
Paramantoida amazonica, Agudelo, Antonio A., 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3797.1.14 |
|
publication LSID |
lsid:zoobank.org:pub:C1FD1B1B-326E-4BE4-B14C-30058ECF02B2 |
|
DOI |
https://doi.org/10.5281/zenodo.6141642 |
|
persistent identifier |
https://treatment.plazi.org/id/BA1F87BD-FF9A-B541-C2E6-F913FB64ACCE |
|
treatment provided by |
Plazi |
|
scientific name |
Paramantoida amazonica |
| status |
sp. nov. |
Paramantoida amazonica View in CoL n. sp.
( Figs. 1−7 View FIGURE 1 View FIGURE 2 View FIGURE 3 View FIGURE 4 View FIGURE 5 View FIGURE 6 View FIGURE 7 )
Type Material. Holotype ♂ ( Fig. 1 View FIGURE 1 ), BRAZIL: Amazonas, Manaus; Reserva Biológica do Cuieiras, ZF-2, S02°35'21'' W60°06'55'', 0 5 xi 2005, armadilha luminosa móvel, A. Agudelo & J. Rafael leg. Deposited at INPA, Invertebrate Collection (M.004).
Paratypes. (19 ♂): BRAZIL, Amazonas (AM): Presidente Figueiredo, Rodovia 240, km 24, Ramal São Francisco, 29–31 x 2008, J.A. Rafael, F.F. Xavier leg, armadilha luminosa dossel ( INPA); Presidente Figueiredo, Est. Balbina, S02°03'36'' W59°55'35'', 19-30 x 2003, F. Xavier & D. Vidal leg. ( INPA); Presidente Figueiredo, rod. 240 km 24, ramal São Francisco, 29–31 x 2008, armadilha luminosa, J.A. Rafael & F. Xavier leg, ( INPA); 2♂ Barcelos, Rio Padauarí, Comunidade Ararinha, N00°30'18'' W64°03'30'', 0 6 vi 2010, armadilha luminosa, A. Agudelo & J. Rafael leg, ( MZUSP); Barcelos, Rio Araça, Comunidade Bacuquara, N00°09'17'' W63°10'35'', armadilha luminosa, A. Agudelo & J. Rafael leg, (NHM-London); Barcelos, Igarapé Ererê, Coruja, N00°06'16'' W63°51'01'', 18–25 vi 2008, F. Xavier leg, armadilha luminosa ( INPA); Barcelos, rio Ererê, Coruja, N00° 06'16'' W63°51'01'', 12–25 viii 2009, F. Xavier & G. Monte leg, armadilha luminosa ( INPA); Barcelos, Rio Deminí, S00°18'05'' W62°45'56'',? viii 2008, R. Machado leg, ( INPA); Manaus, Rod. AM 0 10, km 50, ZF- 2 km 27, Próx. Entrada LBA, S02°35' W60°06', 5 iii 2011, 18–21:00 hrs, armadilha luminosa móvel, J.T. Camara & J.A. Rafael leg, Coleção Entomológica Pe. Jesus Santiago Moure ( DZUP); Manaus, Reserva Biológica do Cuieiras ZF-2, S02°35'21'' W60°06'55'', 0 5 xi 2005, armadilha luminosa móvel, A. Agudelo & J. Rafael leg, ( INPA); Manaus, Br. 174, ZF- 2 km 50, 06–09 viii 2001, R. Guimarães leg, ( INPA, 2933); Manaus, km 12, AM 240,? vi 2007, armadilha luminosa, G Lourido & F. Xavier ( INPA); Manaus, Torre da ZF-2 (45m), S02°35'20'' W60°06'55'', 06-09 i 1997, C. S. Motta & J. Vidal leg, ( INPA, 0016963); Manaus, Rod. AM 0 10, km 50, ZF- 2 km 14, Torre (40m), S02°35' 21'' W60°06'55'', 0 3 iii 2011, 18–21:00 hrs, armadilha luminosa dossel, F. Xavier & J. Câmara leg ( INPA); Manaus, ZF- 2 km 34, Campina, S02°35' 37' W60°12' 39', 11 vii 2008, luz, J.A. Rafael & F. Xavier leg, ( INPA); Manaus, ZF- 2 km 14, Torre (40m), S02°35'21'' W60°06'55'', 18–21 ii 2004, (UV) Black Light, J. Rafael & C. Motta leg, ( INPA); Manaus, rod. Am 240 km 24, S02°00' 55' W59°49' 40',? viii 2005, light, F. Xavier leg, ( INPA). Pará (PA): Rio Trombetas, Oriximiná, ALCOA mines, Monte Branco, 0 5 x 1982, (UV) Black Light ( INPA, 00272552).
