
Deutonura gibbosa, Porco, David, Bedos, Anne & Deharveng, Louis, 2010
|
publication ID |
https://doi.org/10.5281/zenodo.198510 |
|
DOI |
https://doi.org/10.5281/zenodo.5661841 |
|
persistent identifier |
https://treatment.plazi.org/id/C16E0551-217C-FFA6-C1A0-FB497D294201 |
|
treatment provided by |
Plazi |
|
scientific name |
Deutonura gibbosa |
| status |
sp. nov. |
Deutonura gibbosa sp. nov.
Figs 1 View FIGURE 1 , 2–6 View FIGURES 2 – 6 , 8 View FIGURES 7 – 8 ; Tab. 1 View TABLE 1
Type material. Holotype male and 90 paratypes, France, Isère, Montaud, along the road from Saint-Quentinsur-Isère to Montaud. 3.xi.2007, mixed broadleaf forest, litter, berlese. L. Deharveng & A. Bedos leg. (38– 036) (Alt. 500 m, X 5.557634, Y 45.276590). Deposited in MNHN (holotype, 80 paratypes) and in ZIWU (10 paratypes).
Other material. France. Ain: Anglefort, le Grand Colombier, 2 ex, eastern slopes, 13.v.2001, beech forest, litter, berlese, L. Deharveng & A. Bedos leg. (01–012); Culoz, le Grand Colombier, 4 ex., 13.v.2001, beech-fir forest, litter, berlese, L. Deharveng & A. Bedos (01–013).
Haute-Savoie: Cordon, 2 ex., 1xii.1977, beech forest, litter, L. Deharveng (74–160); Dingy-Saint-Clair, Nanoir, 3 ex., 10.v.1983, beech and oak forest, litter, L. Deharveng leg. (74–184); Les Clefs, La Tournette, 2 ex., 27.ix.1976, beech forest, litter, L. Deharveng leg. (74–151); Saint-Jorioz, montagne du Semnoz, 2 ex., 23.vii.1973, small shaft with névé, in daylight, (74–061); ibid., crête du Tertère, 9 ex., 20.viii.1979, under Sorbus , litter, L. Deharveng leg. (74–169); ibid., 14 ex., 1.ix.1980, on lapiez, spruce forest, litter, L. Deharveng leg. (74–170); Sillingy, Chaumontet, Montagne d'Age, at a small spring, 3 ex., 23.vii.1974, in moss, L. Deharveng leg. (74–148); Vallorcine, Col des Montets, 3 ex., 1.ix.1980, larch and Rhododendron forest, litter, L. Deharveng leg. (74–171).
Hautes-Alpes: Aspres-sur-Buech, 8 ex., 4.xi.2007, beech forest, litter, berlese, L. Deharveng & A. Bedos leg. (05–035); La Faurie, Bois de Longeagnes, 1 ex., 13.v.1981, fir forest, litter, J. Monbayé leg. (05–006); Le Monetier-les-Bains, Le Casset, vallon du Petit Tabuc, 1 ex., 19.vii.1997, larch litter, berlese, F. Brouquisse leg. (05–027).
Isère: La Balme-les-Grottes, above the village, 2 ex., 13.v.2001, oak forest, litter, berlese, L. Deharveng & A. Bedos leg. (38–026); Le Sappey-en-Chartreuse, Col de Porte, 12 ex., 5.viii.1977, L. Deharveng leg. (38– 038); Saint-Laurent-du-Pont, Pont Saint Bruno, 13 ex., 3.xi.2007, litter, berlese, L. Deharveng & A. Bedos leg. (38–028); Saint-Pierre-de-Chartreuse, Pont des Cottaves, 1 ex., 3.xi.2007, spruce forest, litter, berlese, L. Deharveng & A. Bedos leg. (38–032).
