
Decidia Stål, 1875
|
publication ID |
https://doi.org/10.5281/zenodo.187440 |
|
publication LSID |
lsid:zoobank.org:pub:561D49DB-4333-4B3B-BE75-F50D6608E5E3 |
|
DOI |
https://doi.org/10.5281/zenodo.6213759 |
|
persistent identifier |
https://treatment.plazi.org/id/BD228785-FFCA-8529-FF66-FABEFCCCFE4F |
|
treatment provided by |
Plazi |
|
scientific name |
Decidia Stål, 1875 |
| status |
|
Decidia Stål, 1875 View in CoL
Type-species: Phasma soranus Westwood 1859: 127 , pl. 17: 3, by monotypy. Decidia Stål, 1875: 57 , 96.
Brunner v. Wattenwyl, 1893: 98.
Rehn, 1904: 91.
Kirby, 1904: 403.
Redtenbacher, 1906: 97.
Beier, 1957: 429.
Bradley & Galil, 1977: 203.
Bragg, 2001: 632.
Conle & Hennemann, 2002: 45, pl. 1: 9 & 8: 81.
Zompro, 2004: 140, 309, fig. 78.
Otte & Brock, 2005: 119.
Autolyca, Caudell, 1906: 192 View in CoL .
Redtenbacher, 1906: 95 (in part).
Conle & Hennemann, 2002: 34 (in part).
Zompro, 2004: 151 (in part).
Otte & Brock, 2005: 57 (in part). Brachyelena Hebard, 1933: 31 , pl. 2: 8. [Synonymised by Zompro, 2004: 140]
Bradley & Galil, 1977: 203.
Bragg, 2001: 630.
Zompro, 2004: 140, 306. [ As a synonym of Decidia Stål, 1875 ]
Otte & Brock, 2005: 109. [ As a synonym of Decidia Stål, 1875 ] Peruphasma Conle & Hennemann, 2002: 95 (in part).
Zompro, 2004: 146 (in part).
Otte & Brock, 2005: 260 (in part).
Conle & Hennemann, 2005: 61 (in part).
Murányi 2007: 57 (in part).
Phasma, Westwood, 1859: 127 View in CoL , pl. 17: 3 (in part).
Description ( Ƥ, 3): Medium-sized (body length Ƥ 38.0 – 59.0 mm, 33 34.0 – 46.5 mm), very robust Anisomorphini with very long legs and tarsi. Body surface smooth and dull to slightly shiny; finely hirsute (ventrally in particular). Ƥ with mesothorax broadened towards the posterior and abdomen swollen medially, 33 of ± uniform width. Either apterous or with scale-like tegmina and well developed alae, which however do not reach the apex of the abdomen.
Head: Almost round in dorsal aspect, about as wide as long, and slightly compressed dorsoventrally. Cheeks convex, vertex smooth and rather flattened. Eyes round, very prominent and projecting hemispherically; their length contained no more than 2x in that of cheeks. Ocelli either well developed to strongly reduced and rudimentary. Antennae long and filiform, distinctly longer than the body and projecting over apex of abdomen. Scapus oval in cross-section and indistinctly longer than wide. Pedicellus narrowed towards the apex, cylindrical and considerably shorter than scapus. All following antennomeres cylindrical in cross-section. III ± elongated and longer than pedicellus. IV much shorter than III, the following increasing in length. All antennomeres finely bristled.
Thorax: Pronotum a little longer but narrower than head, slightly longer than wide and the anterolateral angles with a prominent semi-circular glandular gap. Mesothorax very gently (33) or decidedly widened towards the posterior (Ƥ). Mesonotum less than 2x the length of pronotum; 1.3 – 1.5x longer than wide in Ƥ and 1.7 – 2 x longer than wide in 33. Mesosternum simple and smooth. Metathorax about equal in length but slightly wider than mesothorax. Metanotum wider than long and equal in length or shorter than median segment. Tegmina (if present) scale-like, oval, with a very shallow central hump, at best 2x longer than wide and slightly projecting over posterior margin of metanotum. Alae (if present) at best reaching to abdominal segment VII. Anal region of alae transparent pale brown, grey or slightly orange. Costal region of alae and tegmina with distinct pale net-like venature.
