
Aretopsis sandybrucei, Ashrafi & Ďuriš & Anker, 2021
|
publication ID |
https://doi.org/ 10.5852/ejt.2021.774.1527 |
|
publication LSID |
lsid:zoobank.org:pub:E7061310-45BA-4146-BE1B-84732A21BC54 |
|
DOI |
https://doi.org/10.5281/zenodo.5589767 |
|
persistent identifier |
https://treatment.plazi.org/id/2BC9890C-D171-47BD-96EB-4F737E65F59C |
|
taxon LSID |
lsid:zoobank.org:act:2BC9890C-D171-47BD-96EB-4F737E65F59C |
|
treatment provided by |
Felipe |
|
scientific name |
Aretopsis sandybrucei |
| status |
sp. nov. |
Aretopsis sandybrucei sp. nov.
urn:lsid:zoobank.org:act:2BC9890C-D171-47BD-96EB-4F737E65F59C
Figs 1 4 View Fig View Fig View Fig View Fig
Etymology
The new species is named after Dr. Alexander J. Bruce, or Sandy Bruce for his friends and colleagues, for his tremendous contribution to the taxonomy of caridean shrimps, including an important article on the genus Aretopsis ( Bruce 1969) .
Material examined
Holotype NEW CALEDONIA • ♂ (cl 5.31 mm); Coral Sea ; Chesterfield Plateau between New Caledonia and Queensland; 25°29ʹ S, 159°49′ E; depth 360–400 m; 4 Sep. 2017; MNHN exped.; KANADEEP 1 sta. DW4941; Capel Bank; MNHN-IU-2017-2924 . GoogleMaps
Paratypes NEW CALEDONIA • 1 ovigerous ♀ (cl 4.95 mm); same locality for holotype; 27°07ʹ S, 159°19ʹ E; depth 340–550 m; 19 Sep. 2017; MNHN exped.; KANADEEP 1 sta. DW5011; Nova Bank; GenBank COI gene: MZ389775 View Materials ; MNHN-IU-2017-3144 GoogleMaps • 1 ovigerous ♀ (cl 6.58 mm); same locality as holotype; 23°08ʹ S, 159°31ʹ E; depth: 280 m; 6 Sep. 2017; MNHN exped.; KANADEEP 1 stn. CP4958; Argo Bank; MNHN-IU-2017-2770 GoogleMaps .
Comparative material
Aretopsis amabilis De Man, 1910
AUSTRALIA • 1 ♂ (cl 6.7 mm), 1 ♀ (cl 6.8 mm); Queensland; Great Barrier Reef ; Lizard Island ; Coconut Beach ; 14.6842° S, 145.4700° E; depth 1 m; 25 Aug. 2010; I. Marin leg.; MTQ-W33075 GoogleMaps .
PHILIPPINES • 1 ♂ (cl 5.2 mm); Negros Island ; Bonbonon Point ; 9.0625 N, 123.1260 E; depth: 0–3m; 12 May 1978; Smithsonian exped.; stn. SP-5-2A, R/V Sting Ray V; USNM 213482 View Materials [examined by Anker (2001)] GoogleMaps .
Description
CARAPACE ( Fig. 1A, C View Fig ). Carapace glabrous, slightly swollen (more in females), poorly covering eyes dorsally and laterally, with deep cardiac notch. Rostrum ( Fig. 1A–B View Fig ) very short, triangular in dorsal view, slightly wider than long, compressed laterally, with short dorsal carina and feebly marked lateral lamella; tip rounded, distinctly overreaching distal half of first article of antennular peduncle, falling slightly short of its distal margin. Orbital teeth present in form of small, triangular, subacute infracorneal teeth ( Fig. 1A View Fig ), reaching to about one third of eye length in lateral view; extra-corneal and supra-corneal teeth absent. Anterolateral margin of carapace ( Fig. 1A View Fig ) between infra-corneal tooth and pterygostomial angle somewhat sinuous; pterygostomial angle broadly rounded.
PLEON ( Fig. 1C View Fig ). Pleura of first to fifth pleonites rounded antero- and postero-ventrally; second pleuron greatly expanded in females; sixth pleonite with distinct, fairly large articulated plate.
TELSON ( Fig. 1D–E View Fig ). Telson relatively broad, subrectangular in shape, gradually tapering distally; dorsal surface with anterior and posterior pairs of spiniform setae situated at about 0.5 and 0.7 of telson length, respectively; distolateral angle feebly projecting, with two pairs of spiniform setae, lateral about half as long as mesial; posterior margin between mesial pair of spiniform setae broadly convex.
