
Callichimaera perplexa, Luque & Feldmann & Vernygora & Schweitzer & Cameron & Kerr & Vega & Duque & Strange & Palmer & Jaramillo, 2019
|
publication ID |
https://doi.org/ 10.1126/sciadv.aav3875 |
|
publication LSID |
lsid:zoobank.org:pub:74A09A7E-A192-4481-A37E-A1023EBB300A |
|
DOI |
https://doi.org/10.5281/zenodo.5902754 |
|
persistent identifier |
https://treatment.plazi.org/id/C32E2E2B-6D38-FFF5-0C69-FF43DDCCFC5B |
|
treatment provided by |
Diego |
|
scientific name |
Callichimaera perplexa |
| status |
gen. et sp. nov. |
Callichimaera perplexa gen. et sp. nov.
LSID. urn:lsid:zoobank.org:act:CD4585D1-B198-45E6-8485-45F94167BDEE
LSID. urn:lsid:zoobank.org:act:650E5046-C4FC-4485-A3B5-254DE785F80B
Etymology. The section, superfamily, family, and generic names are derived from the Greek prefix calli- “kalos” (beautiful), alluding to its exceptional preservation, and Chimera, the fabulous mythological beastcommonlyrepresented ascomposedof parts of different animals such as the lion, goat, and snake, alluding to its startling combination of traits present in separate higher decapod taxa, e.g., eubrachyurans, podotreme brachyurans, anomurans, and some macrurans. Thespecific epithet derives from the Latin “perplexus,” referring to its puzzling anatomy and phylogenetic affinities. The gender is feminine.
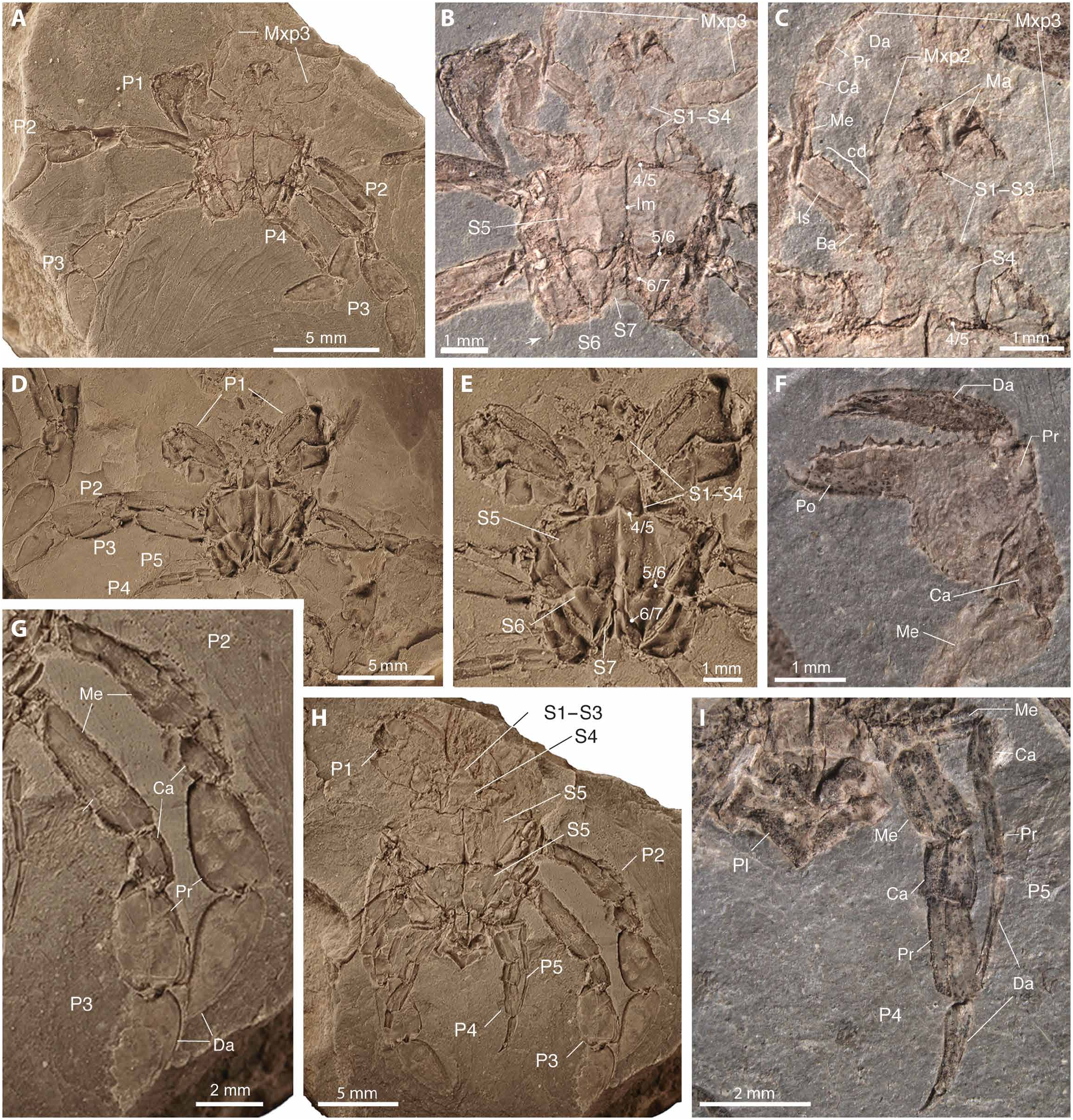
Diagnosis. Smallcrabs (carapace width, <10 mm; carapace length, <16 mm) that have a carapace longer than wide, fusiform, with distinct cervical and branchiocardiac grooves, and bearing axial longitudinal ridge and postfrontal ridges. Sternites 1 to 4 are visible ventrally; sternites 4 to 7 are unfused, with sutures distinct, and axially sulcate by linea media; all sternites are unique in shape and size; sternite 5 is very wide; suture 5/6 is complete, irregular, and sinuous, lacking true sterno-pleonal cavity; thoracic gonopores are not recognized in males or females. The pleon is symmetrical, sexually dimorphic, narrower in males than females, and in both sexes narrower than sternite 6. Pleonal somites are not fused, lacking articulating rings and bearing dorsal median tubercle; pleonites 1 to 3 are exposed subdorsally, and lacking pleonal, sternal, or appendicular locking mechanisms; uropods or