medialmost to lateralmost: 1.4, 6.6, 4.9, 1.0 (seta length/ramus length: 0.66, 3.1, 2.3, 0.47). Longest (inner median) terminal caudal seta 1.1 times as long as urosome.
Antennule 11-segmented, reaching slightly beyond cephalothorax. Armature formula: 7, 4, 8, 4, 1+sp, 2, 3, 2+ae, 2, 2+ae, 7+ae. Aesthetascs homologous with those on segments 12 and 16 in the adult female long, 40 Mm and 34 Mm, respectively. Antenna bearing eight setae on second endopodal segment. Mandible with longitudinal row of large spinules near palp. Segmentation and armature formula of leg 1–4 as in adult female, except for presence of three spines on third exopodal segment of right ramus of P1. Two spines on exp3, state typical of species, present on left ramus of P1. Medial spine on P1 basipodite spinulose, large spinules absent on frontal surface of segment, in between implantation of exo- and endopodites. Couplers caudally pilose on P3–P4, and bare on P1–P2. Spinule ornamentation on caudal surface of P4 coxopodite more similar to that of male and consisting of: 10 robust spinules of equal size along distal margin; long row of robust spinules nearer proximal margin; and long spinules at lateroproximal margin. Other elements absent. Medial expansion of basipodites of P1–P4 apically pilose. Of two terminal spines of P4 endopodite, medial spine 2.1 times as long as lateral.
Medial spine of P5 inserted at 0.6 length of second segment, and 0.81 times as long as segment 2. Apical and lateral setae 3.8 and 2.4 times as long as segment 2. Small spinules present at implantation of apical and lateral setae and medial spine.
Comments
Cyclops ankyrae Mann, 1940 was originally described from a small pool in Central Anatolia (Ankara). Depository of the type (s) is unknown. Kiefer (1955) reported on the occurrence of Cyclops furcifer from the Bosporus region as well as the European borderlands of the Sea of Marmara in Turkey (Emirgân, pond; Kâğithane, puddle; Florya, pool), and found only slight differences between these Turkish specimens and the original description of C. ankyrae (caudal ramus length/width: 7.5–9.0 vs. 7.0 in C. ankyrae ; second segment of P5 longer than wide vs. as long as wide in C. ankyrae ; shape of pedigers 4 and 5 somewhat different in the two forms). From among the 12 females that Kiefer examined, 11 had a spine formula of 2433, characteristic of C. ankyrae , yet in one female from Emirgân the spine armature on the terminal exopodal segments of P1–P4 differed on two sides of the body (2433 and 3433). As intraspecific variation of the spine formula had already been observed in the Eastern European populations of C. furcifer (dominantly 2333, sometimes 3433, or 2433) ( Koźmiński 1933), Kiefer concluded that specimens from Emirgân, Kâğithane and Florya were conspecific with the former taxon, and he considered the Central Anatolian C. ankyrae to be no more than a subspecies of C. furcifer ( Kiefer 1955) . Based on the material originally used by Kiefer (SMNK Glas: 1119, 1391), Einsle (1992) redescribed and illustrated the Cyclops specimens from Emirgân and Kâğithane and identified them as C. ankyrae . Species status of the taxon was reconfirmed in the monograph of the genus ( Einsle 1996a). The Turkish specimens examined here come from the same material that Kiefer (1955) and Einsle (1992, 1996a) investigated.
Einsle (1996a) mentioned two diagnostic characters, which distinguish C. ankyrae from C. furcifer : presence of lateroventral lobes on pediger 5 in C. furcifer (lobes absent in C. ankyrae ); and the spine formula, which is usually 2333 in C. furcifer but 2433 in C. ankyrae . My investigations also revealed other characters that are useful in species identification. The anal operculum is distinct in C. ankyrae ( Figure 1J View Figure 1 ), but weak in C. furcifer ( Figure 1I View Figure 1 ). Hairlike spinules on the lateral margin of the P4 coxopodite present in adult female C. ankyrae ( Figure 3F View Figure 3 , arrowed), are absent in female C. furcifer . In C. ankyrae the mandibular coxopodite frontally bears a longitudinal row of large spinules near the palp ( Figure 2C View Figure 2 ); there is no ornamentation on the mandibular coxopodite in C. furcifer . In C. furcifer the frontal surface of maxilliped syncoxopodite is adorned with one distal group of spinules, while in C. ankyrae ( Figure 2D View Figure 2 ) one or sometimes two additional groups beyond the spinules also present in C. furcifer appear more proximally at the level of insertion of the proximalmost seta. It is worth noting that the above-mentioned character states, with the exception of ornamentation characters on the P4 coxopodite and lack of lateroventral lobes on pediger 5, are useful diagnostic characters in the male specimens too. The last two characters show sexual dimorphism in the genus Cyclops : the spinule ornamentation on the caudal surface of the P4 coxopodite is usually reduced in comparison to that of the female, and the lateroventral lobes that are present on pediger 5 of the female are absent in the male (cf. description C. abyssorum divergens ). The similarity of the morphometric traits and other character states (spine formula of 2433, and strong anal operculum) that are rare within the genus indicate that Cyclops from the Bosporus region and Central Italy is conspecific with C. ankyrae rather than with C. furcifer . The female specimens from Turkey and Italy differ in the relative length of the median terminal (longest) caudal seta [shorter in Italian females ( Table 1)] and some ornamentation characters of the legs [P2 coupler bare in Italian vs. pilose in Turkish specimens (cf. Figure 3A View Figure 3 ); and the oblique row of spinules on the caudal surface of P4 coxopodite present in two Turkish females (cf. Figure 3D,E View Figure 3 ), is absent in the Italian females]. To understand the taxonomic significance of these differences we need more data on the geographic variation of these characters. Intraspecific (intrapopulation) variation of the pilosity of the couplers of P1–P4 has already been observed in C. ladakanus ( Mirabdullayev 2008) , C. furcifer and C. glacialis (unpublished data of the author), which may suggest that the morphological difference mentioned does not indicate a species level separation of the Italian and Turkish populations.