Savoie: Aillon-le-Jeune, Col des Près, 20 ex., 1.v.1982, beech forest, litter, berlese. L. Deharveng leg. (73–038); Bourg-Saint-Maurice, Beaufortin, vallon de la Neuva, 1 ex., 24.ix.1976, lichen, moss on rock, soil in the alpine zone, L. Deharveng leg. (73–021); Saint-Christophe, near grotte de Fontaine Noire, 1 ex., 12.vii.1984, beech forest, litter, berlese, L. Deharveng leg. (73–053).
Italy. Liguria: Imperia: Cossio di Arroscia, near Ponte di Nava, 2 ex., 15.xii.1982, beech forest, litter, L. Deharveng leg. (ITA–015).
Lombardia: Bergamo: Oneta, Valle de Seriana, Monte Alben, near Zambla, 31 ex., 14.viii.1997, beech mixed with a few spruce, litter, L. Deharveng & A. Bedos leg. (ITA–097, –098, –099); Brescia: Col de Vivione, northen slope, 42 ex., 14.viii.1997, Pinus uncinata , birch and Rhododendron , litter, berlese, L. Deharveng & A. Bedos leg. (ITA–100); Brescia, foot of Col de Vivione, Valle Paisco, 16 ex., 14.viii.1997, Castanea , litter, berlese, L. Deharveng & A. Bedos leg. (ITA–102).
Piemonte: Cuneo: Garessio, Capello, road to Pamparato, 23 ex., 15.xii.1982, beech forest, litter, L. Deharveng leg. (ITA–017); Pamparato, Valcasotto, 6 ex., 2.x.1979, beech forest, M. Curti leg. (ITA–002); Ponte di Nava, below Bricco, 120 ex., 15.xii.1982, chestnut forest, litter, L. Deharveng leg. (ITA–013).
Trentino-Alto Adige: Bolzano: above Ora, Castel Vetere hill, 1 ex., 15.viii.1997, oak forest, litter, berlese, L. Deharveng & A. Bedos leg. (ITA–121); Trento: Sdruzzina, Passo Fittanze della Sega, 27 ex., 12.ix.2002, beech forest, litter, berlese, L. Deharveng & A. Bedos leg. (ITA–139).
Veneto: Belluno: Cortina d'Ampezzo, Falcade , east of Passo San Pellegrino, 1 ex., 15.viii.1997, beech, by hand, L. Deharveng & A. Bedos leg. (ITA–124); Belluno: Bosco de Cansiglio, near Spert, 1 ex., 16.viii.1997, beech, litter, berlese, L. Deharveng & A. Bedos leg. (ITA–135).
Slovenia. Lokev: Divaca, Vilenica cave, 5 ex., 27.vi.1981, E. Christian leg. (SLN–Christ001).
Switzerland. Vaud: Bex, Solalex, massif des Diablerets, 1 ex., 1.viii.1982, fir and spruce forest, litter, L. Deharveng leg. ( SUI –004).
Etymology. The species name gibbosa is a latine word meaning humped, in reference to the prominent Di tubercle of Abd. V of the new species.
Description. Length: 1–2 mm. Color: deep blue to light blue. Habitus usual for the genus, rather parallel. Abd. VI completely hidden under Abd. V, deeply bilobed. All tubercles well developed on tergites, indicated by reticulations and tertiary granules without papillae, but with strong secondary granules especially on Abd. V. All dorsal chaetae integrated in tubercles (no free chaetae). Dorsal ordinary chaetae of four types: long macrochaetae, short macrochaetae, mesochaetae and microchaetae. Long macrochaetae long, robust, thick, finely and sparsely rugose, sheathed, rounded-ogival at the apex, except the most lateral ones pointed apically. Short macrochaetae similar to long macrochaetae, but shorter. Mesochaetae similar to ventral chaetae, thin and acuminate, short, smooth. Microchaetae morphologically similar to mesochaetae but very short, limited to Oca in some specimens. S-chaetae of tergites thin and long but much shorter than neighbour long macrochaetae (slightly less than 1/2 as long as nearby macrochaeta on Abd. V). S-chaetae of thoracic tubercles De antero-internal to De1 and equally distant to De1 and De2.