Additional material: VENEZUELA: 2♂, T.F. Amaz., Cerro de la Neblina, Basecamp. 0°50'N, 66°9'44"W, 140 m., 1-10 March 1984, D. Davis & T. McCabe ( USNM) (G. Svenson, pers. com.).
Description (based on holotype and paratypes, all males). General colouration of body dark brown, appearing almost black, with various white-coloured marks.
Head. Vertex convex, with a narrow white line on the epicranial suture ( Fig. 3 View FIGURE 3 b); ocellar region black; antennal scapes with a ventro-lateral white spot; proximal flagellomere slightly longer than the next three flagellomeres taken altogether. Juxtaocular region near compound eyes with two distinct white areas (anterior and posterior); area between antennal insertion and eyes with one small elliptical tubercle. Frontal shield transverse, almost five times as wide as long ( Fig. 3 View FIGURE 3 a) and dorsal edge roundly convex. Maxillary and labial palpi exhibiting white stripes.
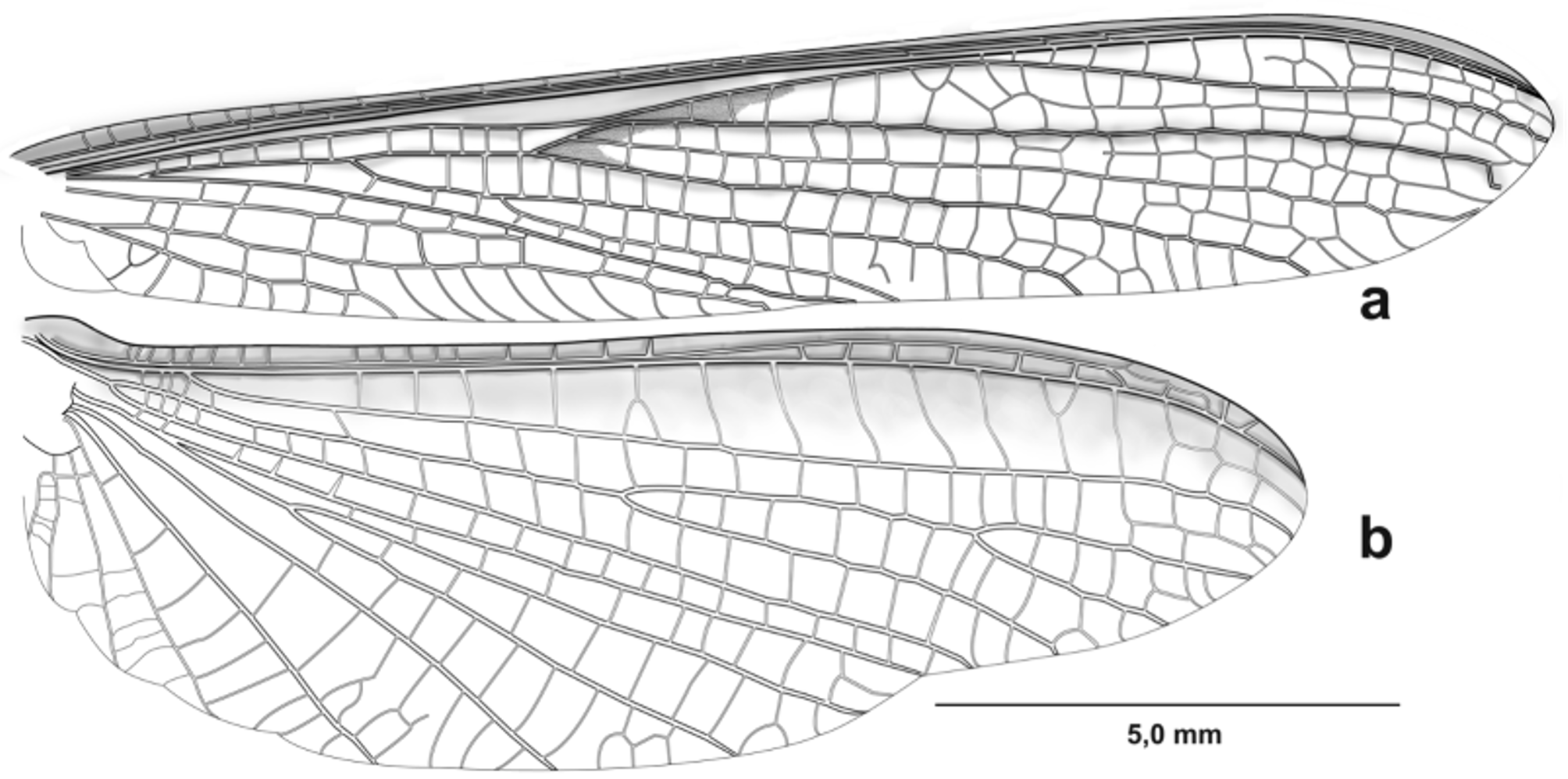
Thorax. metazone 1.2 times longer that prozone, lateral and distal margins of metazone white ( Fig. 3 View FIGURE 3 b). Dorsal and ventral margins bearing setae. Anterior margin of forefemora with few setae, 10–12 antero ventral spines, the first proximal nine arranged in zigzag, followed by several setae on the margin; 3 discoidal spines and 0–2 small, postero ventral spines (spinules); all spines in forefemora entirely black ( Fig. 4 View FIGURE 4 ). Foretibiae slightly curved outward on dorsal view, with white dorsoproximal maculae; 5–6 antero ventral spines, 3–4 posterio ventral spines located distally and several long setae along the proximal region; all spines of foretibiae entirely black. Foremetatarsi about 1,4 times the foretibiae, with a small white basal area and slightly longer than the remaining tarsomeres taken all together. Episternum II-III and epimeron II-III with white maculae ( Fig. 