
Kaloula nonggangensis, Mo, Yunming, Zhang, Wei, Zhou, Shichu, Chen, Tianbo, Tang, Huaxing, Meng, Yuanjun & Chen, Weicai, 2013
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3710.2.3 |
|
publication LSID |
lsid:zoobank.org:pub:7E5079D2-5655-4069-8728-7302191EF180 |
|
DOI |
https://doi.org/10.5281/zenodo.5661176 |
|
persistent identifier |
https://treatment.plazi.org/id/C45587C4-FFB4-FFD7-17D7-FF2D0478FE9B |
|
treatment provided by |
Plazi |
|
scientific name |
Kaloula nonggangensis |
| status |
sp. nov. |
Kaloula nonggangensis View in CoL sp. nov.
Holotype. NHMG1106036, adult male, from the Nonggang National Nature Reserve, Southern Guangxi Province, China (22.4522° N, 106.9354° E; altitude: 186 m a.s.l.), collected on June 28, 2011 by Weicai Chen, Yunming Mo ( Figure 2 View FIGURE 2 A, B, C).
Paratypes. NHMG1106030–35, NHMG1106037–41, adult males collected at the same place of the locality by Weicai Chen and Yunming Mo on June 29, 2011. NHMG1108035, adult female, NHMG1108036– 41 adult males, collected at the near locality of the holotype (22.4809° N, 106.9017° E; altitude: 167 m a.s.l.) by Yunming Mo, Wei Liao and Zhuqiu Song on August 11, 2011.
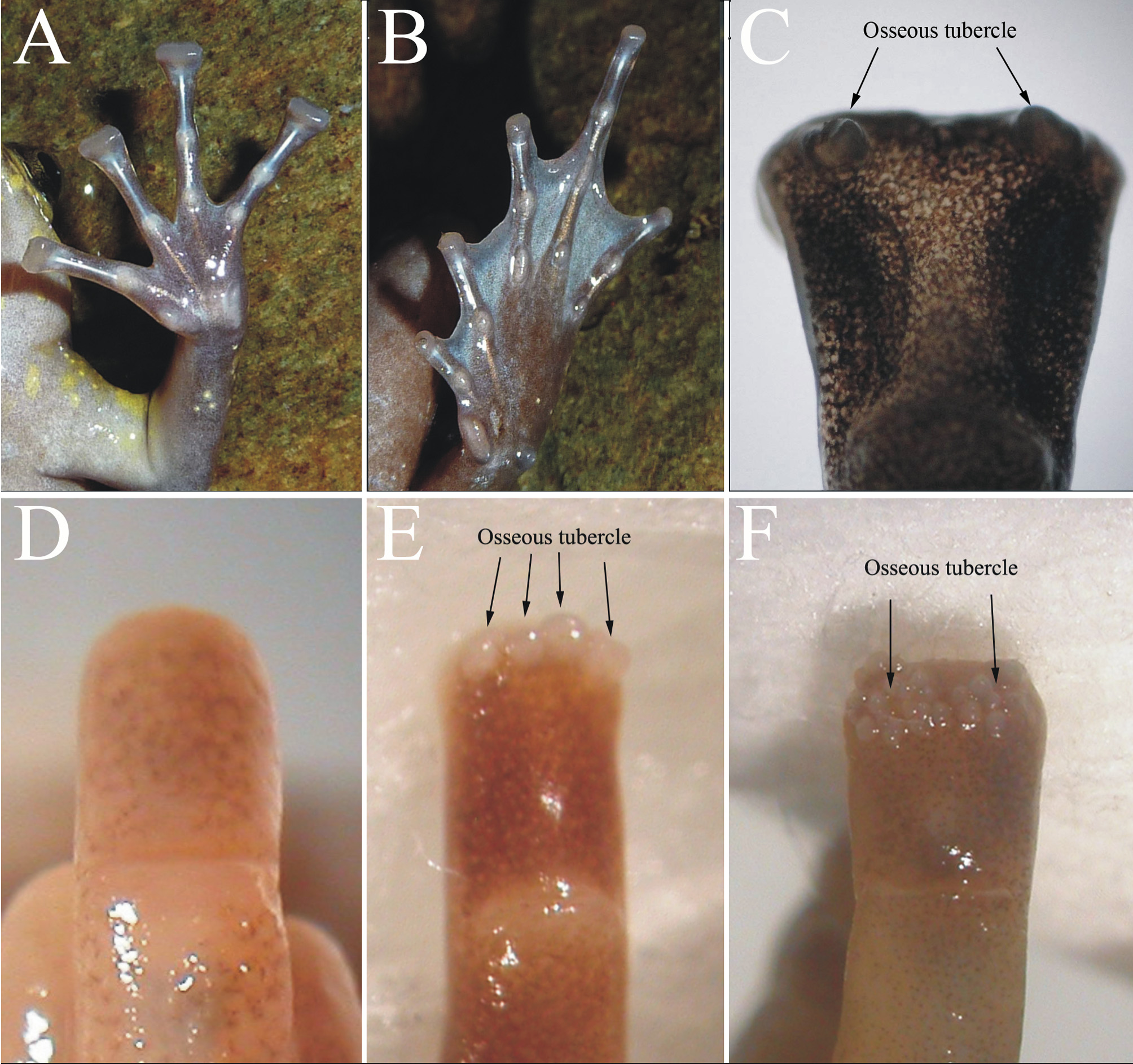
Diagnosis. Assigned to the genus Kaloula on the basis of the following: medium size (41.4–52.7 mm in 18 adult male, 52.2 mm in one adult female), smooth or slightly rough olive dorsum with irregular dark-green marks and brown spots ( Figure 2 View FIGURE 2 A, B); tips of the fingers dilate and truncated ( Figure 3 View FIGURE 3 A); nearly full webbing on toes in males and reduced webbing in females ( Figure 3 View FIGURE 3 B); two side protuberant osseous tubercles on the upper surface of the tips of fingers in male ( Figure 3 View FIGURE 3 C); chest beige with small lemon-colored spots in male ( Figure 2 View FIGURE 2 C); ventral epidermal adhesive gland occupies chest and venter; larvae lacking a keratinized jaw sheath and labial teeth. K. nonggangensis sp. nov. is distinguished from it congeners by a combination of (1) medium size (SVL ranging 41.4–52.7 mm in males), (2) smooth or slightly rough olive dorsum with irregular dark-green marks and brown spots, (3) tips of the fingers dilate and truncated, (4) chest beige with small lemon-colored spots in male, and (5) male with two side protuberant osseous tubercles on the upper surface of the tips of fingers.