uropodal plates are absent. The rostrumis bifid; first andsecond antennaeare short, between the eyes; eyes are very large—the corneais strongly dilated, subglobular, bearing mostly hexagonal facets, and a short ocular peduncle, lacking orbits, orbital fissures, or any protective structure; third maxillipeds are pediform, elongate, with “crista dentata”; lengths of the ischium and merus are slightly longer than the length of the palp, and the merus is positioned far back from anterior of carapace or basal antennal segments. Chelipeds (claws) are isochelous, the manus is stout, with fixed finger deflected ~90°; pereopods (legs P2 and P3) are large and wide, with propodus and dactylus flattened and paddlelike; P4 and P5 are short and narrow, with the dorsal longitudinal keel, lacking spines, notsubchelate or modified tocarry objects, and neither flattened nor paddle-like; P5 is the smallest, well developed but reduced, and carried subdorsally [modified from ( 21)].
Description. Seethe Supplementary Materialsforadetaileddescription of C. perplexa gen. et sp. nov.
Holotype. IGM p881215 , specimen preserved in ventral view ( Fig. 1 View Fig , Ato C), deposited in the paleontological collections of the Colombian Geological Survey, Diagonal 53 #34-53, Bogotá DC, Colombia. Carapace length is 8.5 mm, and carapace width is 5.2 mm.
Additional material. Colombian paratypes IGM p881184 to IGM p881214 and IGM p881216 to IGMp881221 are deposited in the paleontological collections of the Colombian Geological Survey; paratypes MUN-STRI 27044-01 to MUN-STRI 27044-010 and MUN-STRI 27045-01 to MUN-STRI 27045-020 aredepositedinthe Mapuka Museum of Universidad del Norte, Barranquilla, Colombia. Additional nontype materials from the United States, specimens USNM 605049 View Materials to USNM 605056 View Materials , aredepositedinthe Paleobiology Collections of the National Museum of Natural History, Smithsonian Institution, Washington, DC, USA .
Measurements. The range of measurements is as follows: Holotypehas acarapacelengthof 8.5 mmandacarapacewidthof 5.2 mm, the smallestparatype IGM p881220 has a carapace length of 6.6 mm and a carapace width of 3.8 mm, and the largest paratype MUN-STRI 27045-015 has a carapace length of 15.1 mm and a carapace width of 9.6 mm.
