
Liolaemus sitesi, Avila, Luciano Javier, Olave, Melisa & Perez, Cristian Hernan Fulvio, 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3608.4.1 |
|
publication LSID |
lsid:zoobank.org:pub:49065DDB-7EA1-443F-8A07-3462D0B4394C |
|
DOI |
https://doi.org/10.5281/zenodo.6150274 |
|
persistent identifier |
https://treatment.plazi.org/id/B96E87EA-FFD5-4C2E-FF46-252612EEF9A8 |
|
treatment provided by |
Plazi |
|
scientific name |
Liolaemus sitesi |
| status |
sp. nov. |
Liolaemus sitesi sp. nov.
( Figure 2 View FIGURE 2 )
Holotype. — MLP.S 2637, an adult male from Park Ranger Post, 25.9 km S junction Provincial Road 6, road to Auca Mahuida volcano, Auca Mahuida Natural Protected Area (37º 42’ S, 68º 51’ W, 1560 m), Pehuenches Department, Neuquén Province, Argentina. L.J. Avila, M.L. Kozykariski, and M.F. Breitman, collectors.
Paratypes. — LJAMM-CNP 11021, 12300, 12243, 12328-30, males, LJAMM-CNP 12301, 12331-4, females, LJAMM-CNP 12335-7, 12212-3, juveniles, same data as the holotype. LJAMM-CNP 12305-6, males, Cerro del Este (37º 46’ S, 68º 53’ W, 1935 m), L.J. Avila, M.L. Kozykariski, and M.F. Breitman collectors. LJAMM-CNP 10566, juvenile, eastern slope of Auca Mahuida volcano (37º 41’ S, 68º 49’ W, 1332 m), C.H.F. Pérez and D.R. Pérez, collectors. LJAMM-CNP 10558, male, northeastern slope of Auca Mahuida volcano (37º 43’ S, 68º 53’ W, 1757 m), C.H.F. Pérez and D.R. Pérez, collectors. All of these localities are located in Auca Mahuida Natural Protected Area, Pehuenches Department, Neuquén Province, Argentina. LJAMM-CNP 13382, female, northern slope of Auca Mahuida volcano (37º 47’ S, 68º 53’ W, 1486 m), LJAMM-CNP 14234, female, 14236, male, track to Punta de Castillo (37º 49’ S, 68º 54’ W, 1366 m), L.J. Avila and I. Minoli, collectors. LJAMM-CNP 14241, female, 0.5 km W La Aguada (37º 45’ S, 68º 54’ W, 1843 m), L.J. Avila, I. Minoli, M. Kozykariski, D. Janish Alvarez and S. Quiroga, collectors. All of these localities are also located in Auca Mahuida Natural Protected Area, Añelo and Pehuenches Departments, Neuquén Province, Argentina.
Diagnosis. — Liolaemus sitesi is a member of the rothi complex, that includes L. hermannunezi , L. loboi , L. sagei , L. rothi , and L. tromen and several other potential species that require further study (Avila et al. 2006; Morando et al. 2004; Olave unpublished data). Liolaemus sitesi can be easily distinguished from all other members of its clade by its black/dark brown background body coloration with small bright yellow, green or white spots distributed along its dorsal and lateral trunk areas; in full sun males take on a yellow-green iridescent coloration on neck, body, limbs and tail; colors not observed in any other species of the group. Liolaemus sitesi differs from L. rothi , L. sagei and L. tromen by its marked sexual dichromatism. Liolaemus sitesi is smaller than L. rothi (maximum SVL: 79.8 vs. 97.00 mm), has more scales around midbody (72.0 ± 2.9 vs. 64.7 ± 4.27) and more supralabial scales (8.0 ± 0.9 vs. 5.49 ± 0.56). Liolaemus sitesi lacks of the conspicuous yellow spots irregularly distributed along the body of L. rothi . Liolaemus sitesi is larger than L. hermannunezi (maximum SVL: 79.8 vs. 71.4 mm), has more dorsal (82.0 ± 2.5 vs. 71.8 ± 1.1), ventral (101.4 ± 4.2 vs. 87.3 ± 1.0), and supralabial scales (8.0 ± 0.9 vs. 7.1 ± 0.7). Liolaemus sitesi is smaller than L. sagei (maximum SVL: 79.8 vs. 88.00 mm), has fewer scales around midbody (72.0 ± 2.9 vs. 90.3 ± 4.68), fewer dorsal scales (82.0 ± 2.5 vs. 101.2 ± 6.59), more supralabials (8.0 ± 0.9 vs. 5.25 ± 0.60), and fewer infralabial scales (5.6 ± 0.4 vs. 6.22 ± 0.42). Liolaemus sitesi is larger than L. loboi (maximum SVL: 79.8 vs. 72.7 mm), has more scales around midbody (72.0 ± 2.9 vs. 63.7), more dorsal scales (82.0 ± 2.5 vs. 67.1), and fewer infralabials (5.6 ± 0.4 vs. 6.4). Liolaemus sitesi differs from L. tromen by having a slender body, a higher number of scales around midbody (72.0 ± 2.9 vs. 65.8 ± 2.2), more dorsal (82.0 ± 2.5 vs. 72.2 ± 3.1) and supralabial scales (8.0 ± 0.9 vs. 7.0 ± 0.6), and a coloration pattern without suprascapular marks or series of subsquare paravertebral spots fused forming transversal bands.