Description of holotype. Adult male; SVL 47.5 mm; habitus rotund, body rounded; head proportionally small, length 25% of SVL, 76% of head width; snout slightly pointed beyond lower jaw, its tip round in dorsal aspect and in lateral aspect; eyes protruding laterally beyond silhouette of head in dorsal aspect, protruding very markedly beyond dorsal surface of head in lateral aspect; pupil circular; interorbital region flat with some small tubercles; snout less than half of the head length; upper eyelid width slightly smaller than interorbital distance, and much smaller than snout length; eye diameter (4.63 mm) almost equal to snout length (4.64 mm), longer than interorbital distance (3.8 mm) and eye-narial distance (2.1 mm); canthus rostralis indistinct; loreal region sloping, moderately concave; nostrils barely protuberant, very close to tip of snout (eye-nostril distance longer than distance from nostril to tip of snout); internarial region slightly concave; tympanum hidden and indistinct; tympanic fold prominent, extending from posterior corner of eye to supra-axillary region; tongue oval without posterior notch; choanae elongate oval and small, situated at anterolateral edge of palate, separated by a distance about two times of their diameter; dentigerous process of vomer in posterior edge of choanae evident, almost touching in the middle line but vomerine teeth apparently absent; vocal slits large, just posterior to rictus.
Forelimbs. Arms short, forearms not hypertrophied; hand relatively large (HAL 14.3 mm); forearm and hand length (LAHL 23.3 mm) almost as long as half of SVL (47.5 mm); fingers disks obviously dilated and tips truncated; relative length of fingers I <II <IV <III; fingers without dermal fringe on inside of fingers, two side protuberant osseous tubercles on the upper surface of the tips of fingers and webbing absent; subarticular tubercles prominent: 1, 1, 2, 2; additional supernumerary tubercle absent; inner metacarpal tubercle oval, elongate, outer metacarpal tubercle large, flat, nearly divided; nuptial pads absent ( Figure 3 View FIGURE 3 A).
Hindlimbs. Hind limbs relatively short and brawny; heels not overlapping for long distance when legs are folded at right angle to body, and when appressed to body, tibiotarsal articulation reaching the shoulder; tibia 2.7 times longer (TL 19.7 mm) than wide (TW 7.4 mm), and shorter than foot length (FOL 21.3 mm), about 42% of SVL. Relative length of toes: I <II <V <III <IV. Tips of all toes rounded without disks, and not expanded; webbing formula I 1 + – 2- II 1 + – 2 III 1 + – 2+ IV 2 + – 1 V, toes with nearly complete webbing; subarticular tubercles prominent, simple and all present: 1, 1, 2, 3, 2; tarsus smooth, tarsal fold absent, outer metatarsal tubercle round, small, and pointed; inner metatarsal tubercle big, prominent, with sharp outer edge, about two-thirds length of toe I ( Figure 3 View FIGURE 3 B).
Skin. Skin of dorsal surfaces of body, head, and limbs smooth; ventral surfaces of throat, trunk, and limbs smooth; loose skin overlying median subgular vocal sac forms slight sternal fold; ventral epidermal adhesive gland occupies sternal region and venter.
