
Poecilonotini Jakobson, 1913
|
publication ID |
https://doi.org/ 10.5281/zenodo.5324888 |
|
persistent identifier |
https://treatment.plazi.org/id/BE1887AB-9A12-2A05-FE8C-FB3FFDA9F9FE |
|
treatment provided by |
Felipe |
|
scientific name |
Poecilonotini Jakobson, 1913 |
| status |
|
Tribe Poecilonotini Jakobson, 1913 sensu novo
Poecilonotina Jakobson, 1913: 786 (as ‘fam. Buprestini : tribus Poecilonotina’; list; characters in key). Type genus: Poecilonota Eschscholtz, 1829 View in CoL .
Poecilonotina : OBENBERGER (1930): 310 (catalogue; in synonymy of Buprestini : Dicercites).
Poecilonotini (sensu novo): ALEXEEV & BEBKA (1970a): 6; ALEXEEV & BEBKA (1970b): 158 (both:comment; characters; in ‘Ancylocheirinae’ (now Buprestinae ).
Poecilonotini (sensu novo): VOLKOVITSH (2001) : 55 (catalogue of higher taxa; in Chalcophorinae (now Chrysochroinae )), 84, 96 (comment; characters), 108 (summary); BÍLÝ & VOLKOVITSH (2003) : 108 (catalogue of higher taxa; in Chalcophorinae (in present Chrysochroinae )); BELLAMY (2003): 40 (catalogue of higher taxa; in Chrysochroinae ); NELSON et al. (2008): 61 (catalogue; in Chrysochroinae ); BELLAMY (2008): 594 (catalogue; in Chrysochroinae ).
History of classification. The name ‘Poecilonotina’ was first used by JAKOBSON (1913: 773, 786), who used the name ‘Poecilonotina’ both for the category of tribe (sensu lato) and for the category of subtribe (sensu stricto). He replaced by ‘Poecilonotina’ sensu lato and sensu stricto the formerly used names ‘Buprestini’ and ‘Dicercites’, respectively, of the previously widely used system by KERREMANS (1893); the reason for this change is not clear. JAKOBSON (1913: 786) included three subtribes into ‘Poecilonotina’ (sensu lato), ‘ Poecilonotina = Dicercina’ (sensu stricto), ‘Ancylochirina [nov.] = Buprestina’ and ‘Anthaxiina’, and the following genera into the subtribe Poecilonotina (sensu stricto): Cardiaspis Saunders, 1866 View in CoL , Dicerca Eschscholtz, 1829 View in CoL , Poecilonota Eschscholtz, 1829 View in CoL and Lampra Dejean, 1833 (now Lamprodila Motschulsky, 1860 View in CoL ). JAKOBSON (1913) initially used only the form of antennal pores to define the Poecilonotini (sensu stricto); RICHTER (1952) mentioned difference between wing venation (position of R-M vein) in Poecilonota View in CoL and Lampra (now Lamprodila View in CoL ) versus other Dicercini and also differences between the larvae of Lampra (now Lamprodila View in CoL ) and Poecilonota View in CoL versus Dicerca View in CoL .
The tribe Poecilonotini with the genera Poecilonota and Lampra (now Lamprodila ) was independently created also by ALEXEEV & BEBKA (1970a,b), who separated the Poecilonotini in the subfamily Ancylocheirinae Jakobson, 1913 (now Buprestinae ) on the basis of seven visible abdominal tergites, contrary to Dicerca and other genera in the subfamily Buprestinae sensu ALEXEEV & BEBKA (1970a ,b) (now Chrysochroinae ) with eight visible tergites.
COBOS (1986) newly defined two subfamilies: Chalcophorinae Lacordaire, 1857 (now Chrysochroinae ) and Buprestinae and included the genera Dicerca , Poecilonota and Ovalisia Kerremans, 1900 (= Lampra , now subgenus of Lamprodila ) into the tribe Dicercini of the subfamily Buprestinae (he did not mention the name Poecilonotini ).
HOŁYŃSKI (1993) defined the family Buprestidae in quite a new and revolutionary way. In his concept, the tribe Buprestini includes 18 subtribes (seven of them new and also, e.g., the subtribes Chrysochroina , Chalcophorina , Buprestina and Psilopterina ). Dicerca , Poecilonota and Ovalisia (sensu lato, now Lamprodila ) and further 11 genera were placed in the subtribe Psilopterina Lacordaire, 1857 . One of his new subtribes was the subtribe Haplotrinchina Hołyński, 1993 , in which the genera Cardiaspis (erroneously as ‘ Bellamyola nom. nov. ’, see VOLKOVITSH (1997)), Haplotrinchus Kerremans, 1903 and Melobasina Kerremans, 1900 were included. In a short comment he wrote: ‘The Haplotrinchina str. n. make a very well-defined group, though superficially Bellamyola n.n. [= Cardiaspis ...] looks quite dissimilar from Melobasina KERR. [including Nesotrinchus OBB. , which, in my view, cannot be separated even as a subgenus, “ N. ” coeruleipennis (THS.) being much more closely related to M. fossicollis KERR. than to “ N. ” australicus (KERR.) ]’. BELLAMY (1997: 76, resp. 2002: 62) interpreted this comment as a synonymisation of the genera Melobasina and Nesotrinchus Obenberger, 1924 but this interpretation is wrong.
From the comments by HOŁYŃSKI (1993), we can judge that he based his study of the genus Melobasina on M. fossicollis ( Kerremans, 1906) but without any specification of studied specimens. He probably saw specimens from BMNH, identified by B. Levey as ‘ fossicollis ’ but in fact belonging to M. apicalis Kerremans, 1900 (see below under M. apicalis ). Characters used for the subtribe Haplotrinchina in his key (couplets u(b), x(cc) and cc(z)) do not explicitly define either Melobasina or Nesotrinchus . The definition of the subtribe Haplotrinchina by HOŁYŃSKI (1993) is quite insufficient and the reasons for the treatement of genera Melobasina and Nesotrinchus in Haplotrinchina is so vague that it is impossible to validate them.
BÍLÝ (1999) mentioned differences between the larvae of Dicerca and Capnodis Eschscholtz, 1829 and the larvae of Lampra (now Lamprodila ) and Poecilonota . BÍLÝ & VOLKOVITSH (2002: Table 2) published a set of larval characters differentiating Lampra (now Lamprodila ) and Poecilonota from the larvae of Dicerca , Dicercomorpha Deyrolle, 1864 , Haplotrinchus and Polybothris Dupont, 1833 [sic!].
JENDEK (2001) found differences in the general structure of the abdomen between the Dicercini on one hand and Lampra (now Lamprodila ) and Poecilonota and Kisanthobia Marseul, 1865 on the other.
VOLKOVITSH (2001) principially disagreed with the concept of HOŁYŃSKI (1993) and presented results of a detailed study of the antennal structures in the Buprestidae View in CoL (without the Agrilinae ) based on taxa of generic rank. The arrangement and status of suprageneric taxa reflect similarities and presumed evolutionary trends in antennal structures ( VOLKOVITSH 2001: 52 –59); the list used several informal categories: ‘complex’, ‘lineage’, ‘branch’ and ‘generic group’. The term ‘generic group’ was used at three different levels: tribal, subtribal and lower than the subtribal level.
Unlike HOŁYŃSKI (1993), VOLKOVITSH (2001) re-introduced again the subfamily Chalcophorinae (now Chrysochroinae ) but in a rather different concept than did COBOS (1986). Contrary to COBOS (1986) (see above), he placed the genera Dicerca View in CoL , Poecilonota View in CoL and Palmar (now a subgenus of Lamprodila View in CoL ) into the Chalcophorinae (now Chrysochroinae ) and divided this subfamily into two ‘lineages’, Chalcophorioid (now Chrysochroioid) and Psilopterioid (now Dicercioid), and placed the tribe Poecilonotini in the former lineage. The tribe Psilopterini (now Dicercini ) was placed together with the tribe Sphenopterini Lacordaire, 1857 in the latter lineage and divided into four subtribes, including the subtribe Haplotrinchina with the genera Cardiaspis View in CoL and Haplotrinchus View in CoL . The definition of the tribes Poecilonotini and Dicercini (as Psilopterini ) was based mainly on the anennal structures (U2 sensillae absent in Poecilonotini but present in Dicercini ) but also other characters such as morphological differences between the larvae of both tribes (see also BÍLÝ & VOLKOVITSH 2002 ) or a serrate elytral margin in the Poecilonotini ( VOLKOVITSH 2001: 69 , 84, 96). Both VOLKOVITSH (2001) and BÍLÝ & VOLKOVITSH (2003) erroneously attributed the authorship of the tribe Poecilonotini to ALEXEEV & BEBKA (1970a,b).
