
Ecnomiohyla veraguensis, Batista, Abel, Hertz, Andreas, Mebert, Konrad, Köhler, Gunther, Lotzkat, Sebastian, Ponce, Marcos & Vesely, Milan, 2014
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3826.3.2 |
|
publication LSID |
lsid:zoobank.org:pub:4BEDE115-8C6E-46C6-AF6A-1F96209079ED |
|
DOI |
https://doi.org/10.5281/zenodo.5632097 |
|
persistent identifier |
https://treatment.plazi.org/id/BF3C3F31-134F-1967-9AF6-B20F39BFFDB9 |
|
treatment provided by |
Plazi |
|
scientific name |
Ecnomiohyla veraguensis |
| status |
sp. nov. |
Ecnomiohyla veraguensis View in CoL sp. nov.
Ecnomiohyla rabborum – Köhler 2011: p. 224 Fig. 537; p. 226 Fig. 541 b.
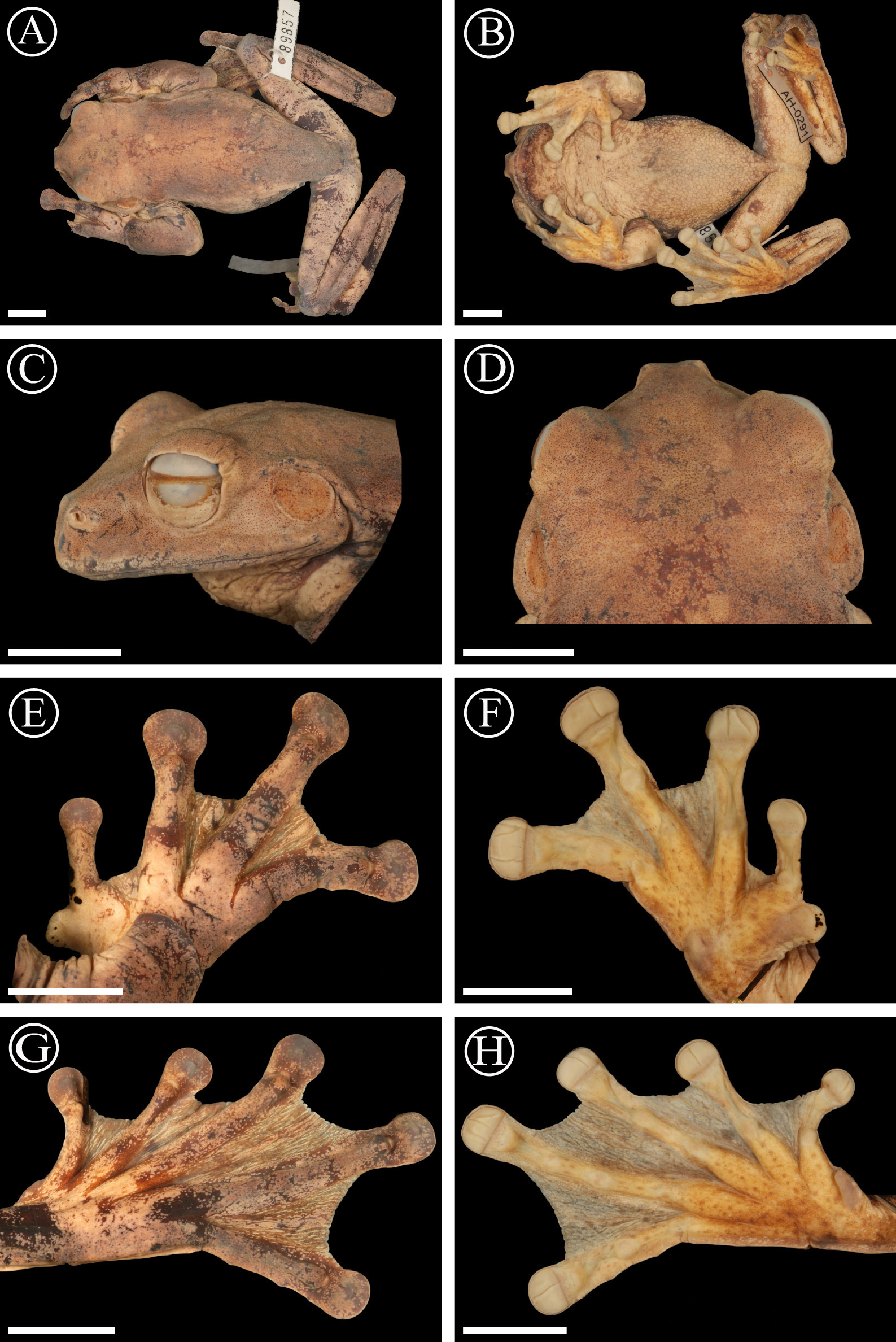
Holotype. SMF 89877 (original field number AH210) an adult male ( Figs. 7–8 View FIGURE 7 View FIGURE 8 ) collected near Cerro Negro (8.5533 °N, - 81.09261 °W, 540 m a.s.l.), Santa Fé National Park, Veraguas, Panama, on 31 March 2009 at 12:00 hrs, collected by Smelin Abrego, Arcadio Carrizo, Andreas Hertz, and Sebastian Lotzkat.