Description of holotype.—Adult male ( Figs. 2 View FIGURE 2 , 3 View FIGURE 3 ), 76.1 mm snout-vent length (SVL); tail length 114.2 mm. Axilla-groin distance 34.9 mm. Head length 16.3 mm (from anterior border of tympanum to tip of snout), 16.3 mm wide (at anterior border of tympanum), 9.1 mm high (at anterior border of tympanum). Snout length 6.4 mm (orbittip of snout distance). Interorbital distance 6.4 mm. Eye-nostril distance 5.1 mm. Orbit-auditory meatus distance 6.4 mm. Forelimb length 22.3 mm. Tibial length 14.9 mm. Foot length 23.6 mm (ankle to tip of claw on fourth toe). Dorsal head scales bulged, smooth, 19 between occiput at level of anterior border of tympanum to rostral, pitted with numerous scale organs in the anterior region, reduced to a single organ, or without organ in the posterior half of the head. Rostral scale wider (2.8 mm) than high (1.4 mm). Two postrostrals, together with anterior lorilabial, separate nasal scales from rostral. Nasal scales longer than wide, rhombic; nostril one-half length of nasal, posterior in position. Scales surrounding nasals 8–6 (on the left and right side respectively). Five internasals. Three frontonasal scales, two on the sides approximately squared, and one central rectangular and twice as large. Five prefrontals, a small pentagonal scale in the center (1.3 mm), and two pairs on the sides: a pair of larger dorso-lateral scales, irregularly hexagonal (1.9 mm), and a pair of medium-sized lateral scales (1.7 mm), almost pentagonal. Six frontal scales, fragmented. Six frontoparietal scales in two longitudinal rows. Interparietal hexagonal (1.9 mm), surrounded by seven scales; five smaller and irregular in front and sides (0.88 mm), and two larger posteriorly (1.6 mm). Parietal eye evident. Parietals irregularly shaped, some bulged. Circumorbitals 9–13 (damaged on the left side). Transversally expanded supraoculars 7–8; lateral supraoculars smaller: 19–18. Anterior canthal approximately square, separated from nasal by one postnasal. Posterior canthal longer than wide, subtriangular. Loreal scales, something bulged, five on each side. Lorilabials longer than wide (8–7), larger than labials. Seven superciliaries on each side, flattened and elongated, anterior four broadly overlapping dorsally. Orbit with 16–14 upper and 14–13 lower ciliaries. Orbit diameter 6.1 x 2.4 mm. Preocular small, unfragmented, longer than wide. Subocular scale elongated, five times longer than wide (4.3 x 0.8 mm). A well marked longitudinal ridge along upper margin of preocular and subocular scales. Postocular small, slightly bulged, very inconspicuous. Palpebral scales small granular and bulged. Supralabials 9–7, quadrangular, convex. Temporals smooth, convex, imbricate with one scale organ in the tip. Anterior auriculars smaller than adjacent posterior temporals, granular. Posterior auriculars small and granular. Smaller than the anterior auriculars. External auditory meatus conspicuous, higher (3.0 mm) than wide (1.1 mm). Lateral scales of neck granular with inflated skin. Mental scale wider (3.0 mm) than high (1.4 mm), in contact with four scales. Mental followed posteriorly by two postmentals, and two rows of three chinshields on each side. Six infralabials on each side, first scale twice as high as posterior infralabials, posterior slightly larger than supralabials. Gular scales smooth, flat, imbricated, with rounded posterior margins, with melanophores. Scales of throat between chinshields slightly juxtaposed, becoming slightly imbricated toward auditory meatus. Thirty-two gulars between tympanum openings. Infralabials separated from chinshields by one to three rows of scales. Antehumeral and longitudinal neck folds well developed; postauricular very distinctive; gular, rictal, dorsolateral, oblique inconspicuous.