Type locality, age, and horizon. Churuvita Group, upper Cenomanian to lower Turonian (~95 to 90 Ma ago), Pesca, Boyacá, Colombia (data files S1 and S2), from carapace- and appendagerich surfaces. Other specimens are from the Frontier Formation, lower-middle Turonian (~90 Ma ago), WY, USA. Seethe Supplementary Materials for a detailed description of the geological, geographical, and paleontological context.
Systematic remarks
Althoughmolecularandmorphological phylogeneticsbringpowerful toolstothestudyof relatednessatthe genotypicandphenotypic levels, the fossil record provides a unique view into the origins of such relatedness by revealing a past morphological diversity otherwise inaccessible. Furthermore, fossils are pivotal for understanding the evolutionof keytraits and provide geographic and chronologic data critical to the calibration of nodes of interest.
Weconsider Callichimaeroidasectionnov.atruecrabor Brachyura insteadof Anomurabasedon thefollowing: (i) itsshort firstandsecond antennaebetween the eyes, (ii) a symmetric, sexually dimorphic pleon, (iii) the absence of articulating rings between pleonites, (iv) a reduced telson, (v) complete absence of uropods or uropodal plates, (vi) the presenceof modifiedmalepleopods 1 and 2 ashighly sclerotizedgonopods but lacking pleopods 3 to 5, while the female bears pleopods 2 to 5, (vii) third maxilliped with well-defined ischium and merus, (viii) the presence of only one pair of chelae or claws (pereopod 1), thus pereopods 2 to 5 are achelate, and (ix) P5 is well developed, visible in dorsal view, and neither subchelate nor modified for carrying or grasping ( Figs. 1 View Fig to 3). However, a precise phylogenetic placement of Callichimaera within Brachyuraisproblematicbecauseof its “chimeric” nature, theunknownmoltinglinea, andpossessionof multipledistinctive characters typicalof several fossil and extant Brachyura and Anomura clades but not collectively seen in any one taxon. These characters include a lobster/raninid-like elongate carapace, pediform maxillipeds with a crista dentata, spanner-like chelipeds, large paddle-like legs P2 and P3, thedissimilarshape andsizeof itssternites, asymmetricalpleon lacking uropods or uropodal plates, a dorsally keeled carapace, and largeeyes lacking true orbits and orbital fissures ( Figs. 1 View Fig to 3).
Phylogenetic remarks
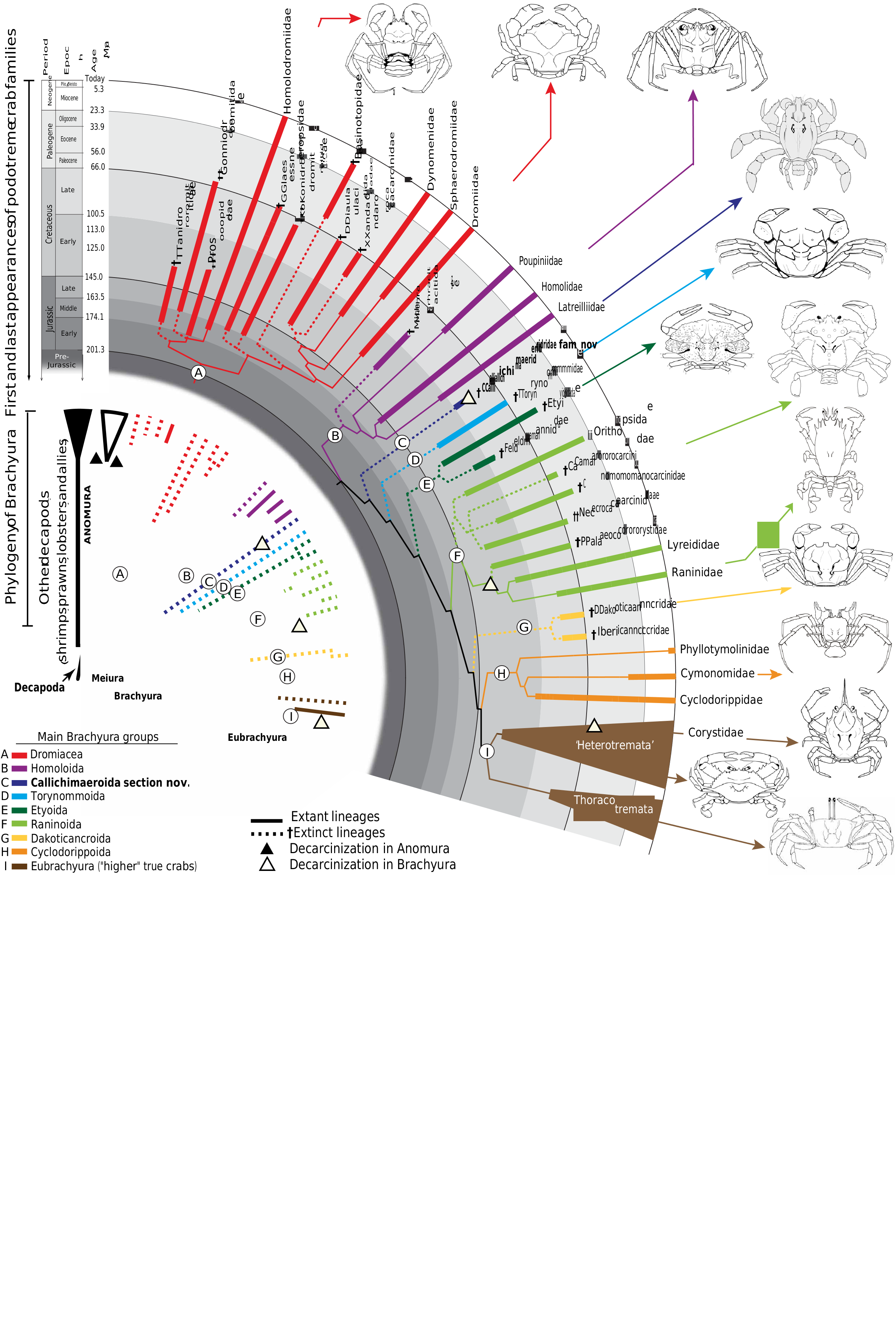