1 View FIGURE 1 ). Meso- and metathoracic femora with a single apical spine; meso- and metathoracic tibiae with two apical spines. Meso- and metathoracic metatarsi with a white proximal area. Mesothoracic wings hyaline, longer than abdomen; costal area opaque; discoidal area with brown macula between medial vein and the ramification of CuA1-CuA3 ( Fig. 5 View FIGURE 5 a). Metathoracic wings hyaline, exhibiting a longitudinal brown stripe along costal area that fades before reaching the apex of the wing ( Fig. 5 View FIGURE 5 b).
Abdomen. ( Fig. 6 View FIGURE 6 a): distal margin of T5, S2–4, lateral margins of T2–3 and last three sternites exhibiting white marks. Supra-anal plate subtriangular. Sub-genital plate exhibiting white lateral margins. Cerci with 13 joints, proximal cercomere slightly longer than any remaining one ( Fig. 6 View FIGURE 6 b).
External genitalia. ( Fig. 7 View FIGURE 7 ): right epiphallus ( Fig. 7 View FIGURE 7 a) with sclerites of ventral membrane well-defined and strongly sclerotized; ventral lobe hook-shaped, flattened; ventral plate (vp) ridge-shaped, connected to another elongate sclerotized area, distally pointing towards the right; apex of mid-arm (= braccio mediale del fallomero in La Greca 1954) (ma) forming a strong, thumb-shaped, sclerotized lobe with short spines ( Fig. 7 View FIGURE 7 a). Left epiphallus and hypophallus complex (leph/hyph, Fig. 7 View FIGURE 7 b) with anterior process (ap) pointing towards the right, gradually sclerotized to form a hard lobe, basal section hook-shaped; pseudophallus (psph) well-developed, sinuous, tildeshaped (~), distal part curving upwards with terminal spines giving a serrated appearance; titillator (ti) smooth and curved dorsally; ventral phallomere or hypophallus (hyph) U-shaped (inverted), distal lobe (dl) without specialization, smooth, and curved dorsally ( Figure 7 View FIGURE 7 b).