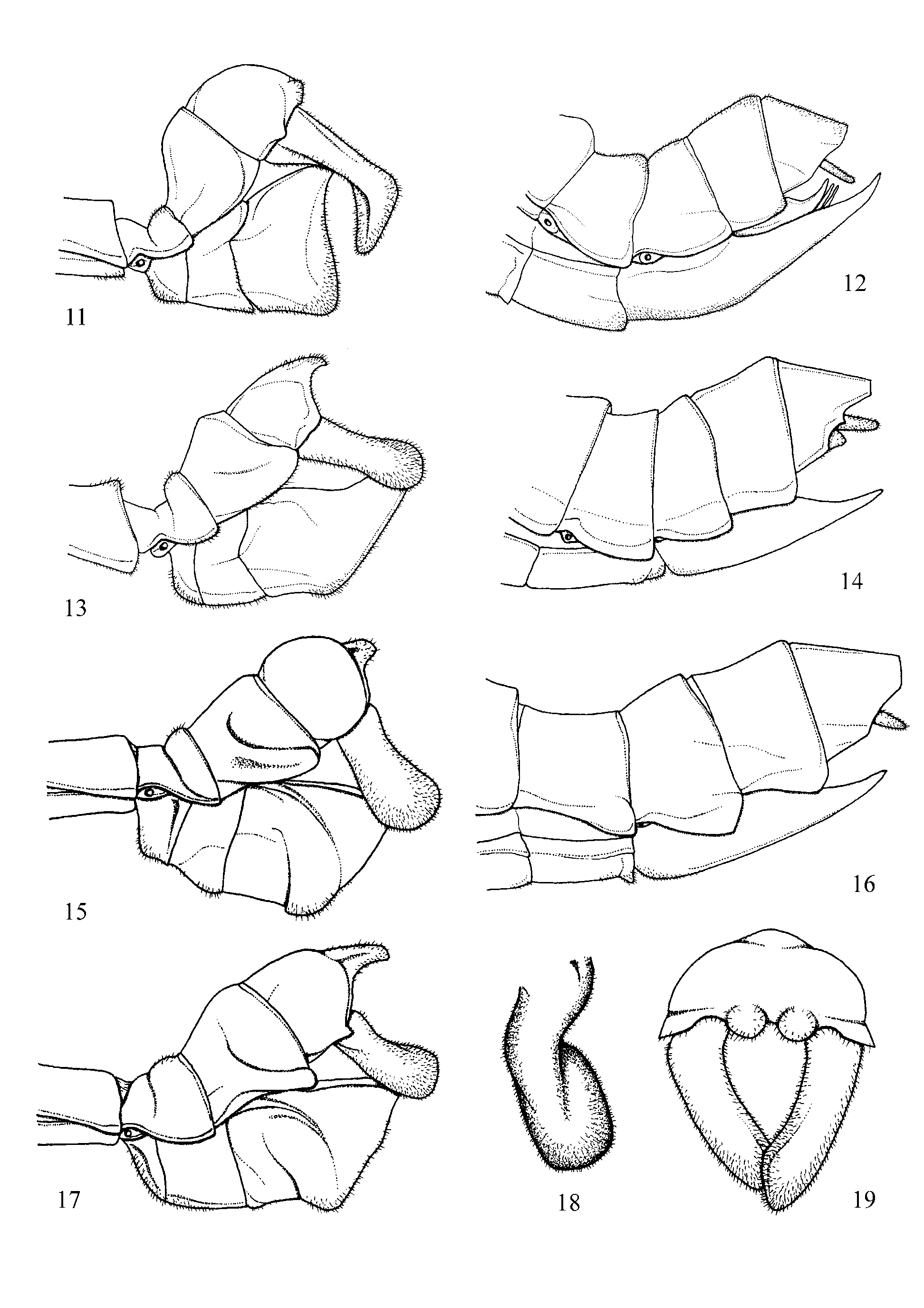
Abdomen: Round in cross-section, about 1.5x longer than head and complete thorax combined in Ƥ and about 1.7 – 2 x longer in 33. Distinctly broadened and swollen medially in Ƥ, much more slender and with segments II – VII roughly of uniform width in 33. Segments VIII – X much broader than previous and clublike in 33, narrower than previous in Ƥ. Median segment transverse in Ƥ and almost quadrate in 33; roughly equal in length to metanotum. Tergites quadrate to transverse with the later margins gently rounded (VII – IX in particular) in Ƥ, longer than wide and parallel-sided in 33. Sternites II – VII smooth and finely hirsute; praeopercular organ of Ƥ indistinct, represented only by a median slit or hole at posterior margin of sternum VII. Tergite IX of 33 with a ± distinct, diagonal bulge on both lateral surface which is directed posteriad and terminates at posterolateral angle of segment. Anal segment of Ƥ tapered towards a widely rounded apex, dorsally with a faint longitudinal median carina and lateral margins with a shallow concave excavation near bases of cerci. Anal segment of 33 bulgy with the lateral margins ± strongly deflexed and the lateral surfaces ± strongly convex or hemispherical. Posterior margin with a median projection of variable length and a notched apex; set with several minute spines interoventrally. Cerci of Ƥ small and much shorter than anal segment; round in cross-section and constricted towards the apex; finely bristled. Cerci of 33 prominently enlarged, longer than anal segment and densely bristled; shape variable. Constricted and cylindrical basally, broadened and ± laterally compressed in apical half, slightly angled inwards. Vomer well developed and sclerotised, longer than wide, roughly triangular and longitudinally separated into two asymmetrical parts. Apex pointed slightly curved towards the right and almost reaching to posterior margin of anal segment. Poculum