Antennae typical of the genus. Ant. IV with S-chaetae thickened, S1 and S2 thinner than others; apical vesicle trilobed. Mouthparts reduced, mandible thin and tridentate, maxilla styliform. Labrum with ventrodistal sclerification rounded and truncated apically, with several minute denticles on apical edge. Labium chaetotaxy as in Fig. 4 View FIGURES 2 – 6 , with chaetae A and C subequal, three times as long as D. Eyes black, 2+2. Head chaetotaxy and tubercles as in Fig. 2 View FIGURES 2 – 6 and Tab. 1 View TABLE 1 . Tubercle Af with elementary tubercles DE and EE present, two elementary tubercles between chaetae A and no granular plate between chaetae A and B.
Tergite chaetotaxy and tubercles as in Fig. 2 View FIGURES 2 – 6 and Tab. 1 View TABLE 1 . Tubercles Di of Abd. V fused into a large, strongly bilobed axial tubercle overhanging Abd. VI. Chaetae Di1, Di2 and Di 3 in elongated triangle with Di1–Di2 about 2.5 times shorter than Di1–Di3. Di1: very long, bent macrochaeta, Di 2: long macrochaeta 2/ 3–3/4 as long as Di1, and Di3: short macrochaeta 3 times shorter than Di 2 in adult. Tergite pseudopora (arranged as 0,1,1/1,1,1,1,?1,0 per half tergite) and muscular insertions shown on Fig. 2 View FIGURES 2 – 6 .
Sternite chaetotaxy as in Fig. 6 View FIGURES 2 – 6 and Tab. 1 View TABLE 1 .
Leg chaetotaxy as in Fig. 5 View FIGURES 2 – 6 and Tab. 1 View TABLE 1 . No ventral tenent hairs on tibiotarsi, B4 and B5 moderately long, not or slightly overpassing apex of tibiotarsi. Claw basally granulated, without inner tooth.
Cephalic chaetotaxy
Mc Ocp me or mi Oca Di + De + 4 ML Di1,De1 Mc Di2,De2 DL + 6 ML DL1,DL5
Mc DL2,DL3,DL4,DL6
L + So + 8-(9) ML So1,L1,L4 Mc or me L2 me (L3),So3 to 6 Vi of head 6
Ve of head 10 or 11 Labrum 4/2,4 Labium 10, 0x Ant. I 7
Ant. II 12
Ant. III 17 +5s Ant. IV dorsally 12 mou+or+i+8s
Postcephalic chaetotaxy * Vel present, sometimes as an uneven mesochaeta Male without modified chaetae ventrally.
Remarks. Deharveng (1979) recognized three forms in the subspecies Deutonura plena plena ( Stach, 1951) , that was subsequently renamed D. deficiens sylvatica Deharveng, 1982 after it was clearly shown that D. plena was actually a species of northeastern Europe. The type material from Ardèche near the Rhône valley in France corresponds to the form B characterized by a pigmented body and the presence of 8 chaetae L on Abd. IV. Specimens from Alps cited by Deharveng (1979), which are now placed in the new species D. gibbosa , also correspond to the form B. Indeed, D. gibbosa sp. nov. shares most chaetotaxic and tuberculisation features with the form B of D. deficiens sylvatica , widespread in southwestern France and northwest of the Iberian peninsula, and recorded from Sardinia by Dallai (1983). The new species is readily separated from it by the morphology of the dorso-internal tubercle of Abd. V, conspicuously bilobed, larger and more elongate, hiding most of Abd. VI in dorsal view. This development of dorso-internal tubercle of Abd. V is associated to a chaetotaxic re-arrangement, with Di1 and Di2 shift backwards in the new species compared to D. d. sylvatica ( Figs. 7, 8 View FIGURES 7 – 8 ). This difference holds for all specimens of the form B examined from the cited regions, including those from Sardinia. The ratio of chaetae Di on Abd. V is also different, with Di2 2/3–3/4 as long as Di1 and almost 3 times as long as Di3, versus Di2 about half as long as Di1 or less, and about 2 times as long as Di 3 in types of D. d. sylvatica .