Measurement (mm) of type series (holotype in [bold]): Body length (head to tip of abdomen) 18–20 [19.2]; head (vertex to distal margin of labrum) 2.9–3.2 [3.0]; distance between lateral margins of the eyes 3.8–3.9 [2.4]; length of pronotum 2.0–2.3 [2.2]; length of prozone 0.8–1.0 [1.0]; length of metazone 1.2–1.3 [1.2]; length of prothoracic length of coxae; 2.4–2.6 [2.6]; length of prothoracic femora 3.0–3.3 [3.3]; length of prothoracic tibiae 2.0–2.2 [2.2]; length of mesothoracic femora 4.0–4.3 [4.2]; length of Mesothoracic tibiae 3.2–3.6 [3.5]; length of Metathoracic femora 5.4-5.9 [5.9]; length of metathoracic tibiae 5.0–5.4 [5.4]; length of mesothoracic wing (base to apex) 17.5–17.9 [17.9]; length of metathoracic wings (base to apex) 14.9–15.2 [14.9].
Female. unknown.
Etymology. The specific name alludes to the current natural distribution of the species, particularly along the middle and lower Rio Negro in the Brazilian central Amazon ( Fig. 8 View FIGURE 8 ).
Comparative notes. Paramantoida is unique among other Neotropical taxa as it exhibits some characters unusual for what is generally observed in most Mantodea . One striking and unique feature is its dark coloration with contrasting white spots. This color pattern is observed in certain wasps, thus Paramantoida perhaps constitutes an example of Batesian mimicry and might be part of a mimicry complex of one or more unidentified aggressive wasps. Some authors have previously suggested wasp mimicry in Mantoida ( Jackson & Drummond 1974; Deyrup 1986). Nonetheless, this particular combination of colors (i.e. black and white) is not known in any other Neotropical genera, except perhaps in lichen mantids of the genus Pseudopogonogaster ( Thespidae , Miopteryginae ), whose species intraspecifically exhibit various colour morphs, including a contrasting black and white form, a pattern most likely used for camouflage on lichen-covered trunks (e.g. Salazar & Carrejo 2002; Rivera et al. 2011). The most outstanding feature of Paramantoida is the variable nature of the postero ventral spines on the forefemora, which are normally absent but, when present, no more than two have been observed; these might actually be considered spinules (sensu Grimaldi 2003) rather than spines. Similarly, the number of postero ventral spines in the foretibiae is three (as in most specimens examined), whereas in Mantoida , no less than four spines have ever been recorded in both femora and tibiae. A similar reduction in the number of spines is only seen in species of the African, Asian and Australian family Amorphoscelidae , as well as in some Thespidae (e.g. Oligonicinae , Thesprotia Stål ), where there may only exist as few as one postero ventral spine on the forefemora. This unique set of characters exhibited by Paramantoida suggests that a reduction in the number of spines in the forelegs might be a trend within this lineage of Mantoididae . The lack of postero ventral spines in the forefemur in Paramantoida contrasts with the fossil record, which have revealed the existence of all sets of spines in the forelegs in extinct species from the Cretaceous ( Grimaldi 2003), many of which are morphologically close to Mantoididae species.
Paramantoida amazonica View in CoL is the largest species among the known Mantoididae View in CoL ; it is slightly but notoriously larger than M. nitida Newman, 1838 View in CoL , M. brunneriana View in CoL and M. fulgidipennis View in CoL ( Fig. 9 View FIGURE 9 a), and about twice the size of M. tenuis (Perty, 1833) View in CoL , M. luteola View in CoL ( Fig. 9 View FIGURE 9 b) and M. beieri ( Kaltenbach 1957) . Sexual dimorphism in Mantoididae View in CoL is inconspicuous, with females being primarily identified by their slightly larger size and more robust appearance. Some females of M. brunneriana View in CoL can reach up to 18 mm, and it is possible that females of P. amazonica View in CoL might exceed 20 mm.
The general morphology of the phallic complex of P. amazonica View in CoL is typical of what is known for Mantoididae View in CoL . The pseudophallus is large in P. amazonica View in CoL , whereas in those species of Mantoida View in CoL where the genitalia is known, the pseudophallus is short and almost triangular as in M. brunneriana View in CoL ( Fig. 9 View FIGURE 9 d), M. argentinae View in CoL and M. schraderi View in CoL ( La Greca & Lombardo 1989, Cerdá 1993; Klass 1997). Interestingly, Cerdá (1993) illustrated the pseudophallus of what he considered an undescribed species of Mantoida View in CoL , very similar to that of P. amazonica View in CoL .







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Paramantoida amazonica
| Agudelo, Antonio A. 2014 |
M. beieri (
| Kaltenbach 1957 |
M. nitida
| Newman 1838 |
M. tenuis
| Perty 1833 |