ANTENNULE ( Fig. 1A–B View Fig ). Antennular peduncle short, stout; dorsally visible portion of first article wider than long; stylocerite well developed, distally blunt, reaching beyond half-length of second article to almost reaching its distal margin; second article very short, wider than long in dorsal view; third article slightly longer than second; lateral flagellum biramous, with fused portion consisting of six or seven units and short accessory ramus consisting of three feebly distinct units; groups of aesthetascs extending from third unit of fused portion to end of accessory ramus.
ANTENNA ( Fig. 1A–B View Fig ). All antennal articles robust; basicerite moderately developed, with subacute distoventral tooth; scaphocerite broad, ovate, with strong distolateral tooth reaching far beyond distally rounded blade, not or only slightly overreaching antennular peduncle, lateral margin straight; carpocerite relatively stout, reaching slightly beyond distolateral tooth of scaphocerite and to end of third article of antennular peduncle; flagellum moderately stout.
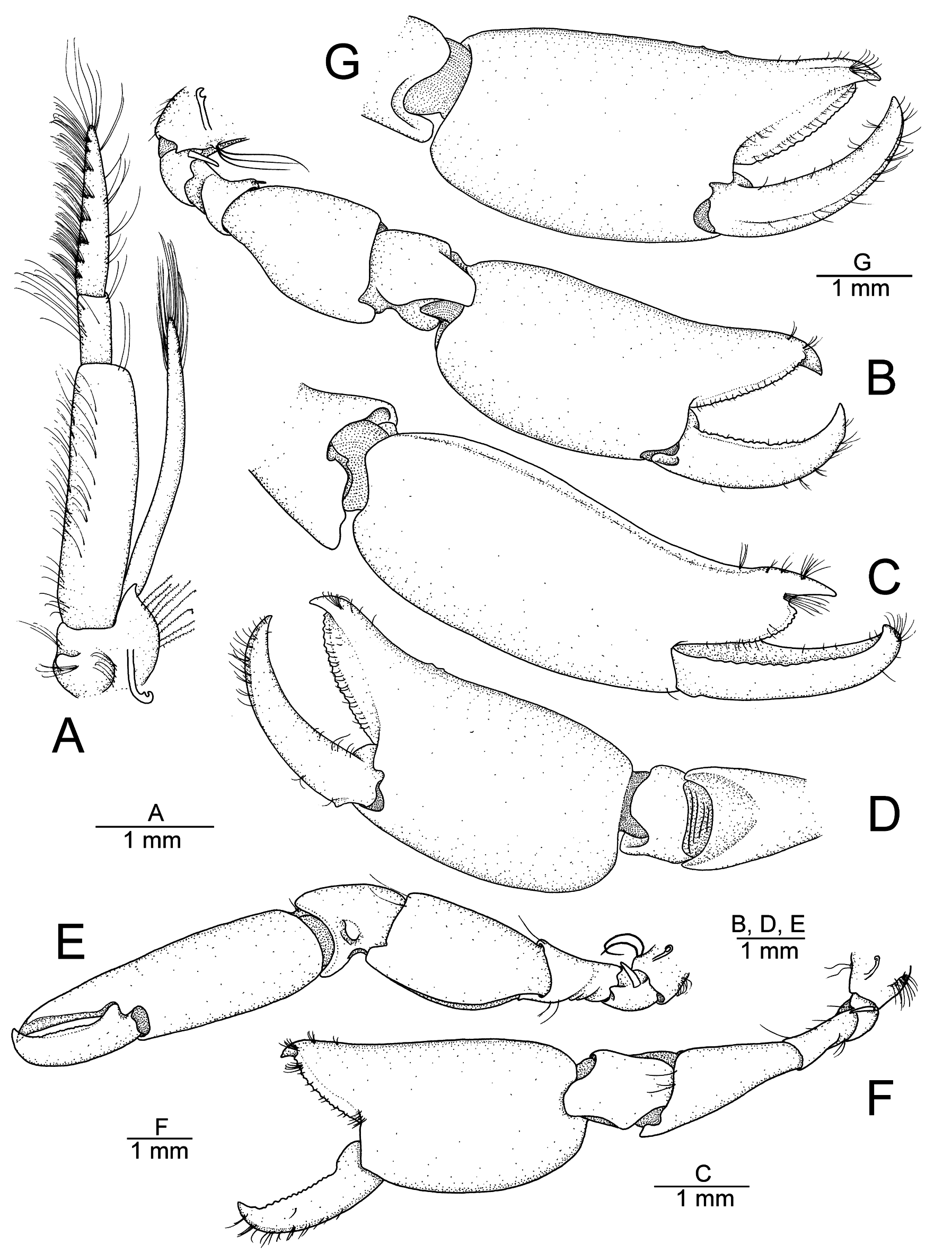
MOUTHPARTS. Mandible with well-developed palp. Third maxilliped ( Fig. 2A View Fig ), with coxa stout, bearing strap-like epipod, lateral plate produced anteriorly with tip pointing slightly laterally; antepenultimate article robust, about four times as long as wide, with blunt distodorsal margin; penultimate article short, noticeably more slender than antepenultimate article, about 2.3 times as long as wide and 0.3 times as long as antepenultimate article; ultimate article slender, tapering distally, about 2.5 times as long as penultimate article, with rows of numerous serrulate setae ventromesially and some long setae at tip, latter without spiniform setae; exopod well developed, reaching to middle of penultimate article.