Distribution and ecology. With a distribution ranging from southwest to eastern Alps and southern Jura, D. gibbosa sp. nov. is one of the most widespread species of the genus. It replaces D. deficiens sylvatica to the east of the Rhône valley ( Fig. 1 View FIGURE 1 ). Given the frequency of the new species in the Alps, it is surprising that the species seems to have been overlooked by previous authors. For instance, none of the Deutonura cited from Austria by Christian (1987) could correspond to D. gibbosa sp. nov. It might be that the species, while present in the whole massif, may be replaced in some large areas like Austria by other Deutonura species, as it seems to be the case in the most southwestern regions of the Alps.
D. gibbosa sp. nov. is frequent in forest litter and rotten wood of any kind of forest, even disturbed, from 300 to 1500 m a.s.l.
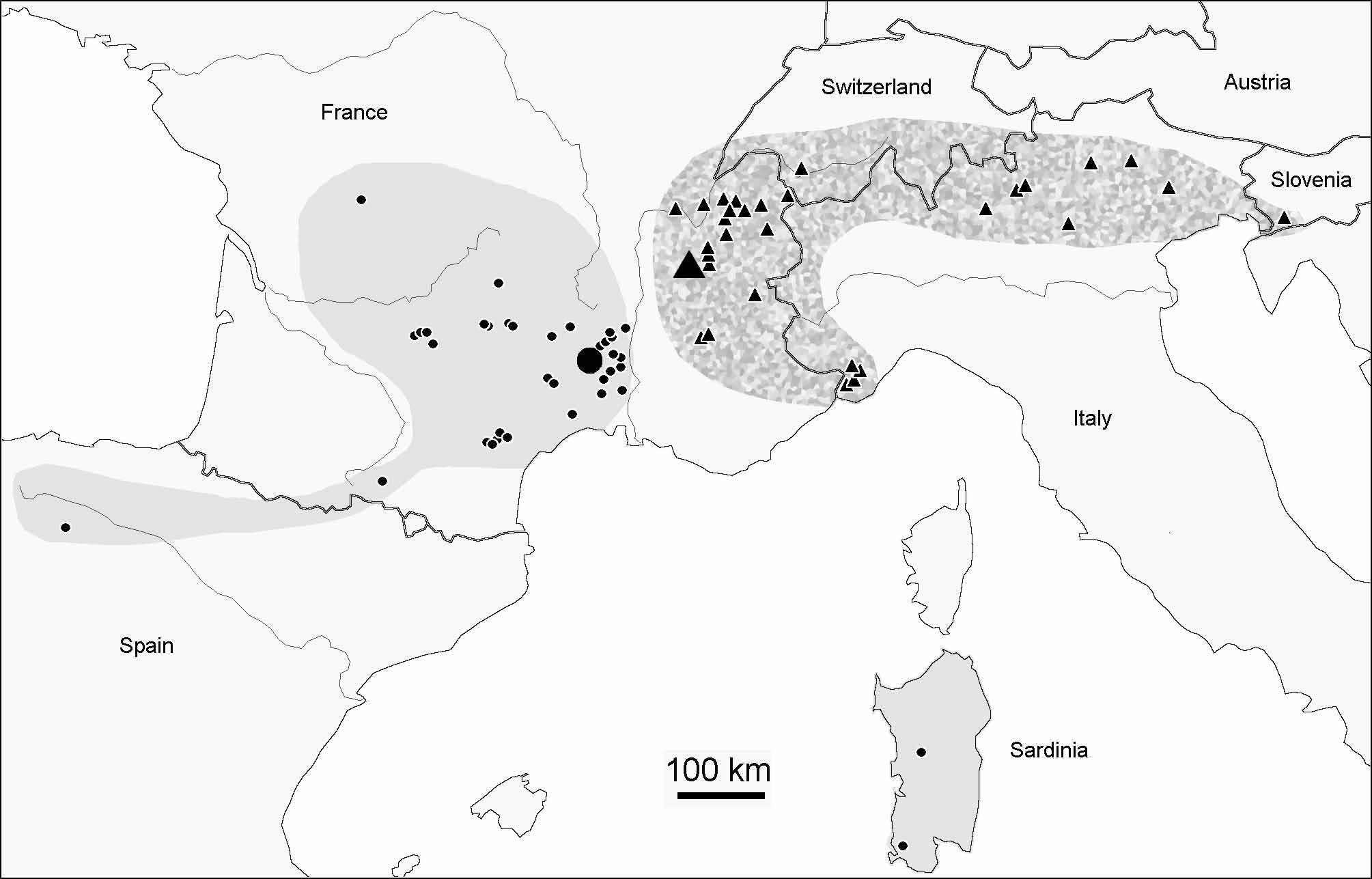
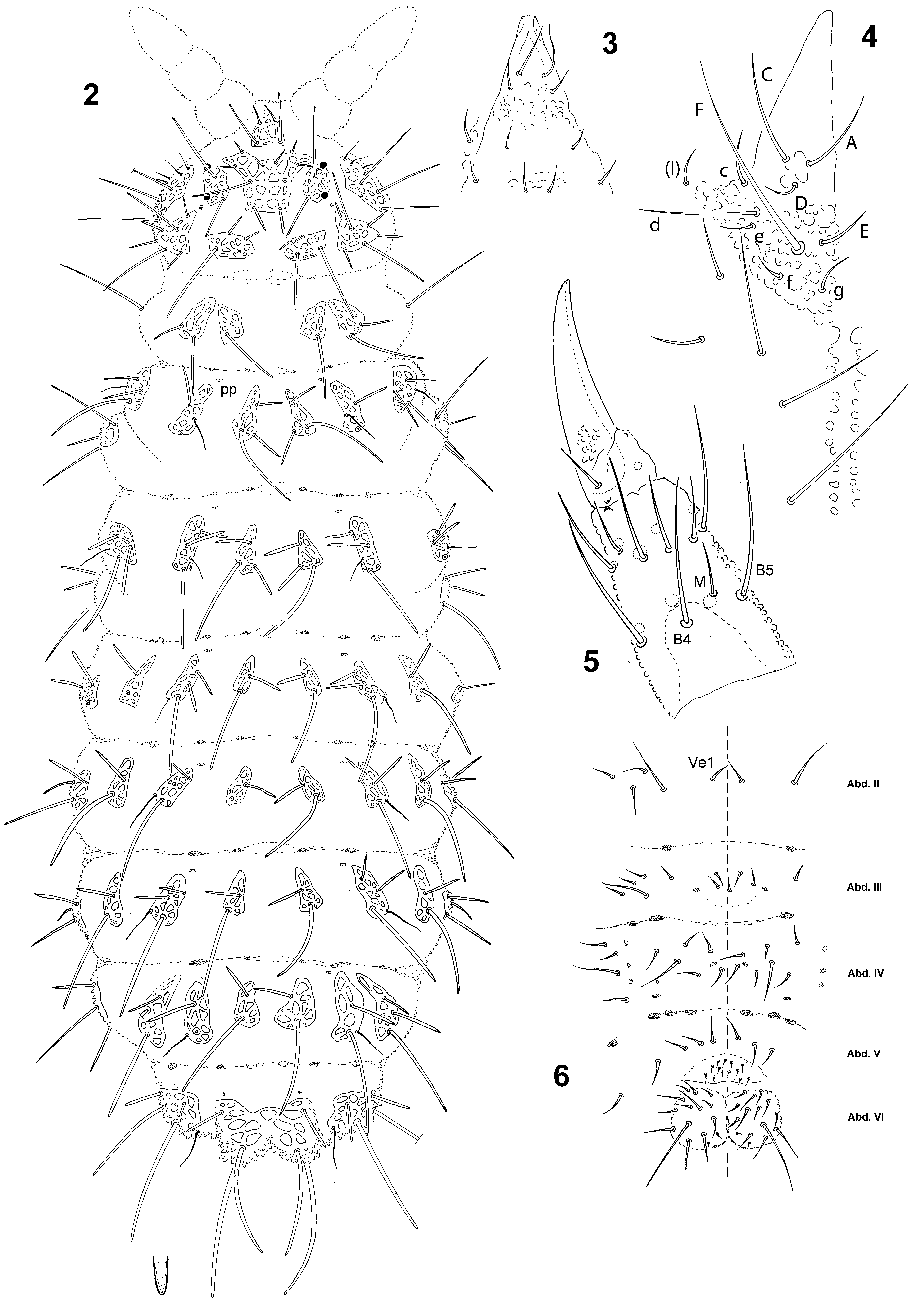
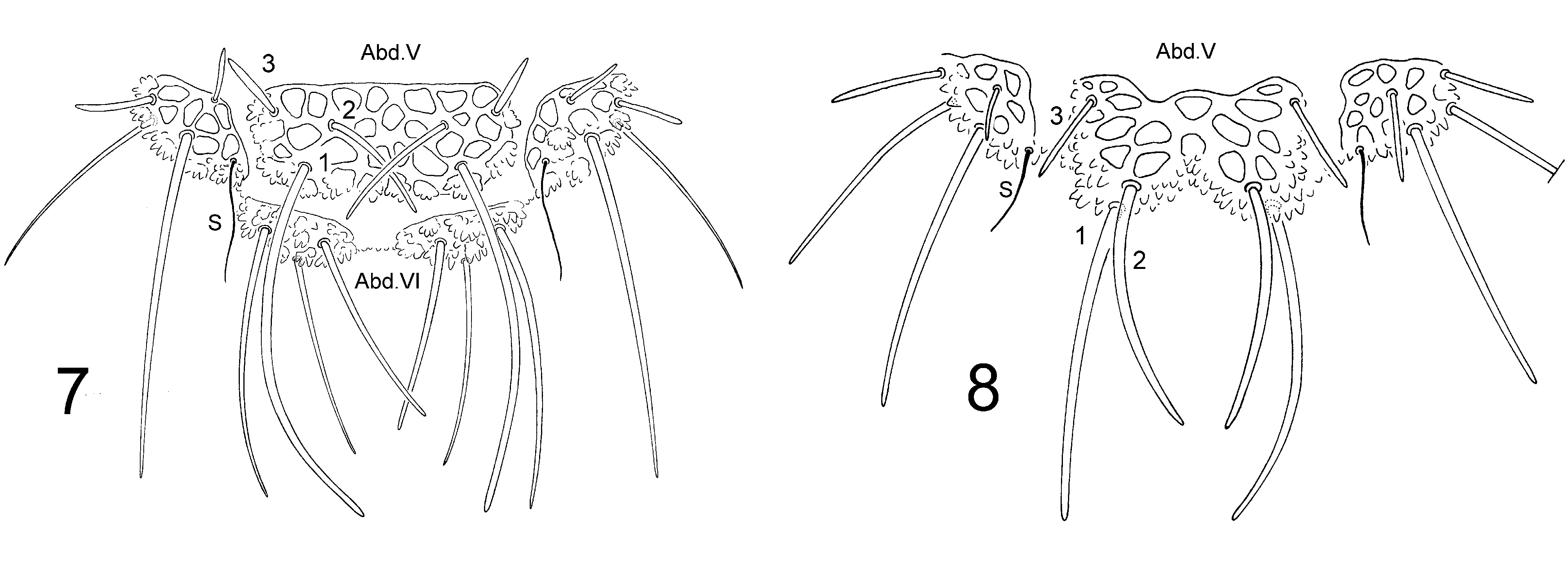
TABLE 1. Chaetotaxy of Deutonura gibbosa sp. nov.
| Group of chaetae | Tubercle Number of chaetae | Type of chaetae | Name of chaetae |
|---|---|---|---|
| Cl | + 4 | ML Mc | F G |
| Af | + 10 | ML | B |
| Oc | + 3 | Mc ML | A,C,D,E Ocm |
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
SubFamily |
Neanurinae |
|
Genus |