Scales of dorsal neck region rhomboidal, imbricate, slightly keeled. Seventy-eight dorsal scales between occiput and anterior surface of thighs. Dorsal body scales rhomboidal, imbricated, slightly keeled to moderately keeled towards the posterior half of body. Dorsal scales grading laterally into slightly smaller, smooth scales at midbody. Scales, immediately anterior and posterior to forelimb and hindlimb insertion, small, smooth, granular, and non-overlapping. Body lateral scales grading from slightly smaller to larger towards the ventral region, at midbody. Ventral body scales rhomboidal, smooth, flat, imbricate, and larger than dorsal scales. Seventy-six around midbody; scales between mental and precloacal pores 99. Scales of cloacal region about equal in size to ventral body scales. Nine conspicuous precloacal pores.
Anterior suprabrachials rhomboidal, imbricated, smooth, slightly larger in size to dorsal body scales. Infrabrachials smaller, granular. Postbrachials smaller, smooth, becoming granular near axilla. Supra-antebrachials similar to suprabrachial. Infra-antebrachials rhomboidal, imbricate, smooth. Supracarpals imbricated, rhomboidal, smooth. Infracarpals strongly imbricated, rhomboidal, smooth, with few scales moderately keeled towards the anterior area. Subdigital lamellae with 3 keels, numbering: I: 11, II: 15, III: 21, IV: 23, V: 14. Claws robust, curved and sharp, opaque light brown.
Suprafemorals as large as dorsal body scales, rhomboidal, imbricated, smooth, some very slightly keeled. Infrafemorals scale small, granular and smooth. Femoral patch with enlarged and bulged scales, with forty-one scales, ending in a blunt margin. Supratibial rhomboidal, imbricated, slightly keeled. Infratibials rhomboidal, imbricated, smooth. Supratarsals rhomboidal, imbricated and smooth. Infratarsal small, rhomboidal, imbricated, smooth. Subdigital scales 1–2 keeled, numbering: I: 10, II: 17, III: 24, IV: 28, V: 18. Claws robust, curved and sharp, opaque light brown. Tail complete, non-regenerated. Dorsal caudal scales quadrangular and moderately keeled. Dorsal-lateral caudal scales quadrangular and slightly keeled. Ventral subtriangular and smooth, toward posterior half slightly keeled.
Color in life. — Dorsal background coloration black/dark brown with nine black transversal series of four blotches with posterior notches occupied by white spots from the occiput to the rump. First three series fused and irregular; fourth and fifth shaped as half-moon, almost in contact; all others becoming well-separated, rounded and uneven to rump. Paravertebral blotches with 3–5 white dots on its posterior margins, each dot composed by 1–2 scales. Five dorsolateral black blotches from the neck to the groin, rounded, with 2–5 white dots on its margins, each dot composed of 1–2 scales. Six dorsolateral, transversals and elongated black blotches from the axilla to the groin, with 4–11 white dots on its margins, each dot composed of 1–2 scales. Lateral neck region, from the postauricular area to the shoulder, black with scattered white spots of 1–8 scales (small and granular). Axilla dark brown with white dots formed by 1–6 scales (small and granular). Body lateral region dark brown with some scattered white dots of one scale. Head dark brown with maxillary region and half temporal area melanic with a white dot on each scale. Limbs light brown with inconspicuous dark reticulations. Tail beige with three inconspicuous black blotch series to level of the postcloacal region, to join in only one blotch becoming less marked toward the tip. Throat and gular region fully melanic. Chest, belly, ventral region of the limbs and tail melanic. In preservative the general color pattern is conserved but the dorsal background and ventral coloration become darker. All yellow or green colors disappear. On the dorsal surface of the head darker blotches become more evident, one longitudinal blotch on the prefrontals and postnasals scales, two subtriangular blotches on anterior region in the orbits, two small blotches at the back of the orbits, and several smaller irregularly arranged blotches in the temporal, parietal and occipital regions.