of 33 very large and bulgy, deeply cup-shaped and ventrally projecting by more than height of tergite IX; posterior margin rounded and reaching to slightly projecting over posterior margin of anal segment. Subgenital plate of Ƥ boat to scoop-shaped, pointed apically and reaching to slightly projecting over posterior margin of anal segment; finely hirsute.
Legs: All very long and slender, with all femora and tibiae distinctly carinate; unarmed. Medioventral carina present on profemora, lacking on meso- and metafemora. Profemora straight and not compressed or curved basally, ranging from slightly shorter to longer than head, pro- and mesonotum combined. Mesofemora considerably longer than mesonotum. Metafemora at least reaching to abdominal segment VII and metatibiae prominently projecting over apex of abdomen. Tarsi very elongate and at least ½ the length of corresponding tibiae; heavily hirsute. Basitarsus at least 2.0x longer than 2nd segment.
Differentiation ( Table 1 View TABLE 1 ): Obviously closely related to the Central American Autolyca Stål 1875 (Typespecies: Autolyca pallidicornis Stål, 1875 ), with which it shares the general body shape, smooth body surface, large and round head, prominent eyes, long legs and tarsi, and straight not basally compressed or curved profemora. It however differs from Autolyca by: the dull and at best slightly shiny body surface; more distinct ocelli; longer antennae; relatively longer legs and tarsi, and more distinct carinae of the femora and tibiae of both sexes. 33 additionally differ by the much larger cerci and lacking the conspicuous finger-like lateral extension of abdominal tergite IX seen in Autolyca . This latter structure is merely represented as a diagonal bulge along the lateral surfaces of tergite IX in Decidia ( Figs. 11, 13, 15, 17 View FIGURES 11 – 19 ). While tegmina and alae may be present in Decidia , the known representatives of Autolyca are invariably apterous.