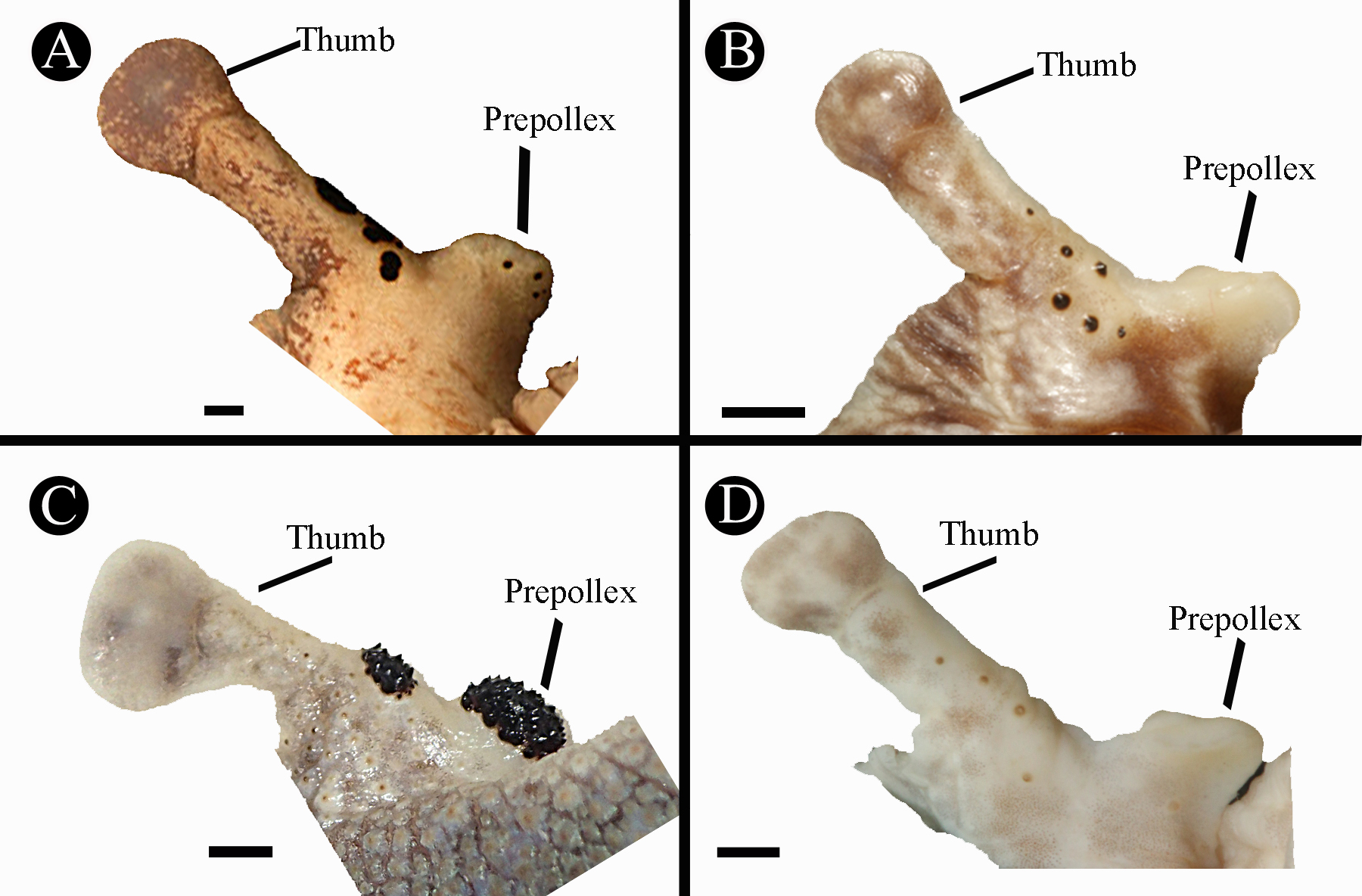
Diagnosis. A medium-sized species of Ecnomiohyla . The single known specimen is an adult male, 57.8 mm in SVL) differing from other known species of the genus by following combination of characters: 1) finger webbing extensive, web touching the finger disk on at least one side on Fingers II–IV; 2) toes extensively webbed; web reaching the toe disk at least on one side on four toes; 3) skin on dorsum finely tuberculate with scattered minute keratin tipped tubercles posteriorly; 4) cranial and dorsal osteoderms present; 5) skin on upper surface of head not co-ossified with underlying cranial elements; 6) humerus without enlarged crista lateralis; 7) prepollex distinct, recurved, with distinct bony prepollical projection, spadelike and directed laterally; 8) 6–8 widely spaced, keratinized black spines present bordering the outer side of the thumb; 9) a distinct scalloped fringe without pointed tubercles on its ventral surface, arising at the heel and continuing on the outer side of Toe V and reaching almost to the disk of Toe V ( Fig. 8 View FIGURE 8 ); 10) dorsal coloration in life smoke gray, with upper surface of forearms bearing a suggestion of lime green ( Fig. 7 View FIGURE 7 ).
