
Kuqaia, Li, 1993
|
publication ID |
https://doi.org/ 10.11646/zootaxa.5343.1.6 |
|
publication LSID |
lsid:zoobank.org:pub:C69D0F8C-E012-4396-9B1E-90BBCC505459 |
|
DOI |
https://doi.org/10.5281/zenodo.8329549 |
|
persistent identifier |
https://treatment.plazi.org/id/BA06879F-FF8B-F025-70DE-13DEFE33FC06 |
|
treatment provided by |
Plazi |
|
scientific name |
Kuqaia |
| status |
|
Kuqaia is not a cladoceran
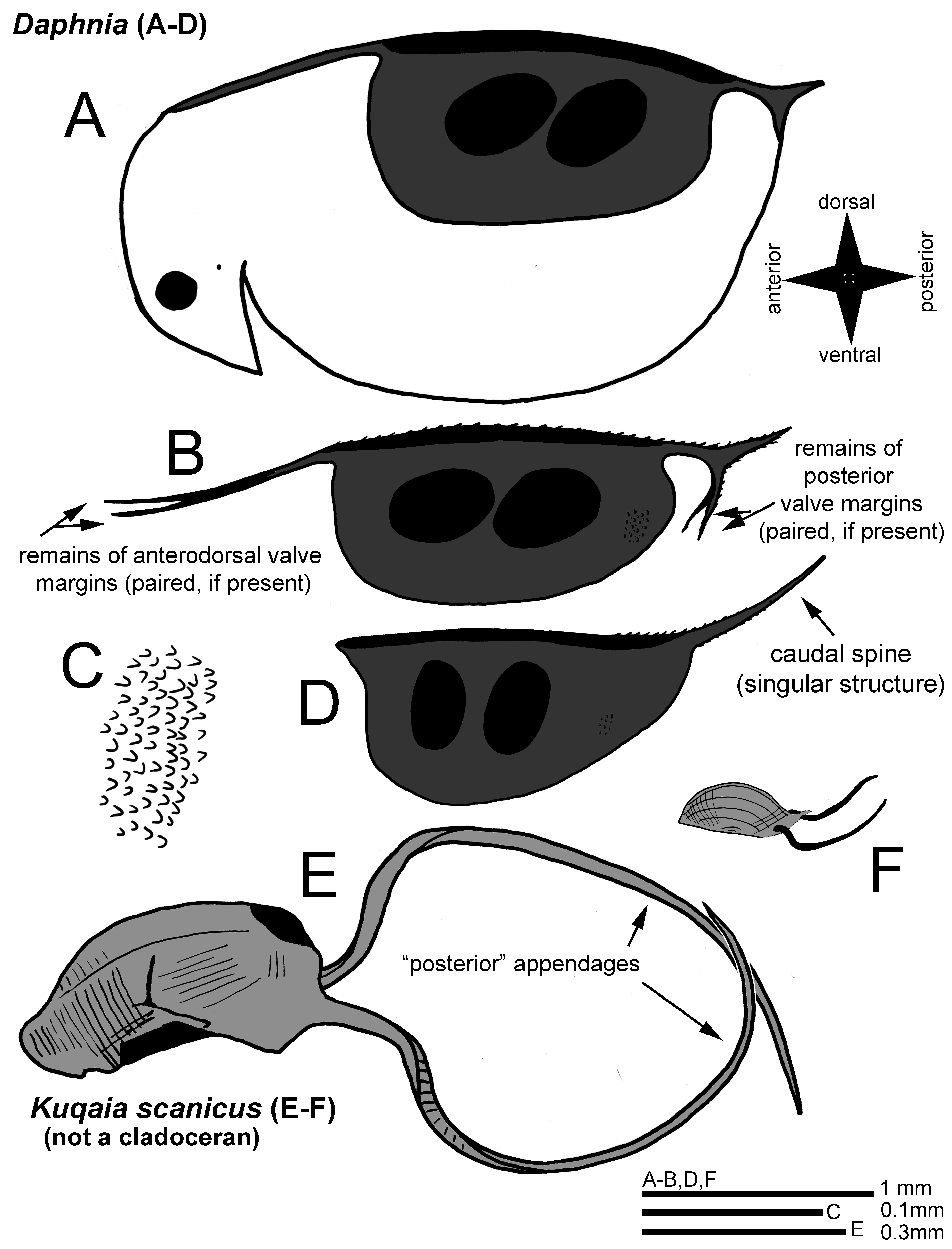
It is difficult to find and to assess the identity of fossils of small-sized extinct taxa in ancient strata ( Cooper et al. 2006). In earlier studies mentioned by Peng et al. (2023), Kuqaia has been attributed to fossil megaspores, gastropods, and the authors mention that these have also been suggested as potentially rotifers, or cocoons of annelids. The taphonomy of the deposits where Kuqaia is generally found ( Peng et al. 2023), indicates a predominantly marine environment with potential freshwater stadia during regressions, which might have been inhabited by cladocerans as these animals were present at the time in truly freshwater habitats (yet if so, they were mostly accompanied by other freshwater invertebrates in the taphonomic setting; Kotov 2009a; Van Damme & Kotov 2016; Van Damme et al. 2022). However, it is clear that the Kuqaia fossils morphologically are not cladoceran ephippia. Based on an examination of the SEM images and interpretations (drawings and text) in Peng et al. (2023) compared to ephippia ( Fig. 1 View FIGURE 1 ), the following observations can be made:
(i) Appendages (“peduncles”). The long appendages (“peduncles”) in K. scanicus are implanted at some distance from a potential “keel” and almost midway a straight margin visible in the fossils ( Fig. 1E View FIGURE 1 and Peng et al. 2023: Fig. 2a; the appendages are on the left in their study and the “keel” is oriented downwards in their figure, also visible in their Fig. 2b). The authors describe that the implantation of these peduncles is at “ the lateral part of the postventral end ”. In all Anomopoda , paired filiform appendages of the ephippia, if present, are the torn off continuations of the bivalved body margin, which are directly a continuation of the dorsal margin in the Daphniidae ( Figs 1A–B View FIGURE 1 ). Among the ephippial appendages, which may extend posteriorly and anteriorly, there can be the (entirely or partly) posterior caudal spines in daphniids (if present; Figs 1A–B and D View FIGURE 1 ) as well as the remnants of the dorsal and posterior valve margins, often with serrations/spinules in the dorsal section and on the spines ( Fig. 1B View FIGURE 1 ; see drawings in Benzie 2005; Mergeay et al. 2005). If present, caudal spines in daphniids can be singular and implanted posterodorsally (in Daphnia ) or paired and implanted posteroventrally (e.g., in Scapholeberis Schoedler, 1858 ). In the subgenus Daphnia (Ctenodaphnia) , because of the dorsal W-shaped structure of the headshield, the anterior appendages can be quite long ( Figs 1A–B View FIGURE 1 ; Benzie 2005), but it is not so in the subgenus Daphnia (Daphnia) ( Fig. 1D View FIGURE 1 ). In other anomopods like chydorids, a torn margin can also be present ( Scourfield 1902; Smirnov 1971). In bosminids, the valve projections (caudal spines or mucros) are implanted posteroventrally and often they are preserved in subfossil ephippia ( Scourfield 1901). The interpretation by Peng et al. ( Fig. 1F View FIGURE 1 ; drawings by Peng et al. 2023: Fig. 4) is that these appendages (“peduncles”) in Kuqaia are implanted laterally and submarginally at the “posterior” end. If this fossil would be an anomopod, any long filiform paired appendages would be continuing directly from the dorsal margin ( Figs 1A–B View FIGURE 1 ), not at a distance below it, and not midway on a straight posterior margin which is at a right angle with either the dorsal or the ventral margins ( Fig. 1E View FIGURE 1 ). Peng et al. (2023) compare their fossils with ephippia of Daphnia (Daphnia) pulicaria Forbes, 1893 and Daphnia (Ctenodaphnia) similis Claus, 1876 and write erroneously that “ the two shorter peduncles of D. similis originate from the central part of the postventral end ”. But the area where any appendages are implanted on the ephippia of D. similis is not the post-ventral end, it is dorsal as in all Daphnia , if appendages are present ( Figs 1A–B and D View FIGURE 1 ), and these are most certainly not homologous structures to the appendages in Kuqaia ( Fig. 1E View FIGURE 1 ). Ephippia of D. magna Straus, 1820 ( Figs 1A–B View FIGURE 1 ) look relatively similar to those of D. similis , yet the latter has a comparatively larger caudal spine as a (singular) projection, extending posteriorly from the posterodorsal corner of the body (e.g., Mergeay et al. 2005). Long paired filiform appendages in D. similis ephippia (if any) would be in an anterior position (as in Fig. 1B View FIGURE 1 ), while the long unpaired caudal spine would be posterior ( Figs 1B and D View FIGURE 1 ). In addition, the appendages in Kuqaia show a broad base ( Fig. 1E View FIGURE 1 ), almost a third of the entire margin on which they are implanted while in anomopods paired ephippial appendages