FIRST PEREIOPODS (= CHELIPEDS) ( Fig. 2B–G View Fig ). Chelipeds unequal in size, subsymmetrical in shape, carried extended with dactylus in ventral to ventrolateral position. Male major cheliped ( Fig. 2B–D View Fig ) (on the right side in the holotype) larger and more voluminous than minor cheliped by about 20% in male. Coxa with strap-like epipod and set of setobranchs. Basis short, stout, with small exopod-like lobe. Ischium slightly longer than basis, widening distally, somewhat cup-shaped, armed with one spiniform seta on distodorsal margin. Merus robust, swollen, somewhat vase-shaped, about 1.5 times as long as maximal width, distoventral margin with subrectangular projection. Carpus robust, about 0.7 times as long as merus and complex in lateral view, with broad prominence, ventral surface smaller, much wider than long. Palm compressed laterally, ovate in general shape, about 1.7 times as long as high, with two small tubercles on dorsomesial surface, near base of pollex; blunt ridge extending along dorsolateral margin of palm onto pollex. Fingers subequal in length, about 0.8 times as long as palm, crossing distally when closed with strongly curved fingertips; pollex with row of low irregular teeth along entire cutting edge; dactylus with small irregular teeth on proximal two thirds, distal third unarmed. Male minor cheliped ( Fig. 2E–G View Fig ) (on the left side in the holotype) generally similar to major cheliped, except for some proportions. Coxa and basis as in major cheliped, smaller in size. Ischium more slender, about three times as long as wide. Merus less robust and less swollen compared to that of major cheliped, vase- shaped, about twice as long as maximally wide. Carpus about cup-shaped, with small bump on lateral surface. Chela generally similar to that of major cheliped. Palm 1.5 times as long as high, bearing three small tubercles on dorsomesial surface, near base of pollex. Fingers about 0.7 times as long as palm; both pollex and dactylus armed with small teeth along their entire cutting edges. Female major cheliped overall similar to that of male (see Fig. 4 View Fig ); female minor cheliped unknown (missing in both paratypes).