Variation. — Liolaemus sitesi shows a remarkable sexual dichromatism ( Figs. 2 View FIGURE 2 , 4 View FIGURE 4 & 5 View FIGURE 5 ). In eleven males ( Table 3 View TABLE 3 ): SVL: 60.8–79.8 mm. Axilla–groin distance: 27.5–38.7 mm. Head length: 13.6–16.9 mm. Head width: 10.9–13.6 mm. Head height: 7.5–9.6 mm. Foot length: 20.2–23.6 mm. Tibial length: 12.4–15.8 mm. Arm length: 19.2–22.5 mm. Midbody scales: 67–78. Dorsal scales (between occiput at the anterior margin of auditory meatus and anterior surface of thighs): 78–86. Ventral scales: 95–104. Third finger lamellae: 18–23. Fourth toe lamellae: 25–29. Supralabial scales: 7–10. Infralabial scales: 5–6. Cloacal pores: 7–10. In eleven females ( Table 3 View TABLE 3 ): SVL: 56.4–73.9 mm. Axilla–groin distance: 25.7–38.7 mm. Head length: 12.2–14.6 mm. Head width: 9.9–11.9 mm. Head height: 6.7–8.2 mm. Foot length: 18.3–22.0 mm. Tibial length: 11.6–13.8 mm. Arm length: 18.0– 20.9 mm. Midbody scales: 67–76. Dorsal scales: 80–87. Ventral scales: 98–112. Third finger lamellae: 19–22. Fourth toe lamellae: 25–30. Supralabial scales: 7–9. Infralabial scales: 5–6. Liolaemus sitesi exhibits strong sexual dichromatism. The female LJAMM-CNP 13382 ( Fig. 3 View FIGURE 3 ) exhibits grayish-brown dorsal background coloration with a series of eleven vertebral dark brown blotches. The three first series formed by three round blotches with a white dot on the back, covering 1–2 scales; the following six series shaped an hourglass with a white dot on the posterior concavity, composed of 1–3 scales; and the last two series, round, with one white dot of one scale. Nine dorsolateral series of dark brown blotches. The first three series round with a white dot on the posterior margin, composed of five scales; the following four series elongated and transversal with a white dot on the posterior concavity, composed of 2–3 scales; the last two that are above the groin, round with a white dot composed of four scales. The lateral region of the body with a yellowish color and five brown lateral blotches, elongated, inconspicuous, with a white dot composed of 2–3 scales. The anterior area of tail, at the level of the postcloacal area, presents three dark brown blotches series, rounded, with a lighter band in the posterior margin. This series then becomes one central blotch from postcloacal area to the tip of the tail. These blotches are brown with a dark brown posterior margin. Limbs lighter brown than the dorsal area with a dark reticulate. Head gray with a blotch in the anterior area of the head fork shaped, and small irregular blotches in posterior area. Lateral head gray with postciliar brown and elongated blotch. Gular region, chest, belly and ventral region of the limbs and tail, gray. In all females dorsal background color is the same, only varying in intensity, just like the series of dorsal blotches. The white dots in the posterior area of the brown blotches vary only in size and scale number. Only one female has white scattered dots as males but in lower density. A single specimen has white blotches of 2–12 scales between vertebral and paravertebral series. All specimens have the brown blotches in the head, but one specimen shows some small white blotches between the brown blotches. Four specimens have two dorsolateral dashed white lines from the occiput to the postcloacal area. In three juveniles, the dorsolateral lines are inconspicuous, but these are more evident in one adult female with a yellowish color from the middle of the body toward the rear. Three specimens also exhibit lateral white lines between axilla and groin but usually this characteristic is not found in mature females of larger size. One specimen shows light gray coloration on the throat and gular region, another black reticulation, and the remaining females an inconspicuous dark reticulate. Ventral coloration of the chest, belly, limbs and tail vary between white and dark gray.