are narrow and thin, as a continuation of the dorsal margin ( Fig. 1B View FIGURE 1 ). There are more arguments, e.g., the absence of any spinules on these appendages in Kuqaia ( Fig. 1E View FIGURE 1 ) which are often present in daphniids ( Fig. 1B View FIGURE 1 ), and the fact that these “peduncles” seem actually very robust structures, sharply pointed at the terminal ends in Kuqaia . In cladocerans, any paired ephippial filiform appendages that are remnants of the valve margins, are flimsy and would never be acute in such a way, while caudal spines (if present) would be implanted differently as stated earlier (dorsally in Daphnia ; Figs 1B and D View FIGURE 1 ). So, not only are these appendages not homologous after re-interpretation, the original suggestion of similarity in position and implantation was based on an erroneous interpretation of Daphnia ephippia by Peng et al. (2023), who confused ventral and dorsal sides of the cladoceran body as well as types and structures of ephippial appendages.
(ii) Dorsal keel. A narrow “dorsal” keel on the body is not uncommon in Anomopoda (e.g., Celsinotum Frey, 1991 in the Chydoridae , or the Macrothrix paulensis- complex in the Macrothricidae ; Frey 1991; Kotov et al. 2015). In several adult Daphniidae , for example in many members of the Ctenodaphnia Dybowski & Grochowski, 1895 lineage in Daphnia , the dorsal midline may often carry a series of small teeth ( Fig. 1B View FIGURE 1 ) of which the expression may be more intense in the presence of predators (e.g., Daphnia atkinsoni -group). While there is sometimes a dorsal keel with a smooth or serrated margin in anomopods, it is not likely to find a ventral keel with an even margin in anomopod ephippia; either the ventral margin of the body is present (with remnants of structures), or when the ventral margins of the valves are torn off like a thread, it leaves an irregular appearance on the remaining ventral portion, like in many chydorids or macrothricids. In daphniids, the ephippial structure does not reach the ventral body margin entirely ( Fig. 1A View FIGURE 1 ). None of these features are visible in K. scanicus, where clearly a thick smooth keel or flange is present (up in Fig. 1E View FIGURE 1 ; oriented downwards in Peng et al. 2023: Figs 2a–b). It does not correspond to the morphology of a dorsal keel or margin, or any ventral margin, in Anomopoda . In fact, it is not clear from the Kuqaia fossil images whether these are actually bivalved structures ( Figs 1E–F View FIGURE 1 ). In Cladocera (not in the raptorial lineages), the bivalved carapace is quite obvious.