Comparison with other species of Ecnomiohyla . Ecnomiohyla veraguensis can be distinguished from other species of Ecnomiohyla by the following characters (with contrasting features for E. veraguensis in parentheses; see Table 3 View TABLE 3 for more details): E. echinata , E. minera , E. rabborum , E. salvaje , E. thysanota , and E. valancifer can be distinguished from the new species by the lack of cranial and dorsal osteoderms (both present); E. rabborum and E. minera are further distinct in having a humeral projection in males (no humeral projection); E. rabborum has substantial finger webbing, web reaching base of disk on one finger (extensive webbing, web touching the finger disk on at least one side on Fingers II–IV); E. fimbrimembra ( Fig. 7 View FIGURE 7 E-F), E. miliaria and E. phantasmagoria lack scalloped fleshy fringes on heels and have pointed heel tubercles instead (scalloped fleshy fringes present, no heel tubercles); E. fimbrimembra and E. salvaje have the skin on the head co-ossified with the cranium, (skin not coossified with cranium); males of E. miliaria and E. phantasmagoria have a sharp prepollical spine protruding from the prepollex (prepollex recurved, no protruding spine); E. bailarina has a strongly tuberculate dorsum (finely tuberculate) and two clusters of numerous, small nuptial spines at the distal end of the prepollex and the base of the pollex (only 6–8 larger, widely spaced nuptial spines along the outer side of the pollex; Fig. 10 View FIGURE 10 ); E. tuberculosa lacks an enlarged prepollical bony projection or keratinized black spines on the prepollex in adult males (enlarged prepollical bony projection and 6–8 widely spaced, keratinized black spines presents); E. miotympanum lacks of scalloped dermal fringes on the outer margin of the forearm and foot, large digital disks, and enlarged prepollices (present in E. veraguensis ); in terms of general appearance, E. sukia is most similar to the new species, but differs by a genetic distance in the 16S gene of 7% and the lack of nuptial spines in adult males (6–8 widely spaced nuptial spines on the outer side of the pollex; see Fig. 9 View FIGURE 9 (B, D) and 10); further, E. sukia lacks keratin tipped tubercles on the dorsum (presence of keratin tipped tubercles on the dorsum).
Description of the holotype. An adult male, as determinated by the presence of nuptial spines, and vocal slits. Measurements of the holotype are indicated in Table 4 View TABLE 4 . Head rounded in dorsal view, wider than long (HL/HW= 85.6%); snout truncate in dorsal and lateral views; nostrils directed laterally; top of head flat; canthus rostralis concave; loreal region concave; a well-developed supratympanic fold running from above the upper margin of tympanum, slightly curved around its upper posterior edge; tympanum prominent, smooth, same color as dorsum, and 60.0% of ED; separated from eye by 3.04 mm; upper surface of body finely tuberculated, scattered tubercles present on dorsal surfaces of the limbs; arms robust, hypertrophied; a fleshy scalloped fringe extends from the elbow along the ventrolateral margin of forearm and continues along the outer edge of Finger IV to base of disk; scallops of fringe largest on forearm, weak scallops along finger; hands moderate in length (HAL/SVL= 36.0%); finger lengths I<II<IV<III, terminal disk on Finger I 70 % of diameter of disks on Fingers II–IV, which are slightly smaller in size than tympanum (3FD/TD= 94%); distal subarticular tubercles on fingers rounded and elevated; a row of supernumerary tubercles present under the first phalanges on Fingers II–III; palmar tubercles rounded and low; prepollex enlarged and recurved, bony prepollical projection spadelike, directed laterally at side adjacent to thumb; 6–7 nuptial spines along the outer side of the pollex; fingers extensively webbed, web extending to base of disks on at least two fingers; webbing formula: I 1 3/4–2 II 3 /4–1 1/4 III 1 1/4–3/ 4 IV; legs relatively long and slender (FL/ SVL= 48.4%), heels of adpressed limbs overlapping about 1/3 length of tibia, thigh 26.70 mm long; distinct fleshy, scalloped fringe begins on heel and extends along ventrolateral margin of tarsus and outer margin of Toe V to base of disk; scallops sinuously serrated, widest on tarsus, smaller along toe; tarsal fold slightly evident; outer metatarsal tubercle barely distinct, inner metatarsal tubercle large (1.12 times 3TD), ovoid, slightly elevated, and spadelike distally; toe lengths I<II<III>V<IV; disks on toes 80–91% of diameter of those on fingers, disk on Toe IV same size as disks on Toes III and V, decreasing in size on Toes II–I; subarticular tubercles rounded and slightly elevated; a row of 5–10 supernumerary tubercles barely distinct under the proximal phalanges on toes; extensive toe webbing, webs extending to base of disks on at least four toes; webbing formula: I 3 /4–1 1/2 II 3 /4–1 1/4 III 3 /4–3/4 IV 1 /4– 3/ 4 V; gular area and venter granulate, fine granulation on undersides of arms and proximal thighs, smooth skin on anterior surfaces of thighs and ventral parts of legs; cloacal opening directed posteriorly at mid-level of thighs, a distinct granular dermal fold under the vent. Tongue slightly cordiform, broader at the base; vomerine ridges large and transverse, well separated medially, placed between the posterior margins of choanae; vomerine teeth 10–14; paired vocal slits extending posteriorly from posterior lateral base of tongue toward angle of jaws.
Coloration of holotype in life ( Fig. 7 View FIGURE 7 ). Coloration in life was recorded at daytime: Dorsal ground color Smoke Gray (44); snout, canthus rostralis, and supraorbital regions Brownish Olive (29) suffused with Olive Green (Auxiliary 47); upper surfaces of forearms with a suggestion of Lime Green (59); dorsal surfaces of finger webbing like dorsal coloration on body, but toe webbing Vandyke Brown (121); ventral surfaces of chin and body Cream Color (54), spotted with Raw Sienna (136); ventral surfaces of hindlimbs True Cinnamon (139); ventral coloration of toe and finger webbings Vandyke Brown (121).
Coloration in preservative ( Fig. 8 View FIGURE 8 ). Dorsal surfaces Grayish Horn Color (268); snout, canthus rostralis, and supraorbital regions Medium Plumbeus (294); darker bars on upper surfaces of limbs Medium Plumbeus (294); cloacal region Pale Buff (1), suffused with Medium Plumbeus (294); groin and posterior surfaces of thighs mottled with Maroon (39) on a Pale Buff (1) ground; ventral regions Pale Buff (1); chin suffused with Maroon (39); toe and finger webbing Burnt Umber (48).
Distribution and natural history. Ecnomiohyla veraguensis is known only from the type locality in the Isthmian-Pacific moist forests ( Fund & Hogan 2012). The holotype was found at noon on a sunny day at the end of the dry season. Relative air humidity at the moment of encounter was 68% at a temperature of 21.8 °C and it was slightly windy. The frog was sitting in a water conserving posture ( Fig. 7 View FIGURE 7 D) on a fern leaf approximately 0.5 m above the ground, next to a water tube that is used by local people to obtain drinking water. Other amphibian species that were observed at Cerro Negro on this expedition conducted between March 31 and April 0 3 2009 include Atelopus varius ( Lichtenstein & Martens, 1856) , Bolitoglossa colonnea ( Dunn, 1924) , Craugastor gollmeri ( Peters, 1863) , C. megacephalus ( Cope, 1875 “1876“), Diasporus citrinobapheus Hertz, Hauenschild, Lotzkat & Köhler, 2012 , Lithobates warszewitschii ( Schmidt, 1857) , Pristimantis caryophyllaceus ( Barbour, 1928) , P. cerasinus ( Cope, 1875 “1876“), P. cruentus , P. museosus ( Ibáñez, Jaramillo & Arosemena, 1994) , P. pardalis ( Barbour, 1928) , Rhaebo haematiticus Cope, 1862 , and Sachatamia albomaculata ( Taylor, 1949) . Since E. veraguensis is only known from a single specimen from a single locality, the distribution is unknown. It is expected to occur along mid-elevations of the Serranía de Tabasará.
Etymology. The species name is derived from the province name Veraguas where the holotype was found, with the Latin suffix - ensis donating a place or locality. The species name has been chosen to accentuate the particular role the province of Veraguas plays in terms of amphibian conservation. It is the only Panamanian province with Atlantic and Pacific coasts, thus encompassing a great variety of habitats for many amphibian species.
Conservation status. As other Ecnomiohyla species, E. veraguensis could be considered as a rare species, due to the habitats it uses, this fact makes it difficult to assess its populations. Like E. bailarina , the data deficient (DD) criterion, according the IUCN (IUCN 2013), seems appropriate for E. veraguensis too, until data on its population trend become available.


| SMF |
Forschungsinstitut und Natur-Museum Senckenberg |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.