Among Cyclops , only two other species, C. ladakanus Kiefer, 1936 [Indian and Chinese Tibet: various waterbodies at 4200–5200 m above sea level (a.s.l.) ( Kiefer 1936; Tai and Chen 1979)] and C. pamirensis Gurvich, 1958 [Pamir: Kara-Kul – salt lake ca. 4000 m a.s.l. ( Gurvich 1958)] bear two, four, three, three spines on the terminal exopodal segments of P1–P4. Yet, unlike C. ankyrae , the pediger 5 is not pointed outwards in C. ladakanus and C. pamirensis . Einsle (1992, 1996a) considered C. pamirensis to be a junior synonym of C. ladakanus and redescribed this latter taxon from Tibet. The somewhat modified drawings in his later publication ( Einsle 1996a) show additional diagnostic characters of C. ladakanus , like the hairy ornamentation of the proctodeum, which can distinguish this species from C. ankyrae . Mirabdullayev (2008) redescribed the Cyclops from Pamir (Lake Shorkul and Karakul), based on material deposited in the Gurvich Collection, and confirmed the synonymy of C. pamirensis with C. ladakanus . Ornamentation of the proctodeum in the Pamir specimens is not mentioned by Mirabdullayev (2008).
The morphology of many representatives of Cyclops is still too poorly understood to propose a sound hypothesis for the systematic position of C. ankyrae within the genus. The microcharacters of the mouthparts, namely the spinule ornamentation of the mandibular coxopodite and maxilliped syncoxopodite, are shared with C. vicinus Ulianin, 1875 and C. kikuchii Smirnov, 1932 (the spine formula is 2333 in both species). On the other hand, the appearance of hairs also on those couplers that are inserted anteriorly to P4, might indicate close relationships with C. furcifer [hairs present on couplers of P2–P4 or P3–P4 (variation observed in females from UK); spine formula variable as mentioned above] and C. ladakanus [couplers pilose on P1–P4 or P2–P4 ( Einsle 1996a; Mirabdullayev 2008), spine formula 2433]. The couplers, except for that of P4, are bare in C. vicinus and C. kikuchii . A hairy ornamentation also appears on the couplers of P2–P4 or P3–P 4 in C. glacialis Flössner, 2001 . This latter species was described from a mountain lake (freshwater, 2650 m a.s.l.) in northwestern Mongolia ( Flössner 2001; Flössner et al. 2005), and shows affinities with C. abyssorum Sars, 1863 (spine formula is 3433 in both taxa). The fact that extended pilosity of the couplers occurs in a morphologically heterogeneous species assemblage may hint at its plesiomorphic state in Cyclops . Cyclops ankyrae , C. vicinus , C. kikuchii , C. furcifer and C. insignis Claus, 1857 share the lack of large spinules arranged in an arc on the frontal surface of the P1 basipodite, a character state that is apparently rare in the genus. More data on the morphology of some poorly known Asian taxa described from Iran ( C. caspicus Lindberg, 1942 , C. kozminskii Lindberg, 1942 ) and Kashmir in India ( C. hutchinsoni Kiefer, 1936 ) may shed new light on the relationships of C. ankyrae .
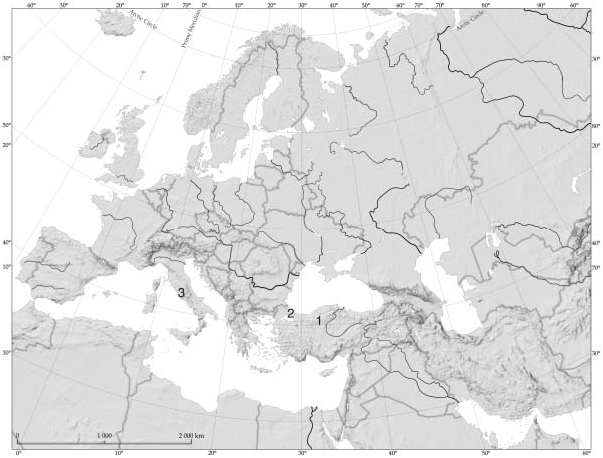
Little is known of the geographic distribution of C. ankyrae ( Figure 5 View Figure 5 ). Beyond the occurrence in Central Anatolia and European borderlands of the Sea of Marmara in Turkey, the species has also been reported from Iran ( Einsle 1996a), Macedonia ( Petkovski 1983), and northeastern and Central Italy ( Stoch 2006).

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Cyclops ankyrae Mann, 1940
| Hołyńska, Maria 2008 |
Cyclops furcifer
| Kiefer F 1955: 108 |
Cyclops ankyrae
| Einsle U 1996: 14 |
| Einsle U 1992: 183 |
| Mann AK 1940: 65 |