(iii) Valve Ornamentation. Anomopoda have a wide range of types of valve ornamentation, from smooth to pustulate (verrucose) to broadly reticulate, striate or with a fine mesh of striae ( Smirnov & Kotov 2009). Such reticulation is relatively better developed in benthic species and less expressed in zooplankters ( Kotov 2006). This is the case in Daphniidae , which may often have a smooth or a dense reticulate pattern on the valves in adults. In most ephippia, the ornamentation becomes much more obvious, because this structure is heavily chitinized. This gives daphniid ephippia generally a densely ornamented surface with small projections ( Fig. 1C View FIGURE 1 ) or a dense reticulation, which is also visible in fossil daphniid ephippia from the Jurassic-Cretaceous boundary ( Kotov 2009a; Van Damme & Kotov 2016). The ornamentation of Kuqaia is entirely different, consisting of grooves and ribs, and it does not correspond to the ornamentation in any anomopod (in fact, any branchiopod). In K. scanicus, parallel lines are at a right angle of the long (“ventral” and “dorsal”) margins or parallel to the “frontal” margin and these lines form some small ridges, giving it a ribbed appearance ( Fig. 1E View FIGURE 1 ; Peng et al. 2023: Fig. 3a); the latter authors state that this ornamentation in Kuqaia “ is suggestive of a morphologically simple, archaic group of cladocerans, possibly on the stem group lineage of Daphnia ”. Other Kuqaia fossils shown in Peng et al. (2023) have strong concentric or radiating deep lines and perforations on the lateral surfaces (K. concentrica Li, 1993, K. radiata Li, 1993), which are deep. Even the appendages in K. scanicus have a marginal row of indentations or transversal ridges ( Fig. 1E View FIGURE 1 ; Peng et al. 2023: Fig. 3c). None of these ornamentations correspond to types known in any anomopod including fossil ephippia from the Jurassic-Cretaceous boundary ( Kotov 2009a) which have a morphology congruent with extant daphniid and moinid species-groups ( Van Damme & Kotov 2016). The general ornamentation of Kuqaia falls, in no way, within any variability of known anomopods, and definitely not within the Daphniidae . The suggestion that this is an “archaic” type of cladoceran ornamentation, is here rejected. “Ribbed” cladoceran ornamentation is unknown – if there are any long continuous concentric parallel lines in the group, it is from the retention of earlier instar carapaces during the moulting process, like in the anomopods Ilyocryptus Smirnov, 1976 or Monospilus Sars, 1861 ( Kotov 2006) . But this is not the case here either. In addition, the double row of indentations on the inner side of each of the appendages in Kuqaia also does not make any sense if these were daphniid-like anomopods (there are no transversal indentations on the appendages but marginal structures such as denticles and short setules; Fig. 1B View FIGURE 1 ).
(iv) Embryo chambers/loculi. In the majority of anomopods and, in particular daphniids and (their sister lineage) moinids, the approximate shape and size of the embryo is visible (to various extent) in both extant and fossil ephippia by the convex expressions of the spheroid embryo chambers, sometimes called loculi ( Kotov 2009a; Kotov et al. 2019). These bulges in the ephippia can be less visible for Chydoridae and Eurycercidae ( Scourfield 1902) . Yet, even in chydorids, there is an embryonal space visible from the lateral outside of the shell as a general protuberance. In all Daphniidae and Moinidae the shape and orientation of the diapausing embryos is therefore very clear, and together with other characters such as the surface ornamentation, it can be used as a diagnostic feature ( Kotov et al. 2019); in Daphniidae and Moinidae there are one to two embryos (sometimes none), not more. In Kuqaia , all impressions or external morphology revealing the shape or size of potential diapausing embryos are absent. Peng et al. (2023) mention the absence of such loculi as a taxonomical feature of Kuqaia , but in Daphnia or related lineages some shape of the embryonal chambers would be visible from outside. In addition, the fossil lacks all features of ephippia that may not have strongly expressed loculi, such as Eurycercidae , which have heavily chitinized carapaces. The absence of loculi is not a differential character in this case, it is merely because they are not anomopods. In general, the shape of these Kuqaia fossils does not correspond, in any way, to ephippia.
From the above, it is clear that Kuqaia is a not an ephippium of a daphniid-like (or other) anomopod. In fact, it is quite obvious that this is not a cladoceran (only anomopods produce ephippia). Objectively, the Lower Jurassic fossils in the study of Peng et al. (2023) do not correspond in the abovementioned morphological features, and a relationship with cladocerans is not realistic. In the past, other “cladoceran-like” fossils have been reported (e.g., Adamczak 1965), yet more recently, these have been considered ambiguous records (e.g., all Paleozoic fossils originally attributed to the group; Van Damme & Kotov 2016). Without actual segmented appendages (e.g., antennae) or recognisable ephippial features, it is definitely not easy.
The characters discussed above include the type and orientation of the external ornamentation, the implantation and shape of the long appendages, the keel and (the lack of) embryonal impressions or loculi. These are not just diagnostic differential characters within the cladocerans – the Kuqaia fossils are simply not part of the group in any way. Arguments can be made about the size (300 µm without the appendages which would be very small for daphniids; see the size of Kuqaia relative to the two Daphnia ephippia in Fig. 1F View FIGURE 1 ) and the entire shape of the structure which does not correspond to known body shapes in cladocerans (or ephippia). The latter are generally conserved, in particular for ephippia (Kotov & Van Damme 2016).
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.