Basking adult males exhibit a brightly yellow or yellow/green emerald iridescent general coloration that disappears almost immediately after capture, this coloration is only evident after a few minutes of basking but not in individuals just emerged from bush or rock shadows, or burrows. Dorsal background of the males only varies in intensity, but in four specimens, blotch series vary between black and brown, slightly darker than the background. The white dots vary in size and a single specimen varied in color. On three males, dots are formed by 1-8 scales. In only two males did the back margin of the brown blotches give the appearance of bands. In one male of the type series, but in several males observed alive but not collected, dots were bright yellow (see figure 2), this coloration was lost in preservative. On the dorsal surface of head, some observed individuals have darker blotches, one longitudinal blotch on the prefrontals and postnasals scales, two subtriangular or quadrangular blotches in the anterior region in the orbits, two small at the back of the orbits and several smaller irregularly arranged upon the temporal, parietal and occipital region. These blotches became evident on the holotype after some time on preservative. In the tail, both the background and the blotches vary in some specimens only in intensity ( Fig. 5 View FIGURE 5 ).
All adult males have ventral melanism with variable degree of development, but in all individuals the gular and throat regions are always black. In three individuals the melanism in the mental region or the tail disappears. In immature males melanism is incipient with less extension and intensity, but one individual has a dark reticulate pattern on the throat and gular region. Juveniles have a color pattern similar to females but with the dorsolateral and lateral light lines always present. The throat and gular region white with a dark gray reticulate, the chest and belly white, and tail gray ( Fig. 5 View FIGURE 5 ).
Etymology.—The specific name is to honor our friend and colleague Jack Walter Sites, Jr. from Brigham Young University, Provo, Utah, USA, who strongly supported and collaborated with us on studies of Patagonian herpetofauna for the last 12 years. Dr. Sites is a herpetologist and evolutionary biologist and devoted the last 30+ years to collaborate with herpetologists from many Central and South American countries, including Argentina, Bolivia, Brazil, Colombia, Chile, Mexico, Peru, Uruguay and Venezuela.
Geographic distribution.— Liolaemus sitesi is known only from Patagonian Steppe vegetation environments found above 1300–1500 m elevation in Auca Mahuida Volcanic Field, Añelo and Pehuenches Departments, in Neuquén Province ( Fig. 6 View FIGURE 6 ). The main physiographic element in the Auca Mahuida Volcanic Field is the volcano of the same name that occupies the majority of the volcanic field and harbors all the Patagonian Steppe environments. This environment is isolated from other similar Patagonian Steppe environments, and surrounded by dune fields or sandy/rocky flatlands with extremely arid characteristics and vegetation typical of the Monte phytogeographic region. Liolaemus sitesi is not in contact with the geographic distribution range of other members of the rothi group; the geographically closest related species is Liolaemus tromen found in the Tromen volcanic field, near 130 km NW of Auca Mahuida volcanic field.
Natural history.— Lizards were found by active search along transects established along the main roads, trails or open field on the slopes of the Auca Mahuida volcano. They were most frequently found basking on rocks and when approached would drop off the rock directly into a burrow, or flee to hide under another rock or burrow under roots. They were often found by turning over rocks. They have never been seen to climb the low vegetation or bury in the sand, but early in the morning juveniles were commonly observed in the sand accumulations found in the edges of shrubby vegetation ( Prosopis denudans, Senna kurtzii, S. arnottiana). Adults were wary and dropped off into burrows below the stones when the intruder is still at a considerable distance. If pursued, they would run from rock to rock across patches of bare soil, usually seeking cover in vegetation.