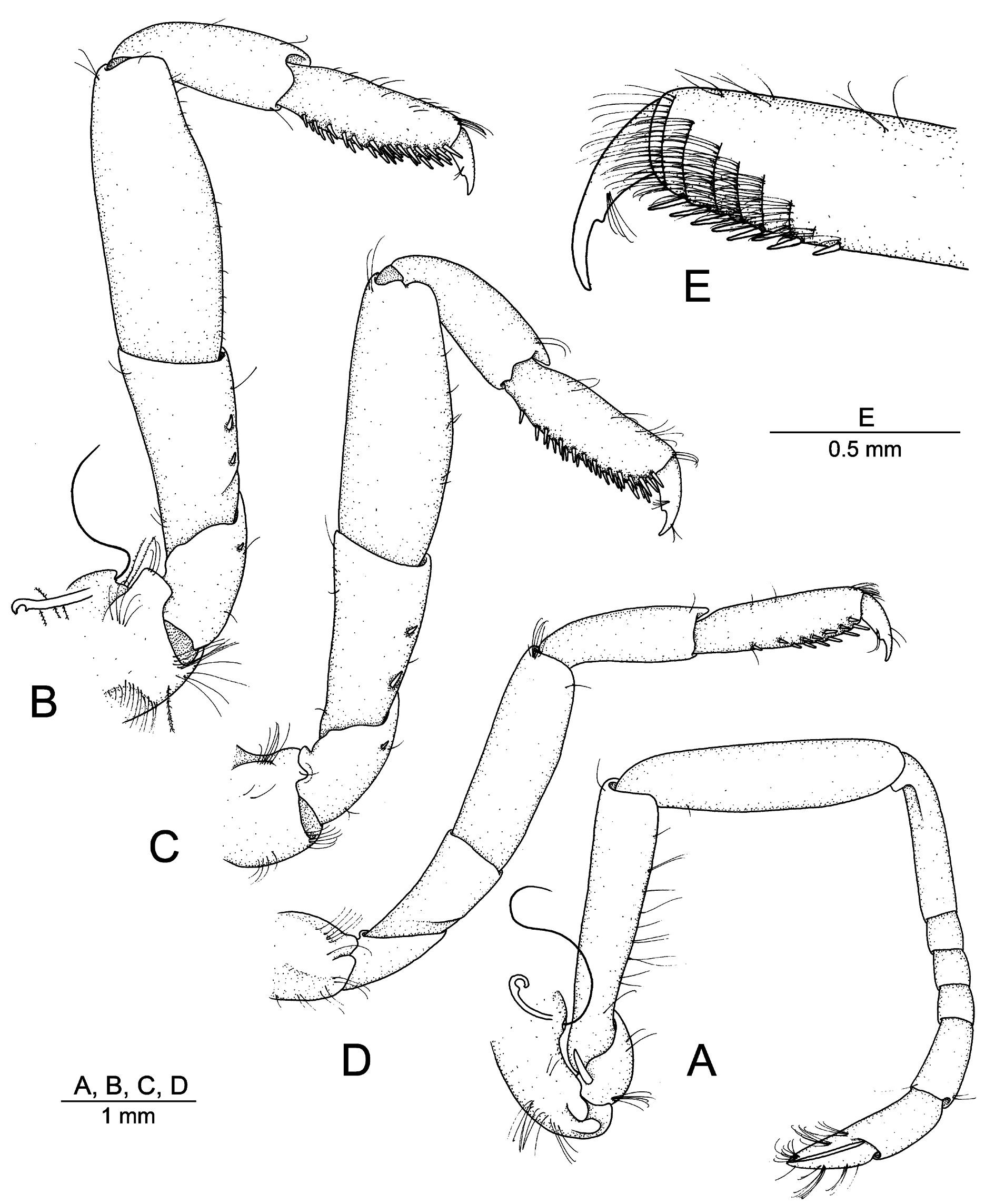
SECOND TO FIFTH PEREIOPODS. Second pereiopod ( Fig. 3A View Fig ) relatively short; coxa with strap-like epipod and single setobranch; basis short, with small exopod-like lobe; ischium and merus subequal in length, latter about 4.5 times as long as maximally wide, ventral margin slightly convex; carpus about 1.2 times as long as merus, with five subdivisions, proximal longest, slightly shorter than length sum of remaining four, palm slightly longer than fingers. All walking legs (third to fifth pereiopods) very stout. Third pereiopod ( Fig. 3B View Fig ) with coxa bearing strap-like epipod and single setobranch; basis distally with one minute spiniform seta on ventrolateral surface; ischium stout, distally widening, with two small spiniform setae on ventrolateral surface; merus robust, about 2.5 times as long as maximal width, about 1.5 times as long as ischium, with distolateral margin forming blunt angle; carpus noticeably more slender than merus, about 0.6 length of merus, 2.7 times as long as wide; propodus slightly longer than carpus, armed with about 25 stout spiniform setae on ventral margin, most of them organised in two rows, distal-most spiniform setae longer and more robust; dactylus stout, slightly less than 0.3 length of propodus, flexor (= ventral) margin with small tooth-like projection. Fourth pereiopod ( Fig. 3C View Fig ) generally similar to third pereiopod, slightly less robust; coxa lacking both strap-like epipod and setobranch. Fifth pereiopod ( Fig. 3D–E View Fig ) noticeably less stout than third and fourth pereiopods; coxa lacking setobranch; basis and ischium unarmed; merus about 3.2 times as long as maximal width; carpus 0.7 times as long as merus; propodus with a dozen of spiniform setae on distal third, increasing in size and density distally and organised in two more or less distinct rows; distomesial surface of propodus with about seven transverse rows of serrulate setae together forming well-developed cleaning brush; dactylus almost 0.4 length of propodus, similar to that of third and fourth pereiopods.