VOLKOVITSH (2001) studied the antennal structures of the following taxa of the Poecilonotini and Haplotrinchina : Palmar subgen. Palmar Schaefer, 1949 (now in Lamprodila View in CoL ) (one species), Palmar subgen. Scintillatrix Obenberger, 1956 (now Lamprodila View in CoL ) (two species), Poecilonota View in CoL (one species), Nesotrinchus View in CoL (one species), Cardiaspis View in CoL (one species) and Haplotrinchus View in CoL (one species); Pseudhyperantha Saunders, 1869 View in CoL and Melobasina View in CoL were not studied. As a result of this comparison, VOLKOVITSH (2001: 55, 68, 96) confirmed Nesotrinchus View in CoL as a valid genus of the tribe Poecilonotini and created for it an independent, informal ‘ Nesotrinchus View in CoL generic group’.
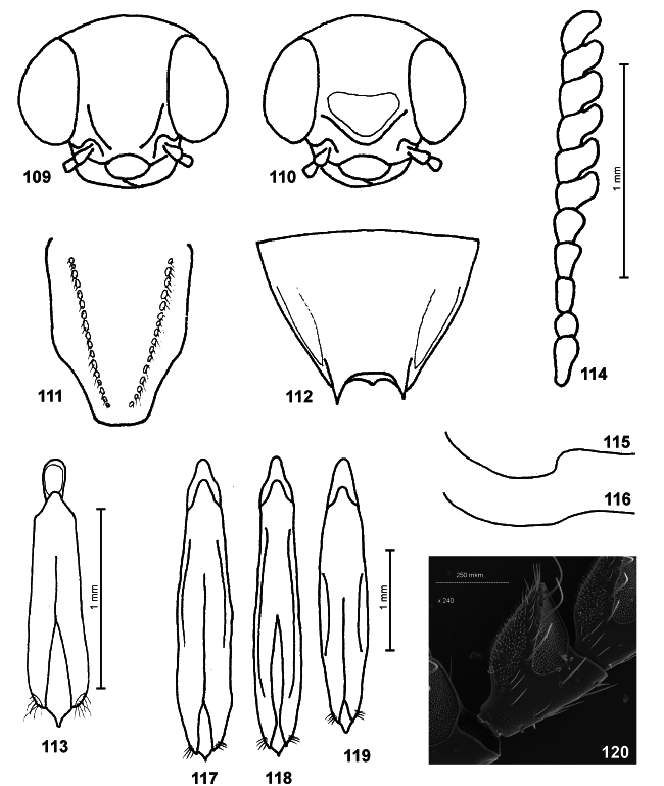
BELLAMY (2003, 2008) accepted the results of VOLKOVITSH (2001) but treated Nesotrinchus View in CoL as a junior subjective synonym of the genus Melobasina View in CoL (misinterpretation, see above). He also included the genus Pseudhyperantha View in CoL ( Figs. 1–6 View Figs , 120 View Figs ) into the subtribe Haplotrinchina (in Chrysochroinae : Dicercini ), a traditional concept introduced by KERREMANS (1893). On the contrary he did not accept the opinion of TOYAMA (1989), who transferred the genus Pseudhyperantha View in CoL into the tribe Buprestini (in Buprestinae ) near to the genus Buprestis Linnaeus, 1758 View in CoL . Finally, HOŁYŃSKI (2008) transferred Pseudhyperantha View in CoL back to the Buprestinae but into the tribe Stigmoderini .
Placement of the genus Pseudhyperantha . Antennal structures of Pseudhyperantha correspond with those of the Buprestini ( Fig. 120 View Figs ): antennomeres triangular, bilaterally flattened, apical pits large, irregular, situated somewhat ventrally. Lateral sensory fields large, formed by B4c sensillae, U2 sensillae missing (see VOLKOVITSH 2001: 71 , Figs. 128–130 View Figs ). The antennal structured of the Stigmoderini were treated in detail by GARDNER (1989: 303, Figs. 36–41 View Figs ) and VOLKOVITSH (2001: 72, Figs. 144–149). The prosternal process is of buprestine type in Pseudhyperantha ( Fig. 5 View Figs ), with preapical lobes missing (see also KOLIBÁČ 2000, Fig. 11 View Figs ); the prosternal processes of Stigmoderini was discussed in GARDNER (1989: 305, Figs. 61–62 View Figs ). Wing venation of Pseudhyperantha (see TOYAMA 1989: 194, Fig. 1 View Figs ) corresponds to Buprestis (see KOLIBÁČ 2000: 152, Fig. 138; FEDORENKO 2009: 260, Fig. 57 View Figs ) rather than a stigmoderine type (see GARDNER 1989: 311, Fig. 66 View Figs ): radial cell (rc) long, vein Rr distinct, Mr stretching far beyond intersection with ‘r-m’, ‘wedge’ cell (2a) closed; for the nomenclature of veins ( Fig. 121 View Figs ) see VOLKOVITSH (2008, Fig. 54 View Figs ) and FEDORENKO (2009: 260, Figs. A103–A106 View Figs ).
Moreover, the ovipositor of Pseudhyperantha ( Fig. 2 View Figs ) possesses all characters of Buprestini : it is simple and tubular with simple styli and without a dorsal valve (more complex structures and a dorsal valve are the synapomorphies of the Stigmoderini (see GARDNER 1989)). The placement of Pseudhyperantha into the tribe Stigmoderini by HOŁYŃSKI (2008) thus cannot be maintained. We accept the opinion of TOYAMA (1989) who attributed this genus to the tribe Buprestini . The subtribal position and closest affinities of Pseudhyperantha within the Buprestini remain unclear. We suppose it should be separated as distinct monogeneric subtribe but this assumption requires further investigations.
The genera Melobasina and Nesotrinchus . We have studied seven specimens of Melobasina (four species) and 29 specimens of Nesotrinchus (three species). Species of Nesotrinchus are very similar to each other; they are undoubtedly congeneric and distributed only on the Pacific islands of Fiji, Tonga, Niue and Wallis. Species of Melobasina are also very similar to each other. They are distributed on the Solomon and Maluku Islands and differ from Nesotrinchus by the colouration and body shape (see Table 1 and Figs. 59–97 View Figs View Figs View Figs ). The frons of Melobasina species is regularly convex and much wider than that of Nesotrinchus , and the latter genus also possesses well-developed supraantennal carinae and deep frontal depression.
New concept of the tribe Poecilonotini . Having studied representatives of the genera mentioned above (see also the list in Material and methods) and further representatives of the genus Lamprodila (subgenera Cinyrisia Hołyński, 2000 , Erialata Zykov, 1998 and Mabomisia Hołyński, 2000 ), we define newly the tribe Poecilonotini and the subtribe Poecilonotina and suggest a new concept of the tribe Poecilonotini (sensu novo) containing subtribes Poecilonotina and Nesotrinchina subtrib. nov.






No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
Poecilonotini Jakobson, 1913
| Bílý, Svatopluk, Kubáň, Vítězslav & Volkovitsh, Mark G. 2009 |
Poecilonotini
| NELSON G. H. & WALTERS G. C. Jr. & HAINES R. D. & BELLAMY C. L. 2008: 61 |
| BELLAMY C. L. 2008: 594 |
| BILY S. & VOLKOVITSH M. G. 2003: 108 |
| BELLAMY C. L. 2003: 40 |
| VOLKOVITSH M. G. 2001: 55 |
Poecilonotini
| ALEXEEV A. V. & BEBKA M. P. 1970: 6 |
| ALEXEEV A. V. & BEBKA M. P. 1970: 158 |
Poecilonotina
| OBENBERGER J. 1930: 310 |
Poecilonotina
| JAKOBSON G. G. 1913: 786 |