Individuals were observed active between 09:00 to 19:00 on sunny days of austral summer, but showed a bimodal activity pattern on the hottest days and a unimodal pattern on cloudy or cold days. Several individuals were observed sharing basking areas, and usually juveniles and adults were found under small bushes, sharing refuge or foraging areas. Juvenile activity was more frequent in early hours of the morning or late hours of the afternoon. They seem to be territorial at some point because aggressive encounters between males or females were observed. This species shares its habitat with other Liolaemus species ( Liolaemus cyaneinotatus , Liolaemus austromendocinus , Liolaemus aff. elongatus ), as well as Diplolaemus leopardinus , Leiosaurus belli , Phymaturus sitesi and P. roigorum . Liolaemus sitesi usually occupies the microhabitat around scattered shrubs separated by an open, loose sand substrate ( Fig. 7 View FIGURE 7 ). No data about reproduction are available, but L. sitesi is probably oviparous like others members of its clade (Cei 1986). Our analysis of two guts revealed a few ant heads and beetle legs, mixed with partially digested small leaves and vegetable remains. Several snakes were observed in the area, including Bothrops ammodytoides , Philodryas trilineata , Pseudotomodon trigonatus and Micrurus pyrrhocryptus , all potential predators of this species.
Remarks.—In 1898, Julio Koslowsky described Liolaemus rothi from the ‘‘territorio del Neuquén’’. Since then, numerous similar populations from central and northwestern Patagonia have been referred to this species. Based on his wide-ranging collecting efforts, Cei (1986) extended the range of L. rothi from southwestern Neuquén Province southward through southwestern Rio Negro Province to the east, including the Somuncurá plateau, and northern Chubut Province. He was the first to propose a “ rothi group” composed at that time of L. rothi and L. ruizleali . In 2003, Etheridge and Christie redescribed Liolaemus rothi , and described L. sagei but they placed both in the more inclusive boulengeri group, which at that time was considered a subgroup of the montanus group, and included 26 species. Morando (2004) and Avila et al. (2006) were the first to propose a rothi complex that included several populations