PLEOPODS AND UROPOD. Second pleopod with appendix masculina greatly overreaching small appendix interna. Female second pleopod without appendix masculina, appendix interna very long (also on following pleopods). Uropod ( Fig. 1D–E View Fig ) reaching well beyond posterior margin of telson; protopod distally with two small blunt projections separated by shallow furrow; exopod relatively broad, ovate, its distolateral margin with small blunt tooth and slender spiniform seta, latter not reaching posterior margin of exopod; diaeresis broadly sinuous, lacking lateral tooth; endopod ovoid, without specific features.
COLOUR PATTERN. Most of carapace and pleon translucent with pale red-wine colour, however, most of it apparently due to the partial visibility of internal organs (possibly ovary); on the other hand, clusters of small red chromatophores clearly visible on anterior flanks of carapace and pleura; mid-dorsal part of carapace and pleon with poorly defined, whitish or colourless longitudinal band stretching from rostrum to (and including) telson; short, whitish streaks and reticulations present on carapace flanks and pleon; all larger appendages, i.e., antennules, antennae, third maxillipeds, all pereiopods and uropods, generally translucent with pale tan-yellow tinge, parts of chelipeds with more intense yellowish colour and hyaline-white distal part of fingers ( Fig. 4 View Fig ).
Distribution
Presently known only from three deep-water banks (Capel, Nova and Argo) of the Chesterfield Plateau, Coral Sea (part of New Caledonia's Exclusive Economic Zone), between New Caledonia and Queensland, Australia.
Ecology
The three specimens of A. sandybrucei sp. nov. were collected by a Warén dredge (DW stations) or a bottom trawl (CP stations) over deep reef bottoms, at a depth range of 280– 550 m. No associations were noted in the field. However, several animals from Argo Bank (stn. CP4958) were photographed immediately after pulling up and emptying the trawl into a bin, among them a female paratype of A. sandybrucei sp. nov. ( Fig. 4 View Fig ) and at least two species of deep-water hermit crabs of the family Paguridae ( Fig. 5 View Fig ), including possibly a species of Diacanthurus McLaughlin & Forest, 1997 (T. Komai, pers. comm. to AA).
Remarks
Aretopsis sandybrucei sp. nov. can easily be distinguished from its two congeners, A. amabilis and A. manazuruensis , by the proportions of the articles of the three last pereiopods (walking legs) and the development of the secondary (= accessory) unguis on the flexor margin of their dactyli. In A. sandybrucei sp. nov., the third, fourth and fifth pereiopods are generally much stouter than their homologues in A. amabilis and A. manazuruensis . This stoutness can easily be observed and measured on the merus of the third pereiopod, which is noticeably swollen and is at most 2.7 times as long as maximal width of A. sandybrucei sp. nov. ( Fig. 3B View Fig ) vs not swollen and at least 4.0 times as long as maximal width in A. amabilis (cf. De Man 1911: pl. 4 fig. 14f; Banner & Banner 1973: fig. 12k; confirmed on comparative material from Lizard Island, MTQ-W33075), and slightly swollen and about 3.2 times as long as maximal width in A. manazuruensis (cf. Suzuki 1971: fig. 12.4). Secondly, in A. sandybrucei sp. nov., the flexor margin of the dactylus of the third to fifth pereiopods bears a very small, i.e., only slightly projecting, secondary unguis ( Fig. 3A View Fig ), which contrasts to a much better developed, i.e., clearly more projecting, secondary unguis on the dactylus of these appendages in the other two species (cf. Bruce 1969: fig. 2c; Suzuki 1971: fig. 12.6a; Banner & Banner 1973: fig. 12k). In addition, in A. sandybrucei sp. nov., the basis of the third and fourth pereiopods is armed with a minute spiniform seta, which does not seem to be the case of A. amabilis based on De Man’s (1910, 1911) description and illustrations (verified on comparative material, MTQ-W33075), and is also absent in A. manazuruensis according to Suzuki (1971: figs 12.4, 5). Another small difference between A. sandybrucei sp. nov., A. manazuruensis and A. amabilis lies in the antepenultimate article of the third maxilliped, in which the distodorsal margin is ending bluntly in the former two species ( Fig. 2A View Fig ; Suzuki 1971: fig. 11.3), but has a distinct rounded projection in the latter species ( Bruce 1969: fig. 3f).