At first glance the winged species of Decidia ( D. soranus (Westwood) and D. magnifica n. sp.) strongly resemble certain species of Peruphasma Hennemann & Conle, 2002 (e.g. the Peruvian P. s c h u l t e i Hennemann & Conle, 2005). However, characteristic features such as the lack of a conspicuous, finger-like dextral appendage on sternite IX and prominently enlarged cerci of 33, or the more prominent ocelli and considerably longer legs and tarsi frequently distinguish representatives of Decidia from Peruphasma .
Comments: Bradley & Galil (1977: 203) placed Decidia in the tribe Anisomorphini were it was retained by Conle & Hennemann (2002: 45) who pointed out the close relation to the Central American Autolyca . Zompro (2004: 140) in contrast claimed relation to Neophasma Redtenbacher, 1906 , stated that “ Decidia species appear like very broad Neophasma species […]” and erroneously transferred Decidia to the tribe Pseudophasmatini . The inaccuracies and resulting problems of the arrangement of the subfamily Pseudophasmatinae and contained tribes Pseudophasmatini and Anisomorphini proposed by Zompro (2004) have already been pointed out and partly corrected by Murányi (2007: 57) and Conle & Hennemann (2008: 8). Zompro (2004: 131) distinguished between Pseudophasmatini and Anisomorphini merely on the basis that the profemora were at least as long or distinctly longer ( Pseudophasmatini ), or equal in length to shorter than the combined length of the head, pro- and mesonotum ( Anisomorphini ). However, the apparently very close relation between Decidia and Autolyca here shown clearly prove this feature to be of no use for distinguishing tribes within the Pseudophasmatinae . They are either as long as or longer than the head, pro- and mesonotum in Decidia and roughly equal in length to these segments combined in Autolyca . The close relation between Decidia and Autolyca is furthermore emphasized by features such as the general body shape, straight profemora, very long legs and tarsi, or strongly elongated basitarsus of both sexes. The lack of wings in Autolyca no longer holds true as a distinguishing feature since two apterous species, D. blapoides ( Redtenbacher, 1906) and D. doylei ( Caudell, 1906) , are here shown to belong in Decidia . Furthermore, 33 of both genera have conspicuously enlarged, characteristically shaped cerci, and the finger-like lateral extensions of abdominal tergite IX seen in 33 of Autolyca are at least represented by a lateral bulge in Decidia . Hence, Decidia is here removed from Pseudophasmatini and re-transferred to Anisomorphini ( rev. stat.). For a more detailed comparison and distinction between Decidia and Autolyca see table 1 below.
Brachyelena Hebard, 1933 was described from a unique Ƥ HT from Cauca, Colombia in ANSP. The typespecies B. hirsuta Hebard, 1933 is obviously conspecific to Decidia soranus ( Westwood, 1859) View in CoL and was synonymised by Zompro (2004: 140). Consequently, Brachyelena is a synonym of Decidia View in CoL .
The eggs of Decidia View in CoL are as yet unknown.
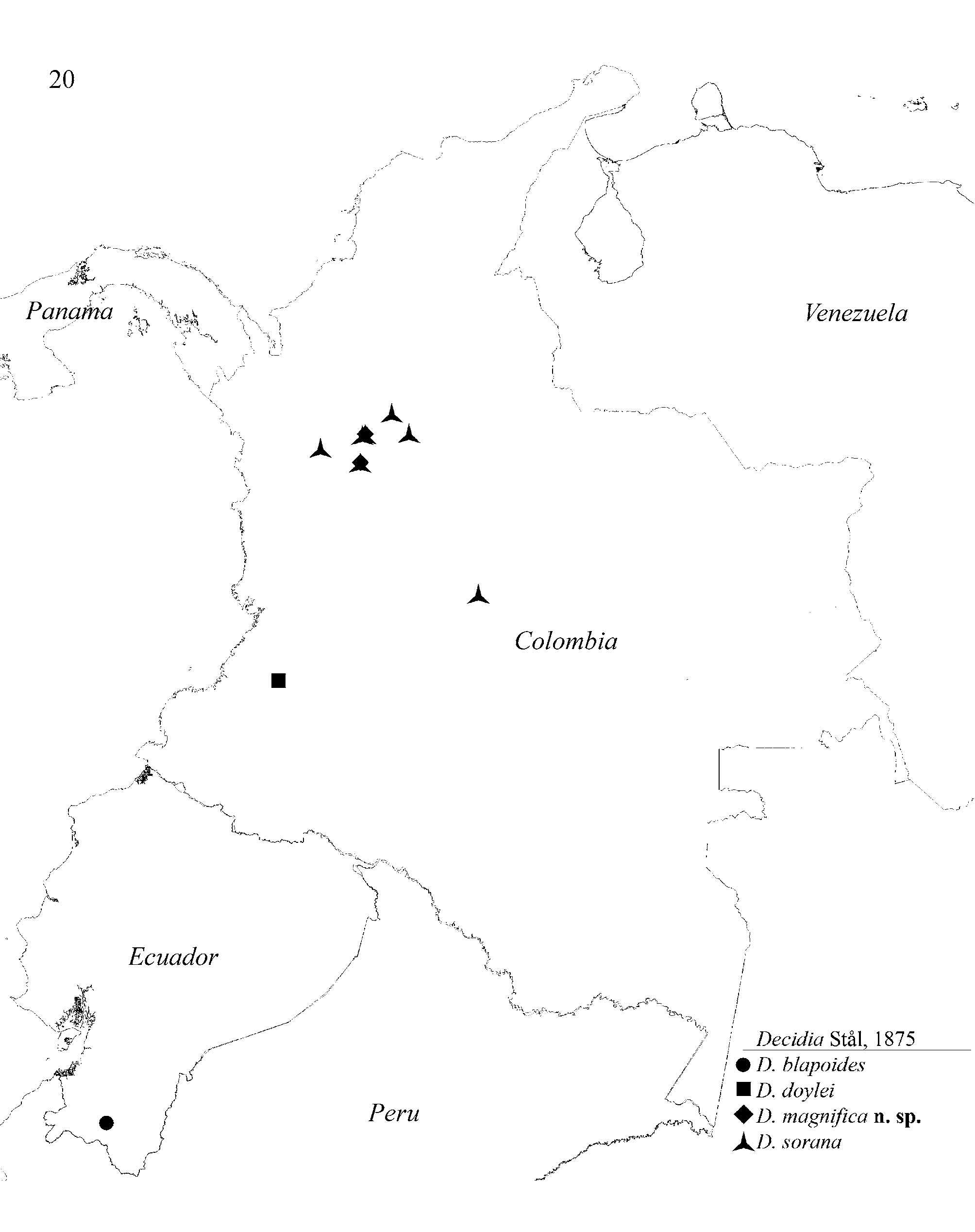
Distribution ( Fig. 20 View FIGURE 20 ): Central Cordilleras of Central and Southern Colombia and Ecuador between 2200 – 3000 m. According to Morrone (2006: 479, fig 2) the distribution of Decidia is restricted to the biogeographical province Cauca of the Northwestern South American Dominion.
Species included:
1. Decidia blapoides ( Redtenbacher, 1906: 95) [ Autolyca ]. n. comb. 2. Decidia doylei ( Caudell, 1906: 192) [ Autolyca ]. n. comb. 3. Decidia magnifica n. sp.
4. Decidia soranus ( Westwood, 1859: 127, pl. 17: 3 [ Phasma ].
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Decidia Stål, 1875
| Conle, Oskar V., Hennemann, Frank H., Ramírez-Mora, Manuel A. & Quiróz, John A. 2009 |
Autolyca
| Caudell 1906: 192 |
Phasma
| Westwood 1859: 127 |