recognized as Liolaemus rothi and Liolaemus spp. Pincheira-Donoso et al. (2007), in the description of L. hermannunezi , were the first to compare morphologically four of the species here included as members of the rothi complex, but they did not include L. loboi , described earlier by Abdala (2003). In a recent paper, Abdala et al. (2012) described a new species of the complex, but on the basis of morphological characters, they were unable to resolve the internal relationships of their “ telsen group”. This species complex still includes additional candidate species in Neuquén and Rio Negro provinces (Olave et al., in prep.), and based on all the available information published in the last few years, there is no doubt that lizard diversity in Neuquén province remains substantially underestimated. Based on current studies in progress, we expect that the number of lizard’s taxa will increase by 15–20 % in the near future, and we hope that an improved understanding of the evolutionary diversity of this conspicuous group of Patagonian vertebrates, will contribute to conservation efforts aimed at preserving the natural regions of the province.




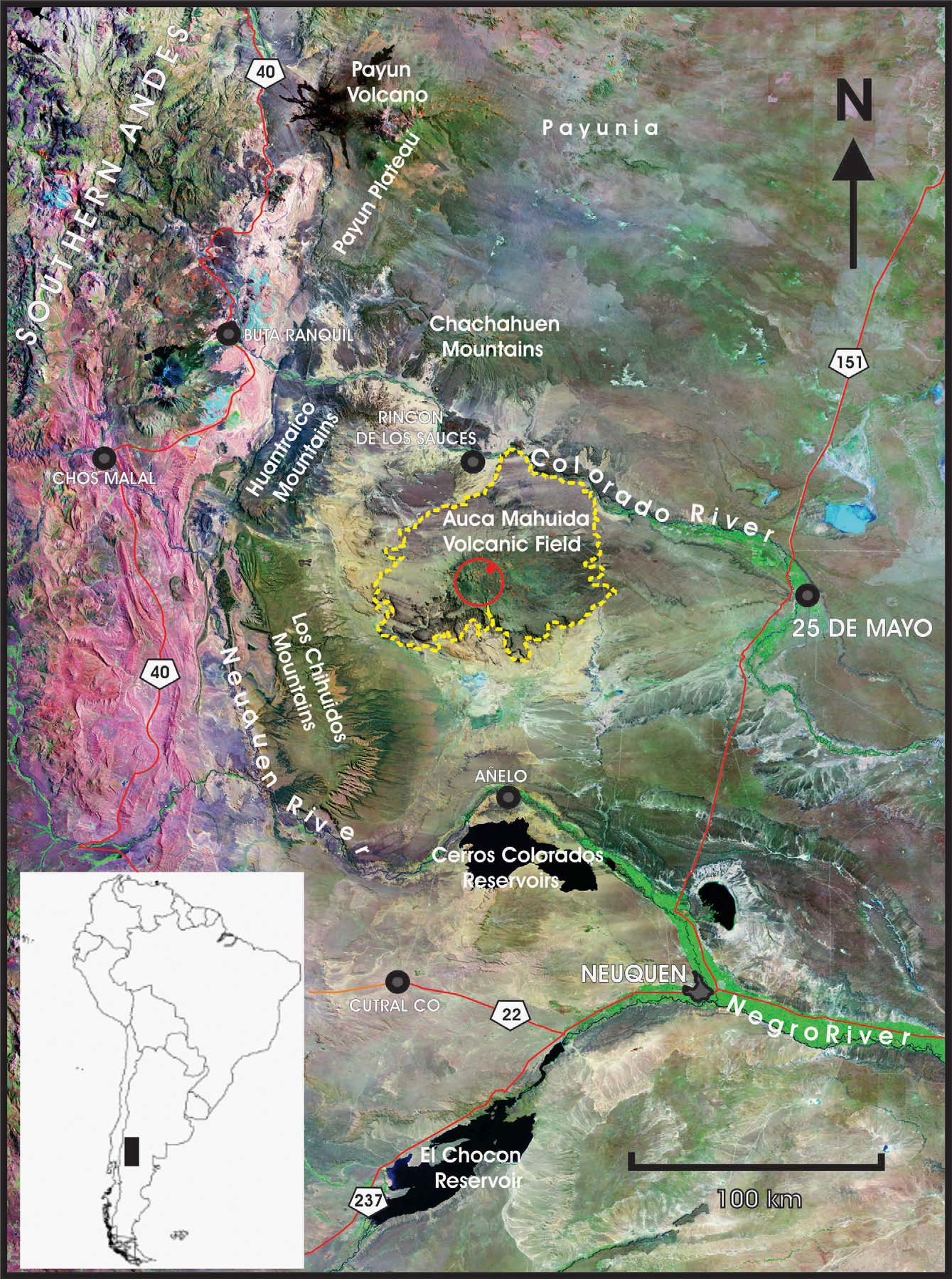

TABLE 3. Morphometric and meristic variation in Liolaemus sitesi sp. nov. type series. Means and standard deviations (SD) of the main morphometric and meristic characters. Measures in mm.
| Males (N= 11) | Females (N= 11) | |||
|---|---|---|---|---|
| Mean | SD | Mean | SD | |
| Snouth-Vent Length | 71.57 | 7.63 | 66.36 | 5.25 |
| Axilla-groin distance | 33.75 | 3.70 | 32.47 | 4.04 |
| Head length | 15.26 | 1.23 | 13.70 | 0.81 |
| Head width | 12.36 | 1.03 | 11.20 | 0.72 |
| Head height | 8.67 | 0.78 | 7.54 | 0.51 |
| Foot length | 21.64 | 1.17 | 20.05 | 1.05 |
| Tibial length | 14.40 | 1.19 | 12.70 | 0.61 |
| Arm length | 21.37 | 1.19 | 19.63 | 1.02 |
| Midbody scales | 71.63 | 3.41 | 72.45 | 2.46 |
| Dorsal scales | 81.27 | 2.64 | 82.81 | 2.31 |
| Ventral scales | 99.27 | 2.93 | 103.63 | 4.34 |
| Fourth toe lamellae | 27.09 | 1.30 | 27.45 | 1.69 |
| Third finger lamellae | 20.18 | 1.25 | 20.00 | 1.09 |
| Supralabial scales | 8.54 | 0.93 | 7.63 | 0.67 |
| Infralabial scales | 5.72 | 0.46 | 5.63 | 0.50 |
| Cloacal pores | 8.22 | 1.16 | - | - |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.