A direct comparison of the chelipeds between the species of Aretopsis is rather difficult because of the variation in the shape and armature of the major chela reported for A. amabilis ( Bruce 1969; Nomura 1986) and the very limited material presently available for A. manazuruensis , known only from the female holotype ( Suzuki 1971), and A. sandybrucei sp. nov., known from the male holotype and two female paratypes. However, none of the specimens of the new species has the fingers largely gaping and armed with prominent teeth, as illustrated for several adult specimens of both sexes of A. amabilis (cf. Ramadan 1936: pl. 2, fig. 12, as A. aegyptiaca ; Bruce 1969: figs 14a–b, e–f; Banner & Banner 1973: fig. 12c; Nomura 1986: fig. 3a–c; Marin 2010: figs 1, 4). Both A. amabilis and A. sandybrucei sp. nov. can be separated from A. manazuruensis by the very unequal and asymmetrical chelipeds, which were described by Suzuki (1971) as “symmetrical in size”, with only the left cheliped illustrated in the habitus drawing (idem: fig. 10), in addition to two figures showing the proximal articles of the right cheliped (idem: fig. 12.2) and the chela of what appears to be the left cheliped (contrary to Suzuki’s figure legend), judging from the position of a strong carpal process present on the lateral surface only (idem: fig. 12.3).
The cheliped merus of A. sandybrucei sp. nov. is ventrally unarmed and distoventrally bears a rectangular projection ( Fig. 2B, D View Fig ), whereas the cheliped merus of at least some specimens of A. amabilis is ventrally armed with spiniform setae and distoventrally bears a prominent tooth ( De Man 1911: pl. 4, fig. 14d; Marin 2010: fig. 2d; comparative material, MTQ-W33075). Bruce (1969: fig. 4) did not illustrate these spiniform setae, but in the text mentioned that “the merus is provided with one or two long slender spines”. Miyake & Miya (1967) stated that the “outer lower margin of the merus is indented and bristled”, possibly referring to the spiniform setae (unclear in their fig. 3d). On the other hand, they were neither illustrated nor described for the material of A. amabilis reported by Ramadan (1936) or Banner & Banner (1973), and therefore, it is not quite clear whether they are present in all specimens so far reported as A. amabilis .
Another cheliped feature that may add to the distinction between A.sandybrucei sp.nov., A. manazuruensis and A. amabilis is the presence (in the first two species) or absence (in the last species) of a slender spiniform seta on the distodorsal margin of the ischium (cf. Fig. 2A View Fig ; Bruce 1969: fig. 4a; Suzuki 1971: fig. 12.2; Banner & Banner 1973: fig. 12d–e). As for the spiniform setae on the merus, the absence of this seta in A. amabilis may be simply due to the lack of detailed observation of the base of the cheliped and/or inaccurate figures (or with all setae simply omitted for clarity), although it was confirmed to be absent in both specimens from Lizard Island.
The colour pattern of A. sandybrucei sp. nov. ( Fig. 4 View Fig ) is generally similar to that reported for A. amabilis ( Vannini et al. 1993: fig. 1 (bottom); Marin 2010: figs 3–4) and A. manazuruensis ( Suzuki 1971: fig. 10). However, the new species is clearly different from A. amabilis and A. manazuruensi s in having a generally less contrasting, somewhat duller colour pattern, including the more translucent and homogenously coloured pale yellow chelae (although with distally white fingers) and tail fan.
Aretopsis sandybrucei sp. nov. is also ecologically separated from its shallow-water congeners, presently being known from a depth range of 280–550 m, i.e., 230 m below the deepest record of A. amabilis , represented by a specimen dredged at 50 m off Ujung Pandang, Sulawesi, Indonesia ( Banner & Banner 1985). However, most records of A. amabilis and the single record of A. manazuruensis , are from the lower intertidal or shallow subtidal depths ranging from 1 to 10 m, often on coral reefs and adjacent habitats ( De Man 1910; Miyake & Miya 1967; Bruce 1969; Banner & Banner 1973; Marin 2010, 2014; present study). The host (or hosts, if multiple) of A. sandybrucei sp. nov. are currently unknown, although the general similarity of the new species to other species of the genus, including the colour pattern, suggests that it may be a deep-water hermit crab of the family Paguridae , possibly Diacanthurus sp. (see above and Fig. 5 View Fig ). Noteworthy, this pagurid carried a shell almost fully occupied by a large sea anemone ( Fig. 5B–C View Fig ), which is also commonly observed in many shallow-water hosts ( Dardanus and Aniculus ) of A. amabilis .



| MNHN |
Museum National d'Histoire Naturelle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |