
Cyrtodactylus guakanthanensis, Grismer, L. Lee, Belabut, Daicus M., Quah, Evan S. H., Onn, Chan Kin, Wood, Perry L. & Hasim, Rosli, 2014
|
publication ID |
https://doi.org/10.11646/zootaxa.3755.5.3 |
|
publication LSID |
lsid:zoobank.org:pub:B06C2DCA-78B6-46F9-8D4F-1716DD20C4A5 |
|
DOI |
https://doi.org/10.5281/zenodo.6141125 |
|
persistent identifier |
https://treatment.plazi.org/id/C03587DF-FF9B-FFEC-FF28-FA03739EBA45 |
|
treatment provided by |
Plazi |
|
scientific name |
Cyrtodactylus guakanthanensis |
| status |
sp. nov. |
Cyrtodactylus guakanthanensis sp. nov.
Gua Kanthan Bent-toed Gecko Cicak Gua Kanthan
Figs 3 View FIGURE 3 , 4 View FIGURE 4 , 6
Holotype. Adult male ( LSUHC 11322) from Gua Kanthan, Perak, Peninsular Malaysia ( 4o45.685’N, 101o07.322’E; 45 m) collected by Daicus M. Belabut, Chan K. Onn, Evan S. H. Quah and L. Lee Grismer on 14 July 2013.
Paratypes. All paratypes ( LSUHC 11321, 11323, 11325–36, 11339) bear the same data as the holotype.
Diagnosis. Cyrtodactylus guakanthanensis sp. nov. can be distinguished from all other Sundaland species by having the following suite of character states: adult SVL 77.7–82.2 mm adult SVL; moderately sized, conical, weakly keeled, body tubercles; tubercles present on occiput, nape, and limbs, and extend posteriorly beyond base of tail; 37–44 ventral scales; no transversely enlarged, median, subcaudal scales; proximal subdigital lamellae transversely expanded; 19–21 subdigital lamellae on fourth toe; abrupt transition between posterior and ventral femoral scales; enlarged femoral scales; no femoral or precloacal pores; precloacal groove absent; wide, dark postorbital stripes from each eye extending posteriorly to the anterior margin of the shoulder region thence forming a transverse band across the anterior margin of the forelimbs; and body bearing five (rarely four) wide, bold, dark bands. The meristic characters are scored against C. sworderi , C. tebuensis and C. quadrivirgatus in Table 5 View TABLE 5 and against all other Sundaland species in Grismer et al. (2012b: Table 5 View TABLE 5 ).
=
Description. Adult male SVL 79.7 mm; head large, moderate in length (HL/SVL 0.29) and width (HW/HL 0.65), somewhat depressed (HD/HL 0.40), distinct from neck, and triangular in dorsal profile; lores weakly inflated, prefrontal region slightly concave; canthus rostralis smoothly rounded; snout elongate (ES/HL 0.44) and rounded in dorsal profile; eye large (ED/HL 0.22); ear opening elliptical and small (EL/HL 0.09); eye-to-ear distance greater than diameter of eye; rostral subrectangular with a deep dorsomedial furrow containing a postrostral; rostral partially divided dorsally, bordered posteriorly by large left and right supranasals, one large medial postrostral (=internasal) and one small azygous postrostral; external nares bordered anteriorly by rostral, dorsally by two supranasals (anterior one largest), posteriorly by five postnasals and ventrally by first supralabial; nine (R, L) square to rectangular supralabials extending to and tapering smoothly below posterior margin of orbit; eight (R), seven (L) infralabials tapering smoothly posteriorly to below posterior margin of orbit; scales of rostrum, lores, top of head, and occiput small and granular; scales on top of occiput intermixed with slightly enlarged tubercles; dorsal and ventral superciliaries rectangular; mental triangular, bordered laterally by first infralabials and posteriorly by left and right rectangular postmentals contacting medially for approximately 50% of their length posterior to mental; one enlarged row of sublabials extending posteriorly to 4th or 5th infralabial; gular scales small and raised, grading posteriorly into slightly larger, flatter, throat scales, and thence into large, flat, imbricate pectoral and ventral scales.
Body relatively short (AG/SVL 0.42) with weak, tuberculate ventrolateral folds; dorsal scales small and granular, interspersed with larger, conical, semi-regularly arranged, weakly keeled tubercles; tubercles extending from occiput to anterior one-third of tail; tubercles on occiput and nape relatively small, those on body largest; approximately 17 longitudinal rows of dorsal tubercles and 32 paravertebral tubercles; 37 flat, imbricate ventrals, ventrals much larger than dorsals; patch of enlarged precloacal scales lacking pores; precloacal groove absent.
Forelimbs moderate in stature, relatively short (FL/SVL 0.17); granular scales of forearm larger than those of body and interspersed with small tubercles; palmar scales rounded; digits well-developed, inflected at basal interphalangeal joints; subdigital lamellae slightly enlarged proximal to joint inflections, digits narrower distal to joints; claws well-developed, sheathed by a dorsal and ventral scale.
Hind limbs more robust than forelimbs, moderate in length (TBL/SVL 0.18), covered dorsally by granular scales interspersed with larger tubercles and anteriorly by granular scales; ventral scales of femora flat and larger than dorsals; ventral tibial scales flat and imbricate; a single row of enlarged femoral scales extend medially from proximal region of femur making contact with large precloacal scales; this enlarged row is bordered anteriorly by two rows of slightly smaller scales; femoral pores absent; dorsal and ventral femoral scales meeting abruptly on posteroventral margin of thigh; plantar scales low and slightly rounded; digits well-developed, inflected at basal interphalangeal joints; subdigital lamellae enlarged proximal to inflected joints, digits narrower distal to joints; 20 (L,R) subdigital lamellae on 4th toe; claws well-developed, sheathed by a dorsal and ventral scale.
Original tail widest at base, tapering to a point, approximately last one-fifth of tail missing; dorsal scales on base of tail granular, becoming flatter posteriorly; no median row of transversely enlarged subcaudal scales; caudal scales arranged in semi-whorls; three enlarged tubercles on both sides of base of tail; base of tail bearing lateral, bulbous swellings; all postcloacal scales moderately sized, flat, and imbricate.
Coloration in life ( Fig. 3 View FIGURE 3 ). Ground color of head, body, limbs and anterior portion of tail beige; snout and top of head brown bearing darker mottling; wide, dark brown postorbital stripes from each eye extend posteriorly to the anterior margin of the shoulder region, thence connecting medially and forming a transverse band across the base of the anterior margin of the forelimbs; single wide dark brown chevron-shaped marking on nape and base of occiput; four wide, dark brown body bands between limb insertions that extend laterally one-half way down flanks; single wide, dark brown sacral band; five dark caudal bands becoming darker and wider posteriorly; dorsal surface of limbs beige bearing darker mottling; ventral surface of head, body, and limbs immaculate off-white except for fine black stippling in scales; subcaudal region nearly black with six light bands encircling tail.
Variation. The paratypes closely resemble the holotype in all aspects of coloration and pattern ( Fig. 3 View FIGURE 3 ). The body bands in LSUHC 11325–26, and 11339 are oblique as opposed to being transverse. The regenerated tails of LSUHC 11321, 11323, 11326, and 11339 are mottled and lack banding. Hatchlings and juveniles (LSUDPC 8175– 79) are more boldly marked, having a nearly immaculate ground color and generally darker bands and the posterior one-third of the tail is white ( Fig. 3 View FIGURE 3 )—a character noted in other karst-dwelling species of Cyrtodactylus ( Grismer et al. 2012a). Meristic variation is presented in Table 6 View TABLE 6 .
Distribution. Currently, Cyrtodactylus guakanthanensis sp. nov. is known only from the type locality of Gua Kanthan in the Gunung Kanthan limestone forest, Perak, Peninsular Malaysia ( Fig. 1 View FIGURE 1 ).
Natural history. All lizards specimens were active at night on the limestone walls, adjacent boulders surrounded by limestone forest, and the limestone forest vegetation. Lizards only occurred in areas where cliff faces were eroded and exfoliated, providing cracks and holes in which to take refuge during the day ( Fig. 4 View FIGURE 4 ). No lizards were found in areas where the cliff faces were smooth and bearing no surface irregularities. Many of the specimens observed were found in the adjacent vegetation between 0.5–4 m above the ground on the trunks of trees or on vertical and horizontal branches and vines, indicating that the surrounding forest itself is an integral part of this new species’ habitat preference. One specimen was observed deep within the cave where vegetation was absent. Hatchlings as small as 33.8 mm SVL (LSUDPC 8175; Fig. 3 View FIGURE 3 ) were observed but no gravid females were found, suggesting that the reproductive season occurred in July.
Etymology. The specific epithet guakanthanensis is an adjective and refers to the type locality Gua Kanthan (=Kanthan Cave) within the Gunung Kanthan limestone forest.
Comparisons. Within the Cyrotdactylus sworderi complex, C. guakanthanensis is separated from C. quadrivirgatus by having an abrupt transition between small and large postfemoral scales and a banded dorsal pattern. From C. sworderi it differs in having fewer paravertebral tubercles (32–34 versus 36–38); lacking as opposed to having precloacal pores; having as opposed to lacking enlarged femoral scales; having wide as opposed to narrow whitish caudal bands ( Fig. 1 View FIGURE 1 ); and having a banded as opposed to spotted dorsal pattern ( Fig. 1 View FIGURE 1 ). It differs from C. tebuensis in potentially having few suprlabials (nine or 10 versus 11–13); having wide as opposed to narrow whitish caudal bands which are fewer in number (nine versus 10–14; Fig. 1 View FIGURE 1 ); and having a banded versus a spotted or striped dorsal pattern ( Fig. 1 View FIGURE 1 ). These differences are compared between all members of the C. sworderi complex in Table 5 View TABLE 5 .
The Gunung Kanthan limestone forest surrounds 400 million year old limestone formations that harbor a number of unique plants and animals. Owing to the purity of its limestone, the area is scheduled to be quarried. Our data indicate that Cyrtodactylus guakanthanensis sp. nov. does not range continuously throughout the limestone forest but appears to be restricted to areas that have an eroded complex limestone microhabitat. One of the quarry zones targeted is Gua Kanthan, the type locality of C. guakanthanensis sp. nov. Grinding down the limestone in this area and cutting the surrounding forest will destroy the microhabitat to which this new species has adapted and our data indicate this species may not occur elsewhere.
The discovery of Cyrtodactylus guakanthanensis sp. nov. adds to a growing body of evidence that karst regions should be protected and better studied. If reptiles are an indication of the hidden diversity within these unique habitats, then limestone forests may be some of the most biotically rich habitats in Peninsular Malaysia with a level of herpetological endemism approaching that of Malaysia’s islands (see Chan et al. 2010; Grismer 2008, 2011b; Grismer et al. 2011). Terminating this species before it is discovered, described, and studied is not only illogical, it is tantamount to discarding a wrapped gift before it is opened and its value assessed.
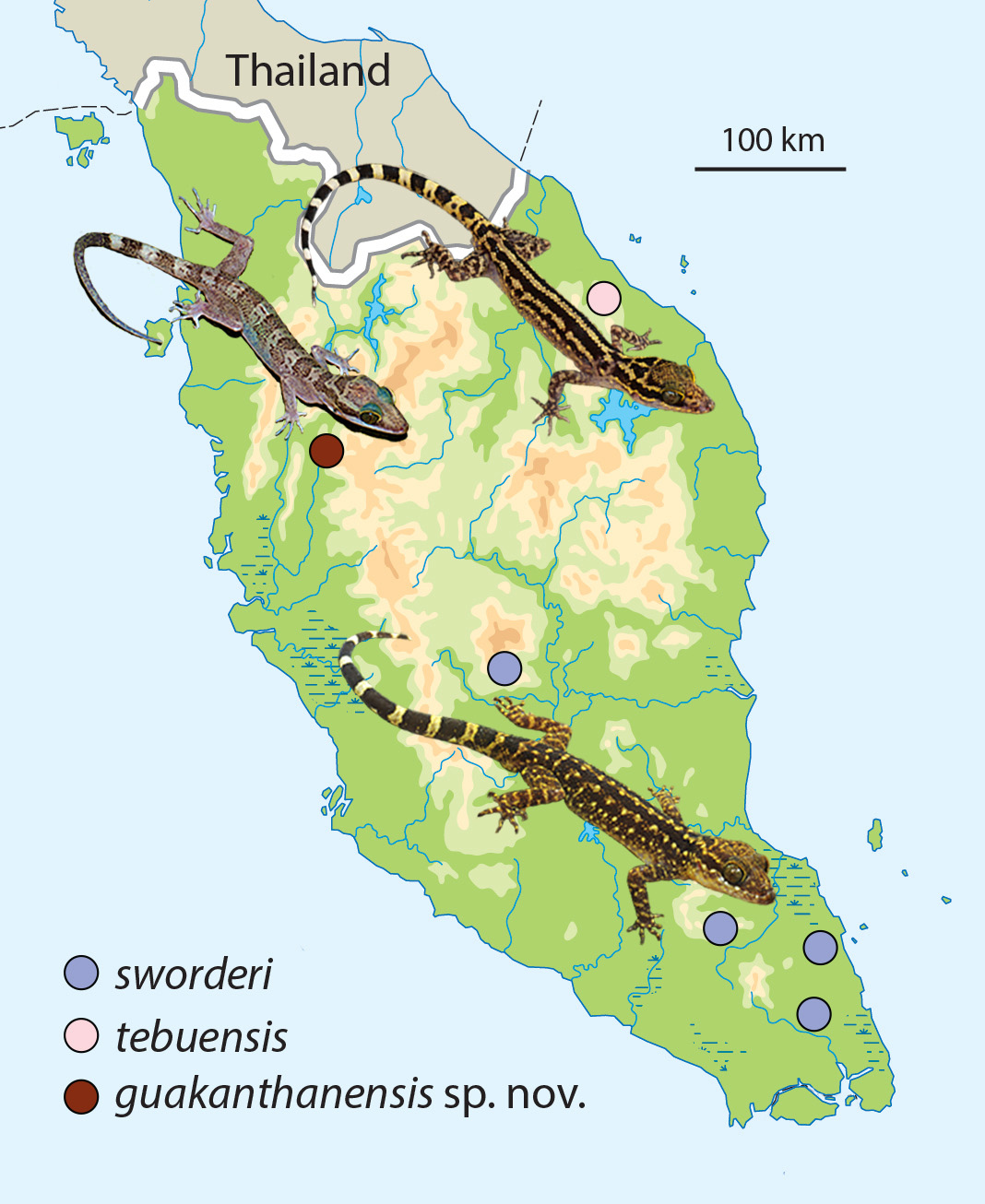


TABLE 5. Diagnostic characters (in bold) separating the species of the C. sworderi complex from Cyrtodactylus guakanthanensis sp. nov.
| guakanthanensis sp. nov. tebuensis sworderi | quadrivirgatus | |
|---|---|---|
| Supralabials | 9,10 11–13 10 – 13 | 8 – 11 |
| infralaials | 7,8 8 – 10 10 | 8 – 11 |
| No. of paravertebral tubercles | 32 – 34 32 – 39 36–38 | 32 – 39 |
| No. of ventral scales | 37 – 44 43 – 51 40 – 49 | 28 – 40 |
| Postfemoral scale transition abrupt No. of 4th toe lamella | yes yes no 19 – 21 17 – 21 16 – 20 | no 18 – 23 |
| Precloacal and /or femoral pores | absent absent present | variable |
| No. of precloacal pores | 0 0 5–9 | 0–12 |
| No. of enlarged femorocloacal scales | 36 – 41 31 – 37 0 | 31 – 44 |
| Light caudal bands | wide narrow narrow | wide |
| No. light bands on original tail | 9 10–14 9 – 15 | 9 – 15 |
| Dorsal pattern | banded Striped/spotted spotted | variable |
TABLE 6. Morphological characters of type series of Cyrtodactylus guakanthanensis sp. nov. / = data unavailable.
| LSUHC | LSUHC | LSUHC | LSUHC | LSUHC | LSUHC |
|---|---|---|---|---|---|
| 11321 | 11322 | 11323 | 11325 | 11326 | 11339 |
| paratype | holotype | paratype | paratype | paratype | paratype |
| sex f | m | m | m | m | m |
| supralabials 10 | 9 | 10 | 9 | 10 | 10 |
| infralabials 7 | 8 | 8 | 8 | 8 | 7 |
| No. of paravertebral tubercles 34 | 32 | 33 | 32 | 33 | 33 |
| No. of ventral scales 44 | 37 | 42 | 42 | 41 | 38 |
| 4th toe lamellae 21 | 20 | 20 | 21 | 21 | 19 |
| Preclacal pores present (1) or absent (0) 0 | 0 | 0 | 0 | 0 | 0 |
| No. of enlarged femorocloacal scales 37 | 39 | 36 | 41 | 39 | 36 |
| No. of light bands on original tail / | / | / | 9 | / | / |
| No. of body bands 5 | 4 | 5 | 5 | 5 | 5 |
| SVL 82.2 | 79.7 | 79.8 | 78.2 | 77.7 | 81.8 |
| TL 91.5 | 78.3 | 83.4 | 97.9 | 85.7 | 89.2 |
| TW 7 | 7.4 | 7.3 | 7.1 | 7.2 | 6.8 |
| FL 12.9 | 13.3 | 13 | 13.2 | 12.6 | 12.7 |
| TBL 14.7 | 14.7 | 15.5 | 15.4 | 14.5 | 16 |
| AG 36.2 | 33.3 | 34.1 | 35 | 34.5 | 37.8 |
| HL 23 | 22.3 | 22.8 | 22 | 21.3 | 23 |
| HW 15.1 | 14.5 | 15.1 | 15 | 13.3 | 15.1 |
| HD 9.5 | 9 | 9.6 | 9.5 | 8.3 | 9.7 |
| DE 5.5 | 5 | 5.5 | 5.2 | 4.7 | 5.6 |
| EE 6.5 | 6.3 | 6.2 | 6.3 | 5.8 | 6.5 |
| ES 9.9 | 9.8 | 10 | 10.1 | 10.1 | 10.2 |
| EN 7.8 | 7.5 | 7.6 | 7.6 | 7.5 | 7.8 |
| IO 4.2 | 3.9 | 4.2 | 3.8 | 3.8 | 4 |
| EL 1.7 | 1.9 | 1.7 | 2.3 | 1.5 | 1.9 |
| IN 2.5 | 2.6 | 2.5 | 2.4 | 2.4 | 2.4 |
| Discussion |
| LSUHC |
La Sierra University, Herpetological Collection |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |