
Hyloscirtus japreria, Rojas-Runjaic & Infante-Rivero & Salerno & Meza-Joya, 2018
|
publication ID |
https://doi.org/ 10.11646/zootaxa.4382.1.4 |
|
publication LSID |
lsid:zoobank.org:pub:B2888C18-AE2D-40E2-BFEC-BCB929790317 |
|
DOI |
https://doi.org/10.5281/zenodo.5964502 |
|
persistent identifier |
https://treatment.plazi.org/id/BF23165A-FFDE-FFBB-07D0-FE24D0AEF9BB |
|
treatment provided by |
Plazi |
|
scientific name |
Hyloscirtus japreria |
| status |
sp. nov. |
Hyloscirtus japreria View in CoL sp. nov.
( Figs. 2–3 View FIGURE 2 View FIGURE 3 , 5–6 View FIGURE 5 View FIGURE 6 , 10 View FIGURE 10 )
Suggested common name in English: Perijá’s Stream Frog Suggested common name in Spanish: Rana Torrentícola de Perijá
Hyloscirtus View in CoL sp.— Rojas-Runjaic et al. (2016: 36)
Holotype. Adult male, MHNLS 19236 View Materials (field number SP 294; Figs. 2–3 View FIGURE 2 View FIGURE 3 ), from Guacharaca Camp , Tetari Kopejoacha creek , Rio Negro basin, Sierra de Perijá, Machiques de Perijá municipality, Zulia state, Venezuela (10°04’21.9 N – 72°51’16.7 W; elevation 1,661 m asl), collected on 27 May 2009, by Fernando J. M. Rojas-Runjaic and Edwin E. Infante-Rivero. GoogleMaps
Paratypes. 21 specimens (16 males and five females): Two adult males, MHNLS 18837–18838 View Materials (field numbers SP 44–45) from the creek behind the house at Cerro Las Antenas , Sierra de Perijá, Rosario de Perijá municipality , Zulia state, Venezuela (10°20’37.0”N – 72°33’41.0”W; 1,430 m asl), collected on 27 March 2008, by Fernando J. M. Rojas-Runjaic; two adult males, MHNLS 18867 View Materials , 18869 View Materials (field numbers SP 74, 76) and one adult female, MHNLS 18868 View Materials (field number SP 75), from the same locality, collected on 28 March 2008, by Fernando J. M. Rojas-Runjaic, Edwin E. Infante-Rivero, Pablo Velozo and Paul Granado; two adult males, MHNLS 18971–18972 View Materials (field numbers SP 117–118), from the same locality, collected on 10 July 2008, by Fernando J. M. Rojas-Runjaic, Pedro Cabello, Kripsy Herrera and Arlene Cardozo. One male, MHNLS 22568 View Materials (field number SP 218), from the same locality, collected on 26 April 2009, by Fernando J. M. Rojas-Runjaic, Adriana Becerra and Arnaldo Ferrer. Two adult males, MHNLS 18888–18889 View Materials (field numbers SP 95–96) from the creek near to second antenna, Cerro Las Antenas, Sierra de Perijá , Rosario de Perijá municipality GoogleMaps , Zulia state, Venezuela (10°19’40.0”N – 72°35’27.0”W; 1,832 m asl), collected on 29 March 2008, by Fernando J. M. Rojas-Runjaic and Pedro Cabello; one adult male, MHNLS 18988 View Materials (field number SP 134), from the same locality, collected on 12 July 2008 by Fernando J. M. Rojas- Runjaic and Pedro Cabello. Two adult females, MHNLS 19166–19167 View Materials (field numbers SP 224–225), and one adult male, MHNLS 19168 View Materials (field number SP 226) with the same locality and collectors as the holotype, collected on 22 May 2009 GoogleMaps ; two males, MHNLS 19234 View Materials , 19237 View Materials (field numbers SP 292, 295), and one female, MHNLS 19235 View Materials (field number SP 293) with the same locality, date and collectors as the holotype. Three males, UIS-A 5496–5498 (field numbers NG 277–279), and one female, UIS-A 5495 (field number NG 276) from El Manantialito creek , Nuevas Ideas farm, Sierra de Perijá , vereda Barriales-Nuevas Ideas, El Molino municipality, La Guajira department, Colombia (10°35’15.87”N – 72°49’06.90”W; 1,754 m asl), collected on 19 July 2012, by Fabio L. Meza-Joya. GoogleMaps
Referred specimens. Three specimens (two juveniles and one metamorph): juvenile MHNLS 18858 (field number SP 65) from the creek behind the house at Cerro Las Antenas, Sierra de Perijá, Rosario de Perijá, municipality, Zulia state, Venezuela (10°20’37.0”N – 72°33’41.0”W; 1,449 m asl), collected on 27 March 2008 by Pedro Cabello and Edwin Infante; juvenile MHNLS 19254 (field number SP 312) with the same locality, date and collectors as the holotype; metamorph UIS-A 5499 (field number NG 280) from El Manantialito creek, Nuevas Ideas farm, Sierra de Perijá, vereda Barriales-Nuevas Ideas, El Molino municipality, La Guajira department, Colombia (10°35’16.0”N – 72°49’12.0”W; 1,962 m asl), collected on 19 July 2012, by Fabio L. Meza-Joya.