Both molecular and morphological phylogenetic studies have recovered Brachyura, or true crabs, as a monophyletic and sister group to Anomura (false crabs and allies) ( 2, 10, 11, 22). Yet, phylogenetic relationships within Brachyura remain unsettled largely because of the lack of early, intermediate body forms. Results of our Maximum Parsimony (MP) and Maximum Likelihood (ML) analyses largely agree on the arrangement of ingroup taxa, including placement of Callichimaera as an independent lineage branching off before the extinct highertaxa †Torynommoida and †Etyoida ( Fig. 5 View Fig and figs. S5, S6A, and S7). Although Callichimaera isalsorecoveredin thesame positionunder Implied Weight MP (IWMP) with moderate K values (K = 6) (fig. S6C), underlow K values (K = 3), Callichimaera is pulled into a clade with Palaeocorystidae and Raninidae + Lyreididaeasaresultof along-branchattractionduetothehomoplastic characters shared among decarcinized crabs and, therefore, artificially inflating the fit value of the tree (fig. S6B). Similarly, under IWMP high K values (K = 12), Callichimaera is pulled into a clade with Orithopsidae and Necrocarcinidae + Cenomanocarcinidae as a result of a long-branch attraction due to the convergentcarapaces ornamentedwithseveral dorsallongitudinalkeelsorcarina (fig. S6D).
The Bayesian Inference (BI) consensus tree presents the least resolvedtopologywithseveralmajor brachyuranlineagescollapsedinto a polytomy, yet showsa clearparaphyletic podotremegrade (fig. S8). Discrepancies between topologiesrecovered by probabilistic methods (ML and BI) under the same model of evolution (Mk) are not unusualconsidering differences inthecriteria for selectingthe best ML and BI topologies. Since BI uses marginal likelihoods to select the optimal topology (as opposed to the joined likelihood in the ML estimation), it is more sensitive to inconsistencies in a dataset.
Although BI has been shown to produce accurate topologies whendealing with morphological data ( 23, 24), themethod remains sensitive to the consistencyof the phylogenetic signal present in the dataset, selection of priors, and heterogeneity of evolutionary rates across different lineages on a tree. The lack of resolution in our BI consensus topology is best explained by high topological disparity in the posterior sample of trees, caused by multiple cases of convergent traits and disparity inevolutionary rates (pedomorphosis, high phenotypic plasticity, etc.) across lineages. In light of the ongoing debate over relative performance of parsimony and model-based approaches when analyzing morphological data ( 23 – 29), we present results of phylogenetic analyses under multiple optimality criteria, which agree in the placement of Callichimaeridae as a new, independent lineage.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.