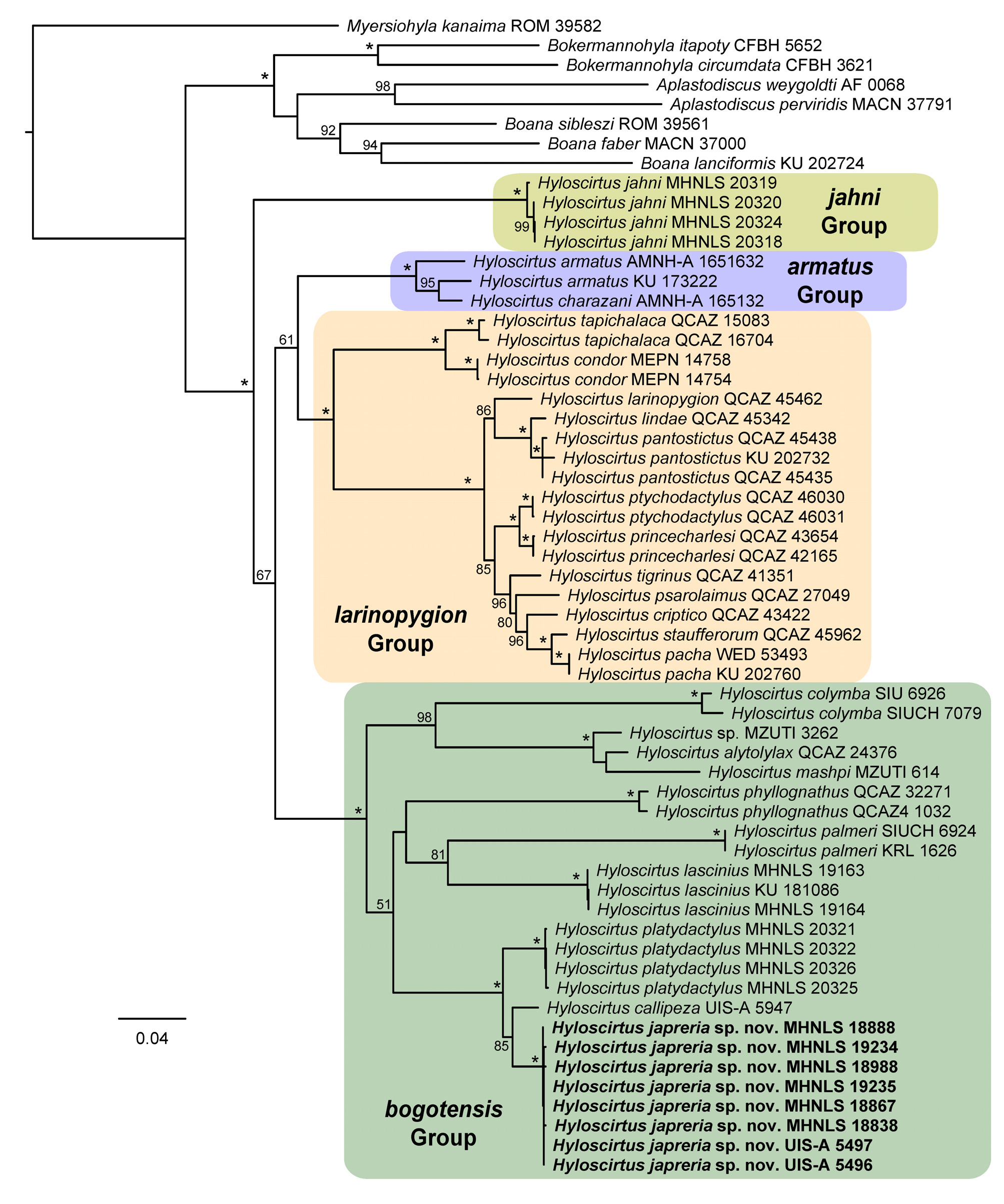
Definition. A new species of Hyloscirtus , assigned to the H. bogotensis species Group ( sensu Duellman 1972 , and Faivovich et al. 2005), by its phylogenetic placement ( Fig. 1 View FIGURE 1 ) and morphologically, by the presence of a mental gland in males, and white parietal peritoneum. The new species is defined by the unique combination of the following characters: (1) Adult males smaller than females (males: 28.8–32.7 mm of SVL vs. females: 35.6–39.1 mm); (2) skin texture smooth on dorsum, flanks, throat, chest, forearms, dorsal and inner surfaces of thighs; areolate on ventral surface of thighs and coarsely areolate on belly; (3) body slender (2.4 to 3.0 times longer than wide); (4) snout rounded in dorsal view and profile; (5) tympanum distinct, small (td: 3.4–5.0% of SVL); tympanic membrane not differentiated; tympanic annulus visible through the skin; about 1/4 of the tympanum concealed dorsally by the supratympanic fold; (6) dentigerous processes of vomers conspicuous, straight, narrowly separated from each other; with 5–8 teeth each; (7) vocal slits longitudinal, extending from sides of tongue to the angle of mouth; (8) vocal sac bilobate, median and subgular, moderately distensible and evident externally; (9) mental gland present in males, disc-shaped, small, covering about the anterior third of throat; (10) ulnar fold present; (11) fingers with fleshy dermal fringes; (12) fingers II–IV basally webbed; (13) nuptial pads absent; (14) prepollex swollen, elliptical, not modified as a protruding spine; (15) calcar tubercle absent; (16) outer and inner tarsal folds present, inner poorly defined; (17) toes with wide lateral fringes; (18) toes extensively webbed; (19) supracloacal fold present; (20) cloacal sheath short; (21) cloaca surrounded by numerous warts; (22) in life, dorsum usually pale yellow to greenish yellow, with dense dark brown punctuation, and several reddish brown spots; (23) whitish stripes on external border of upper eyelids and supratympanic folds, longitudinally on the mid-dorsum, on supracloacal fold, outer ulnar folds, inner and outer tarsal folds, and also on dorsal internal surface of shanks; (24) white warts around the cloaca; (25) axillae bright yellow and groins ocher yellow in life; (26) interdigital membranes on hands and feet bright yellow in life; (27) white parietal peritoneum covering ventral surface of abdominal cavity almost entirely; (28) iris gray with thin black reticulation.
Diagnosis. Hyloscirtus japreria sp. nov. is readily distinguishable from all other species in the H. armatus and H. larinopygion groups by the presence of mental gland in males (present in almost all species of H. bogotensis species group). Of the 16 species currently included in the H. bogotensis Group only five are known to occur on the northwestern portion of the Andes (northern Cordillera Oriental of Colombia, Sierra de Perijá and Cordillera de Mérida in Venezuela); these are: H. callipeza , H. estevesi , H. jahni , H. lascinius and H. platydactylus ( Fig. 4 View FIGURE 4 ).
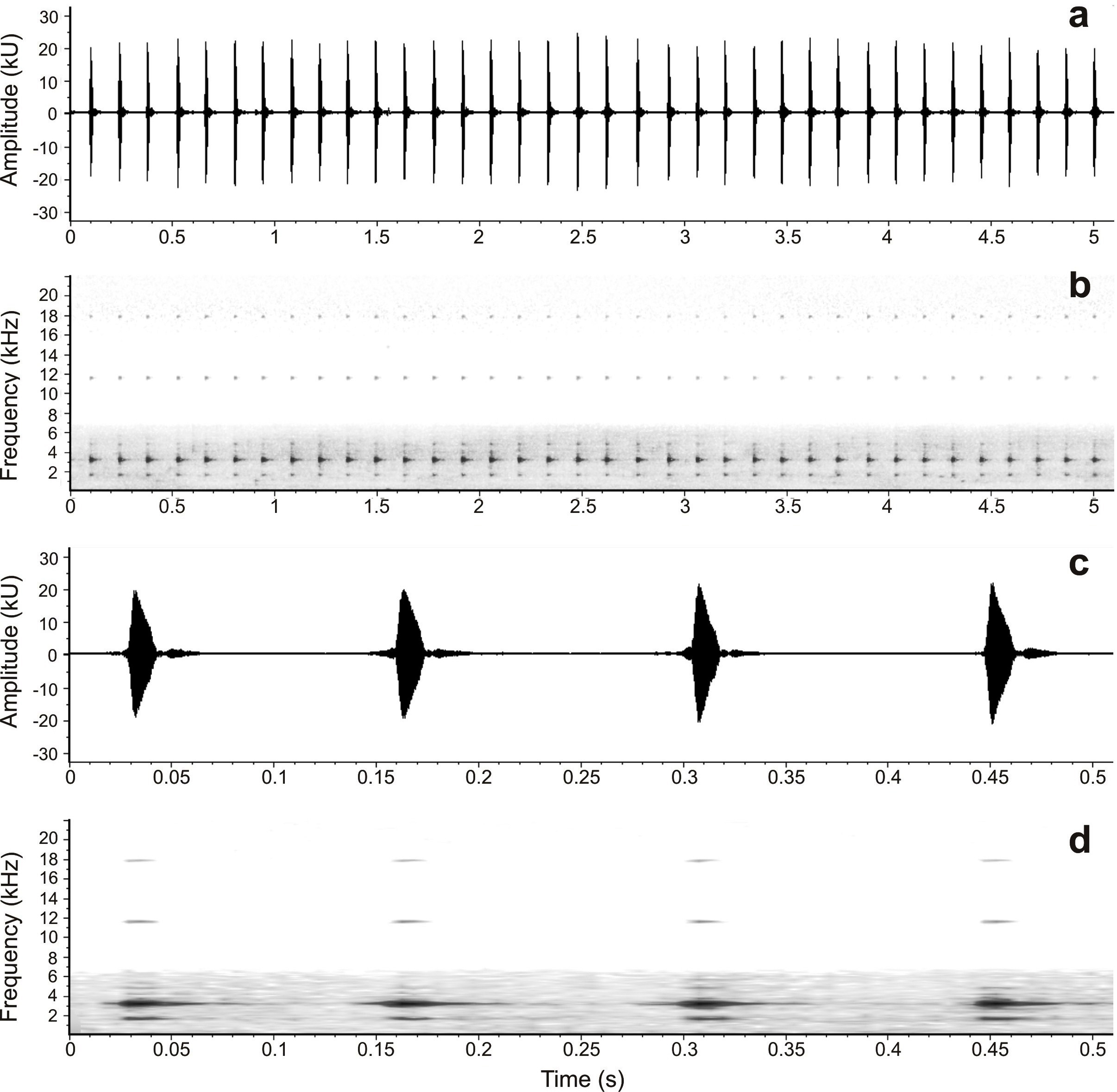
Hyloscirtus japreria sp. nov. differs from H. callipeza (characters of the latter in parenthesis) by having ulnar, outer and inner tarsal folds (absent), snout rounded in profile (truncate), tympanic annulus distinct (barely visible), skin of belly coarsely areolate (granular), webbing bright yellow in life (pale orange), by having a whitish ulnar stripe (absent), a whitish stripe on dorsal internal surface of shank (absent), and iris gray with thin black reticulation (bronze with thin black reticulation); also adult females of H. japreria sp. nov. are larger than females of H. callipeza (maximum SVL: 39.1 mm in H. japreria sp. nov. vs. 36.7 mm in H. callipeza ). These two species also differ in temporal and spectral characteristics of advertisement calls: H. japreria sp. nov. emits much longer trills (79.72– 159.62 s vs. 4.99– 17.50 s), and more notes by trill (421–1,058 vs. 29–102 notes) than H. callipeza ; the notes of H. japreria sp. nov. are also longer (0.037– 0.046 s vs. 0.016–0.019) and less spaced (time between notes: 0.101– 0.105 s vs. 0.146– 0.181 s) than those of H. callipeza . The two species also differ in the number of amplitude peaks per note, with a single peak in H. japreria sp. nov. ( Fig. 7c View FIGURE 7 ) and three peaks of amplitude per note in H. callipeza ( Fig. 11c View FIGURE 11 ). Finally, both peak and fundamental frequencies are lower in H. japreria sp. nov. than in H. callipeza (peak frequency: 3.08–3.15 kHz vs. 3.20–3.33 kHz; fundamental frequency: 1.54–1.59 kHz vs. 1.59– 1.65).
Hyloscirtus jahni View in CoL is known only from Cordillera de Mérida in the Venezuelan Andes ( La Marca 1985; Barrio 2004). The new species can be readily distinguished from H. jahni View in CoL by lacking spicules on the skin in males (spicules present on snout, lips, chin, chest, belly, upper arms, forearms, thighs, shanks, hands, and fingers of adult males), snout not projected beyond lip (snout projected), supratympanic fold distinct (strong), head narrower than body (head wider than body), vocal sac bilobate (single), mental gland small and discoidal (large and transversely elliptical), and adult females larger than those of H. jahni View in CoL (maximum SVL 39.1 mm in H. japreria View in CoL sp. nov. vs. 34.4 mm in H. jahni View in CoL ). These two species also differ in temporal characteristics of advertisement calls (parameter values of H. jahni View in CoL taken from La Marca [1985]): The calls of H. japreria View in CoL sp. nov. consist of long trills of tonal notes (79.72– 159.62 s) whereas the call of H. jahni View in CoL is composed of single pulsed notes. Hyloscirtus japreria View in CoL sp nov. presents a single amplitude peak per note while each note of H. jahni View in CoL contains twelve peaks of amplitude. The notes of H. japreria View in CoL sp. nov. are also longer (0.037– 0.046 s vs. 0.013 s) and less spaced (time between notes: 0.101– 0.105 s vs. 0.3 s) than those of H. jahni View in CoL . Finally, the rate of note emission is higher in H. japreria View in CoL sp. nov. than in H. jahni View in CoL (5.30–6.71 vs. 3.0 notes/s). Hyloscirtus japreria View in CoL and H. jahni View in CoL are not sister species in our phylogeny ( Fig. 1 View FIGURE 1 ).
Hyloscirtus estevesi View in CoL is known only from his type locality in the Río Albarregas, Mérida state and Boconó, Trujillo state (both localities in the Cordillera de Mérida). This species was originally described as a centrolenid ( Centrolenella estevesi ) and recently assigned to the H. bogotensis View in CoL species Group by Faivovich et al. (2005). The new species can be readily distinguished from H. estevesi by not having the snout projected beyond lip (snout projected), by having the head narrower than body (head wider than body), and by having melanophores on dorsal surface of FII–IV (melanophores only present on FIV).
Hyloscirtus lascinius View in CoL occurs on western versant of Cordillera Oriental in Colombia and Cordillera de Mérida in Venezuela ( Rojas-Runjaic et al. 2016). Hyloscirtus japreria View in CoL sp. nov. also is readily distinguishable from this species by not having calcar tubercle (present, large and triangular), by having tympanic annulus distinct (barely visible), snout rounded in profile (snout sloped in profile), skin on belly coarsely areolate (granular), dorsum not sprinkled with whitish dots (dorsum sprinkled with whitish dots), by having, in life, axillary and inguinal regions bright yellow and bright ocher, respectively (axillary and inguinal regions greenish blue), supratympanic fold delineated with a whitish stripe (supratympanic fold without a whitish stripe), and iris gray with thin black reticulation (iris golden with thin reddish reticulation). These two species also differ in temporal and spectral characteristics of advertisement calls (parameter values of H. lascinius View in CoL taken from Rojas-Runjaic et al. [2016]): H. japreria View in CoL sp. nov. emits longer note trills (421–1,058 notes by trill) whereas H. lascinius View in CoL emits single notes or groups of two to four notes. The notes of H. japreria View in CoL sp. nov. are shorter (0.037– 0.046 s vs. 0.091–0.164) and less spaced (time between notes: 0.101– 0.105 s vs. 0.700– 1.075 s) than those of H. lascinius View in CoL . Finally, both peak and fundamental frequencies are higher in H. japreria View in CoL sp. nov. than in H. lascinius View in CoL (peak frequency: 3.08–3.15 kHz vs. 2.13–2.37 kHz; fundamental frequency: 1.54–1.59 kHz vs. 1.08–1.22). Hyloscirtus lascinius View in CoL and H. japreria View in CoL sp. nov. are not sister species in our phylogeny ( Fig. 1 View FIGURE 1 ).
Hyloscirtus platydactylus View in CoL is known from the eastern slope of Cordillera Oriental in Colombia (Acosta-Galvis 2000) and Cordillera de Mérida in Venezuela ( La Marca 1985, Barrio 2004). Hyloscirtus japreria View in CoL sp. nov. is similar to H. platydactylus View in CoL , but can be distinguished from the latter by having ulnar, outer and inner tarsal folds (ulnar, outer and inner tarsal folds absent); mental gland small, disc-shaped, covering about the anterior third of throat (larger, rhomb-shaped, covering about the anterior half of throat); eye diameter greater than eye-to-nostril distance (eye diameter equal to eye-to-nostril distance); whitish stripe extending on outer border of the upper eyelid and on supratympanic fold (whitish stripe absent); whitish stripes on supracloacal fold, ulnar fold, outer and inner tarsal folds, and also on dorsal internal surface of shanks (whitish stripes on supracloacal, ulnar, outer and inner tarsal folds, and on dorsal internal surface of shank, absent); dorsal surfaces of fingers I–II and toes I–III immaculate bright yellow in life (ocher yellow, densely stippled with blackish brown); dorsal surfaces of finger discs and toe discs spotless (spotted with blackish brown); iris gray with thin black reticulation (golden to copper with black reticulation). These two species also differ in temporal and spectral characteristics of advertisement calls (parameter values of H. platydactylus View in CoL taken from Duellman [1972] and La Marca [1985]): The notes emitted by H. japreria View in CoL sp. nov. are longer (0.037– 0.046 s vs. 0.020) and less spaced (time between notes: 0.101– 0.105 s vs. 0.140 s) than those of H. platydactylus View in CoL . The two species also differ in the maximum rate of note emission, that is lower in H. japreria View in CoL sp. nov. than in H. platydactylus View in CoL (6.71 vs. 9.00 notes/s). Peak frequency is also the fundamental in H. platydacylus (2.50–3.49 kHz) and includes in its variation the peak frequency of H. japreria View in CoL sp. nov. (3.08–3.15 kHz), but fundamental frequency of this last species is lower than that of H. platydacylus (1.54– 1.59 vs. 2.50–3.49 kHz). Hyloscirtus platydactylus View in CoL and H. japreria View in CoL sp. nov. are not sister species in our phylogenetic analysis ( Fig. 1 View FIGURE 1 ).
From the remaining species of the Hyloscirtus bogotensis View in CoL species Group, H. japreria View in CoL sp. nov. is also readily distinguishable by the following characters: from H. denticulentus View in CoL , H. palmeri View in CoL and H. piceigularis View in CoL by lacking calcars (present in all these three species); from H. albopunctulatus View in CoL and H. phyllognathus View in CoL by lacking white spots on the dorsum (present in both species); from H. alytolylax View in CoL and H. colymba View in CoL by having an inner tarsal fold delineated with a whitish stripe (absent in both species); from H. lynchi View in CoL by having tarsal folds and mental gland evident externally in males (tarsal folds absent and mental gland not evident in H. lynchi View in CoL ), from H. bogotensis View in CoL by having tarsal and supracloacal whitish stripes (both stripes absent in H. bogotensis View in CoL ); from H. simmonsi View in CoL and H. torrenticola View in CoL by having ulnar and tarsal folds (absent in both); finally, from H. mashpi View in CoL by having an inner tarsal fold delineated with a whitish stripe (inner tarsal fold and whitish stripe absent in H. mashpi View in CoL ) and by lacking a dark brown middorsal stripe (present in H. mashpi View in CoL ).
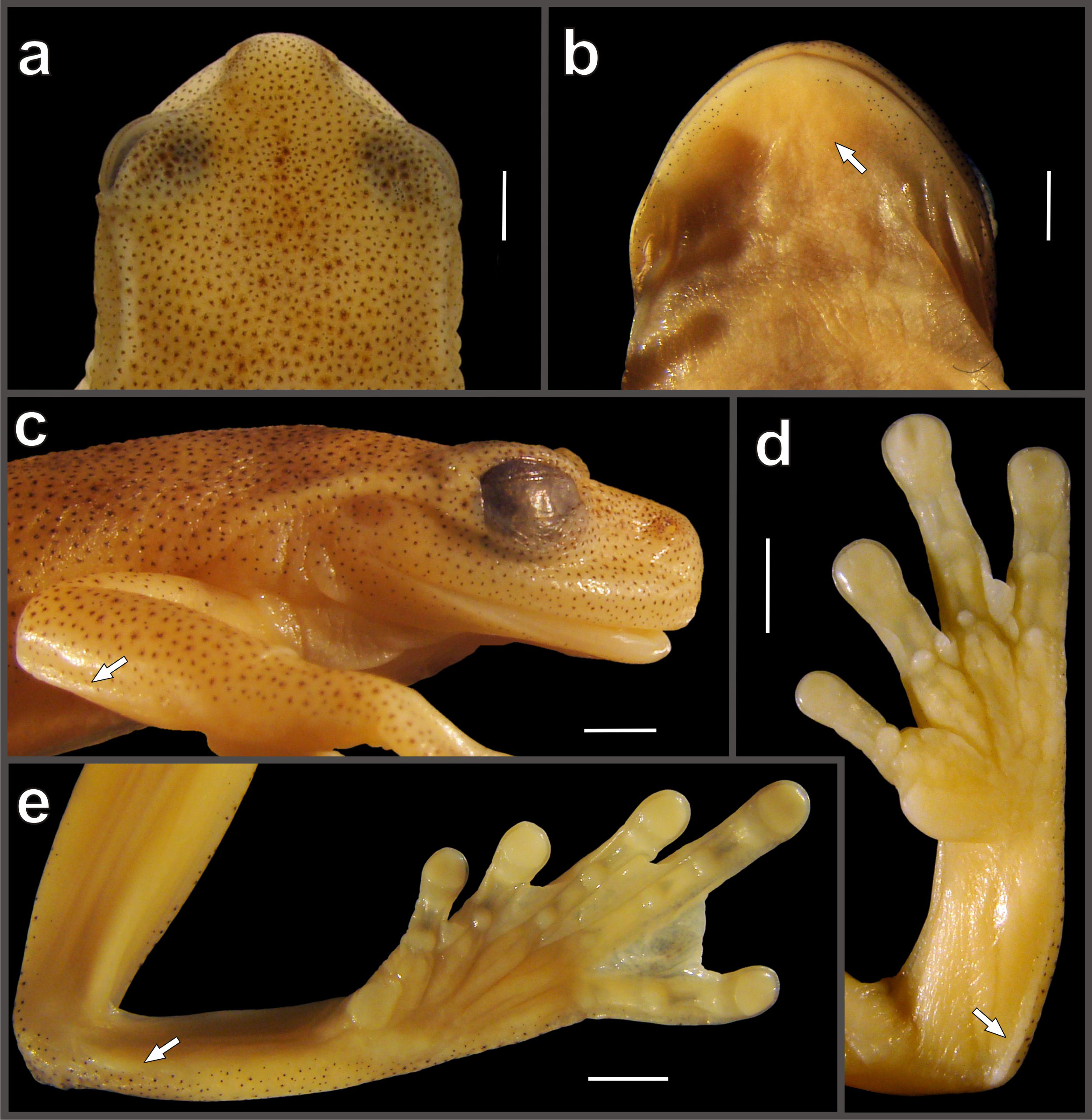
Description of the holotype. An adult male ( Figs. 2a–b View FIGURE 2 ) of 29.9 mm SVL. Body relatively slender (about 2.4 times longer than wide). Head slightly wider than long (HW: 35.6% of SVL; HeL: 32.9% of SVL; HeL/HW: 0.9). Snout rounded in dorsal view and profile ( Figs. 3a,c View FIGURE 3 ). Eye-naris distance slightly shorter than eye diameter (EN/ ED: 0.9). Canthus rostralis distinct, slightly concave in dorsal view, rounded in cross section; loreal region concave and sloped; lips rounded, not flared; nostrils slightly protruded anterolaterally. Internarial region and top of head flat. Interorbital distance wider than upper eyelid. Eye prominent (ED: 10.2% of SVL). Tympanum distinct, small (TD/ED: 0.4; TD: 4% of SVL); membrane not differentiated but most of circumference of tympanic annulus visible through the skin; about 1/4 of the tympanum concealed dorsally by the supratympanic fold. Supratympanic fold developed, starting at posterior end of upper eyelid and reaching posterior margin of insertion of arm ( Fig. 3c View FIGURE 3 ). Mental gland present, small (diameter about 11% of SVL), disc-shaped, covering the anterior third of throat ( Fig. 3b View FIGURE 3 ). Dentigerous processes of vomers conspicuous, straight, narrowly separated from each other; each process bears 5/6 (right/left) teeth. Choanae large, elliptical, not concealed by palatal shelf of maxillary arch. Tongue oval, completely attached to the floor of mouth; vocal slits large, longitudinal, originating on sides of tongue and extending diagonally toward angle of mouth. Vocal sac moderately distensible, evident externally, bilobate, median and subgular.
Forearm wider than upper arm; axillary membrane absent. Outer ulnar fold present ( Fig. 3c View FIGURE 3 ). Fingers short and thick; discs on fingers round, slightly expanded laterally, with clearly defined circumferential groove; disc on FIII slightly wider than tympanum diameter. Relative lengths of fingers: I<II<IV<III. Fingers with fleshy dermal fringes; webbing present among fingers II–IV ( Fig. 3d View FIGURE 3 ); webbing formula II 2 -–3+ III 2 ½–2+ IV. Subarticular tubercles swollen, round or oval; distal tubercle of FIV larger than all others and weakly bifid. Supernumerary tubercles present, small and rounded. Palmar tubercle small, flat, bifid, poorly defined. Prepollical tubercle elliptical, thick, not modified as a spine. Nuptial excrescences absent.
Hind limbs robust; ThL: 46.0% SVL; SL: 50.3% SVL; FL: 40.6% SVL. Calcar tubercle absent. Outer and inner tarsal folds present ( Fig. 3e View FIGURE 3 ), inner poorly defined. Toes short, with wide lateral fringes; toe discs small, rounded, barely expanded, slightly smaller than those on fingers. Relative length of toes: I<II<III<V<IV; extensive toe webbing, formula: I 2 -–2+ II 1 +–2½ III 1 ½–2½ IV 2 +–1+ V. Outer metatarsal tubercle small, rounded, slightly elevated; inner metatarsal tubercle elliptical, flat, about three times the size of the outer; subarticular tubercles round, moderately elevated; supernumerary tubercles present, small and rounded.
Cloacal opening directed posteroventrally at mid-level of thighs; cloacal sheath short; margins of vent with numerous small folds; numerous warts around the cloaca; supracloacal fold present. Skin on dorsum, flanks, throat, chest, forearms, dorsal and inner surfaces of thighs, smooth; areolate on ventral surface of thighs and coarsely areolate on belly. White parietal peritoneum present, covering almost all ventral surface of abdominal cavity.
Color in life of the holotype (based on photographs). Dorsal background color pale yellow, densely and finely punctuated with dark brown, and with scattered small reddish brown spots. Flanks paler than dorsum and almost without dark brown punctuation. Ventral surfaces immaculate yellowish cream, except margins of the lower jaw that are punctuated with dark brown. Whitish stripes present on external border of upper eyelids and supratympanic folds, longitudinally on the mid-dorsum, on supracloacal fold, ulnar fold, inner and outer tarsal folds, and on dorsal internal surface of shanks. White warts around the cloaca. Axillae bright yellow; groins bright ocher. Fingers I–II, toes I–III, and interdigital membranes of fingers and toes immaculate bright yellow. Iris gray with thin black reticulation.
Color of the holotype in preservative (after eight years, May 2017). Dorsal and ventral background color pale cream ( Fig. 2a–b View FIGURE 2 ). Head, lower jaw, and all dorsal surfaces (except fingers I–II and toes I–III), with dense and fine pale brown punctuation. Whitish stripes on border of upper eyelids, inner and outer tarsal fold, dorsal internal surface of shanks, and mid-dorsal, all barely visible; whitish and more defined stripes on the supratympanic, ulnar and supracloacal folds. Warts around the cloaca with faint whitish coloration.
Measurements of the holotype (in mm). SVL: 29.9; SL: 15.0; ThL: 13.7; FL: 12.1; HaL: 9.4; HeL: 9.8; HW: 10.6; IO: 4.1; InD: 2.6; EN: 2.6; ED: 3.0; TD: 1.2; ETS: 4.5; F3D: 1.5; T4D: 1.4; F1L: 5.5: F2L: 7.0. See Table 2 for measurements of the type series.
Morphological and color variation. Hyloscirtus japreria sp. nov. exhibits sexual dimorphism in size ( Fig. 2 View FIGURE 2 ), with adult females larger than males (males: 28.8–32.7 mm [30.3 ± 1.1; n = 17] of SVL vs. females: 35.6–39.1 mm [37.1 ± 1.4; n = 5]). Mental gland, vocal slits and vocal sac are secondary sexual characters present in males and absent in females. Variation of general morphometric characters of the type series is shown in Table 2. The proportions of the head (slightly wider than long to as wide as long) are very similar in the type series and do not differ between sexes (HeL/HW: 0.9–1.0; n = 22), but the relative size of the head is slightly larger in males (HeL: 29.4–33.5% of SVL [32.1 ± 1.0; n = 17]; HW: 31.8–37.0% of SVL [34.6 ± 1.3; n = 17]) than in females (HeL: 28.7–30.4% of SVL [29.7 ± 0.7; n = 5]; HW: 28.8–33.7% of SVL [32.0 ± 2.0; n = 5]). The tympanum is relatively small (3.4–5.0% of SVL [4.2 ± 0.4; n = 22]) and its ratio with respect to the eye has little variation between sexes (TD/ED: 0.3–0.4 [n = 17] in males vs. 0.4–0.5 [n = 5] in females). EN/ED ratio is also variable; however, in all but one specimen of the type series, eye-naris distance is smaller than eye diameter (0.6–1.0 [0.8 ± 0.1; n = 22]). The eye is prominent and relatively bigger in males than females (ED: 10.2–12.0% of SVL [10.8 ± 0.6; n = 17] in males vs. 9.2–10.4% of SVL [9.9 ± 0.5; n = 5] in females). ETS/ED ratio is also slightly variable but snout is always longer than eye diameter (1.1–1.5 [1.3 ± 0.1; n = 22]). Hand length ranges from 24.5–31.6% of SVL (29.0 ± 1.7; n = 22). Thigh length ranges from 43.5–48.9% of SVL (46.4 ± 1.6; n = 22); shank length ranges from 44.8–52.8% of SVL (49.2 ± 1.9; n = 22), and foot length ranges from 35.2–42.8% of SVL (38.8 ± 1.7; n = 22). Hand webbing variation: II (2-–2+)–(3+–3¼) III (2½–3-)–(2+–2½) IV; modal hand webbing formula: II 2 -–3+ III 2 ½–2+ IV. Foot webbing variation: I 2 -–(2+–2½) II (1+–1½)–(2–3-) III (1+–1½)–(2+–3-) IV (2+–3-)–(1+–1¼) V; modal foot webbing formula: I 2 -–2+ II 1 +–2½ III 1 ½–2½ IV 2 ½–1+ V.
Skin texture, warts, folds, shape of the head and aspect of the tympanum do not exhibit relevant variation in the examined specimens. Mental gland very conspicuous and similar to the holotype in almost all males of the type series; roughly pentagonal in shape in MHNLS 18971, and less defined in MHNLS 18837, 19234 and 19237.
Color pattern is variable in Hyloscirtus japreria sp. nov.; metamorphs and juveniles are typically apple green with dorsal and interorbital blackish spots ( Fig. 5f View FIGURE 5 ). Background color in live adults can be pale cream, pale yellow, greenish yellow, pale green, and yellowish brown ( Fig. 5 View FIGURE 5 ); dorsal pale brown punctuation is very variable (in life and in preserved specimens), with some specimens showing dense and finely punctuated patterns, others scattered punctuated, or with scattered reddish brown spots alternated with densely (or scattered) punctuation ( Figs. 5–6 View FIGURE 5 View FIGURE 6 ). Blackish brown canthal stripes and blackish brown spots above the upper eyelids are present in some specimens (male MHNLS 22568, and females, MHNLS 18837, 18867–18868 18972, 18988, 19235). The intensity of the whitish stripes on the different folds is also variable, and the mid-dorsal whitish stripe can be present (males MHNLS 18838, 18971–18972, 19234, 22568, and females MHNLS 18868, 19235, 19166–19167) or absent (males MHNLS 18837, 18867–18868, 18888–18889, 18988, 19168, 19237). Ventrally, some specimens have scattered dots on the mandible (males MHNLS 18837–18838, 18867, 18869, 18888–18889, 18971–18972, 18988, 19168, 19234, and female MHNLS 19166).
Call description. Advertisement calls of Hyloscirtus japreria sp. nov. are formed by very long trills of short tonal notes ( Fig. 7 View FIGURE 7 ). Notes present amplitude modulation, with a single peak of amplitude ( Fig. 7c View FIGURE 7 ). The average duration of note trills among the three specimens recorded is 114.93 ± 40.24 (79.72–159.62) seconds. The average rate of note emission is 6.36 ± 0.61 (5.30–6.71) notes/s, and each note trill contained in average 738 ± 284.18 (421– 1058) notes. Average note duration is 0.041 ± 0.005 (0.037–0.046) seconds, and the average duration of silent intervals between notes 0.102 ± 0.002 (0.101–0.105) seconds; notes do not have frequency modulation. Average peak frequency is 3.13 ± 0.04 (3.08–3.15) kHz. Lower frequency of notes is 2.99 ± 0.06 (2.92–3.03) kHz on average, and average upper frequency reaches 3.28 ± 0.01 (3.27–3.28) kHz. The fundamental frequency is 1.57 ± 0.02 (1.54–1.59) kHz. At least three other harmonics are detectable in the spectrograms, at 4.63–4.78 kHz, 11.52– 11.58 kHz, and 17.79–17.85 kHz ( Figs. 7b,d View FIGURE 7 ).
Distribution. Hyloscirtus japreria sp. nov. is known from three localities on the eastern versant of the Sierra de Perijá (Zulia state) in Venezuela, and an additional locality on the western versant in Colombia (La Guajira department) ( Fig. 8 View FIGURE 8 ). All localities are between 1,430 and 1,832 m asl. Records of Hyloscirtus platydactylus from Cesar, La Guajira and Norte de Santander departments in Colombia ( Ruiz-Carranza & Lynch 1982, Acosta-Galvis 2000, Romero-Martínez & Lynch 2012) and from Zulia (Sierra de Perijá) in Venezuela ( La Marca 1994) could correspond to H. japreria sp. nov. and need verification.
Habitat and natural history. The three localities where Hyloscirtus japreria sp. nov. was found in the Venezuelan side of the Sierra de Perijá correspond to narrow rapid creeks with numerous small waterfalls, surrounded by primary forest ( Fig. 9a View FIGURE 9 ). The phytocoenoses of these localities correspond to ombrofilous submontane/montane evergreen forest, which ranges from 800–2,500 m asl ( Huber & Alarcón 1988). Colombian specimens (western versant of the Sierra de Perijá) were found at a narrow and partially dry creek, surrounded by secondary forest ( Fig. 9b View FIGURE 9 ). This area corresponds to premontane humid forest ( Holdridge 1967). The region has a biseasonal climatic regime, with a dry season from December–April, and a rainy season from May–November, with maximum rain peaks in May and October, and minimum rain in January and July ( Masciangioli & Febres 2000; IDEAM 2016).
Hyloscirtus japreria sp. nov. is a nocturnal and riparian frog. Its presence in the creeks can be readily detected by its conspicuous vocalization, but calling males are visually difficult to detect because they generally call while hidden. All males collected in the two localities at Cerro Las Antenas were found calling actively at the end of the dry season (March) and during the rainy season (July). Also several males were actively calling in May 2009 at the creek of Guacharaca Camp, and in July 2012 at El Manantialito creek. In all localities calling males were found in a variety of sites, generally hidden in bromeliads ( Fig. 10b View FIGURE 10 ), leaf-sheath of Heliconeaceae plants, within rock crevices, and behind waterfalls ( Fig. 10a View FIGURE 10 ), but occasionally they were calling while exposed on ferns leaves, palms, and bush branches near small waterfalls ( Figs. 5c View FIGURE 5 , 10c View FIGURE 10 ). All females were found on leaves and branches near calling males. Only one female was found at Cerro Las Antenas in March 2008, another three at Guacharaca Camp in May 2009, and one at El Manantialito in July 2012. All these five females contain well-developed eggs in their oviducts (noted through skin of the flanks). Metamorphs and post-metamorphs ( Fig. 5f View FIGURE 5 ) were seen in March 2008 at the creeks of Cerro Las Antenas, and in July 2012 at El Manantialito creek. All the specimens collected released a strong odor like moist leaf litter when they were handled.
Etymology. The specific epithet japreria is a nominative used as a noun in apposition. Japreria is an indigenous ethnic group of Carib linguistic affiliation, self-designates as Yankshit. Originally, they lived in the Colombian side of the Sierra de Perijá, and later moved to the Venezuelan side along the Colombian border ( Halbmayer 2013). The Japreria had been traditionally considered as a subgroup of the Yukpa ethnic group ( De Villamañán & De Santelos 1957), but currently are recognized as an independent group ( Oquendo 2004). Its population was ca. 200 individuals according to the 2002 census and its idiom is currently considered as threatened ( Lewis et al. 2016). We nominate this new taxon honoring the Japreria people and warning on its cultural loss.
Call description of Hyloscirtus callipeza . Advertisement calls of Hyloscirtus callipeza are composed of short tonal notes arranged in long trills ( Fig. 11 View FIGURE 11 ). All notes have multiple amplitude modulation, with three peaks of amplitude per note ( Fig. 11c View FIGURE 11 ). The average duration of note trills is 10.40 ± 6.43 (4.99–17.50) seconds. The average rate of note emission is 5.72 ± 0.18 (5.51–5.83) notes/s, and each note trill contained in average 59.7 ± 37.9 (29– 102) notes. Average note duration is 0.018 ± 0.001 (0.016–0.019) seconds, and the average duration of silent intervals between notes is 0.157 ± 0.010 (0.146–0.181) seconds; notes do not have frequency modulation. Average peak frequency is 3.27 ± 0.03 (3.20–3.33) kHz. Average low frequency is 2.98 ± 0.03 (2.92–3.06) kHz, and average high frequency is 3.61 ± 0.05 (3.47–3.66) kHz. The fundamental frequency is 1.62 ± 0.02 (1.59–1.65) kHz. No additional harmonics were detected in the spectrograms ( Figs. 11b, d View FIGURE 11 ).








| MHNLS |
Coleccion de Mastozoologia, Museo de Historia Natural de La Salle |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.