Color of holotype in life. Dorsal parts of head and dorsum, flank, forelimb, thigh, tibia and foot light olivegreen with dark moss-green marbling; loreal region, tympanic region and tympanum slightly dark olive-green; dorsal parts of two sides shoulder light olive-green; lower part of flanks with some lemon spots; lower jaw, throat, margin of throat and chest beige with some small lemon spots; belly, ventral part of forelimbs and hind limbs creamy white.
Color of holotype in preservative. Dorsum brown with dark marbling. Ventral surface of chest, throat, belly, interior portions of arms and thighs cream color.
Measurements. Holotype: SVL 47.5, HL 11.7, HW 15.2, SL 4.6, IND 2.8, IOD 4.1, UEW 2.7, ED 4.6, LAHL 23.3, HAL 14.3, HLL 62.7, TL 19.7, TW 7.1, FOL 21.3.
Etymology. The species epithet is a Latin adjective referring to the locality in which the new species was collected. The suggested English name is the Nonggang narrow-mouthed frog.
Tadpole. Six tadpoles (Voucher No.: NHMG_T20110801, NHMG_T20120901–05) from stages 36 to 42 sensu Gosner (1960) were collected from the type locality of K. nonggangensis sp. nov. Morphological measurements and characteristics were examined following Altig & McDiarmid (1999). The 16S rRNA sequence (~540 bp) of a specimen (Voucher No.: NHMG_T20120901) is 99% identical to a reference sequence of paratype (Voucher No.: NHMG1108036). Measurements of one tadpole at developmental stage 38: total length 37.5 mm, head-body length 0.54 time of tail length; eyes lateral, small, and visible from ventral view. In dorsal view, body nearly rectangle, snout broadly rounded. Interpupilar distance 0.84 maximum body width, internarial distance 0.17 of interpupilar distance. Width of oral apparatus 0.27 head-body length; lips not expanded; lacking labial teeth, horny beak and papillae. Nostrils closed at stage 38. Spiracle median, opening slightly anterior the end of body, free portion with an arched membrane, the inner wall attached to body wall; anal tube median and elongate, and the inner wall fused to ventral fin. The base of tail musculature strong, its height 0.35 of tail height; maximal tail height located at the proximal 1/2 of the tail, caudal muscles tapering gradually. Dorsal fin originates at the tail bodyjunction and the ventral fin originates at the ventral terminus of the body. The dorsal fin is nearly equal to the ventral fin in height. Dorsal fin and ventral fin have some yolk-yellow spots. Especially, dorsal fin near the tail body-junction has a yolk-yellow line. Tail tip broadly rounded. Head and body brownish with some faint yellow pigments ( Figure 2 View FIGURE 2 G, H).
Variation. Individuals of the type series are generally similar in morphology. The female (NHMG1108035) is two-third webbed (webbing formula, I 2 - – 2+ II 2 - – 3 III 2 – 3 IV 3 + – 2- V). Two specimens (NHMG1108035–36) have some small spinous tubercles in the anal region. Five specimens (NHMG1106033, 1108035–36, 1108038, 1108040) have two subarticular tubercles on the third toe, and the remaining specimens have three subarticular tubercles on the third toe. Four individuals (NHMG1106040–42, 1108037) have a slightly rough dorsum scattered with a few small rounded tubercles. The tympanic fold is indistinct in six individuals (NHMG1106036, 1108037– 39, 1108040–41). All males have two side protuberant osseous tubercles on the upper surface of the tips of fingers. Between two side protuberant osseous tubercles, four individuals have small tubercles ranging from 1 to 4 (NHMG1106031: L II (left finger II), 3; NHMG1106033: L II, 4; NHMG1108036: R I (right finger I), 2; NHMG1108039: L I, 1; R III, 1), which size is about 1/5 of two side protuberant osseous tubercles. Female also have two side protuberant osseous tubercles but which size is only 1/2 that of the male. In preservative, seven specimens (NHMG1106031–34, 1106035, 1106039, 1108040) have a gray dorsum, and the remaining specimens are dark brown.
Male secondary sexual characters. Nuptial pad absent; external subgular vocal sac present; throat with some lemon spots; chest and belly with epidermal adhesive gland.
Ecology. K. nonggangensis sp. nov. were observed in primary or secondary karst evergreen forest, and cultivated fields near the forest. Presently, K. nonggangensis sp. nov. is only known from the Nonggang National Nature Reserve. Sites where we observed the species ranged from 150–200 m elevation. On June 29 and August 11, 2011, the species was observed to gather in the temporary plash after rainstorm.
Sequence divergence. Uncorrected sequence divergences between K. nonggangensis sp. nov. sequences and all homologous sequences available on the genus Kaloula are listed in Table 2. The uncorrected p -distance genetic distance between K. nonggangensis sp. nov. and K. verrucosa tissues examined was 1.4–1.6%, while that between K. nonggangensis sp. nov. and other species was>1.8% (Table 2).
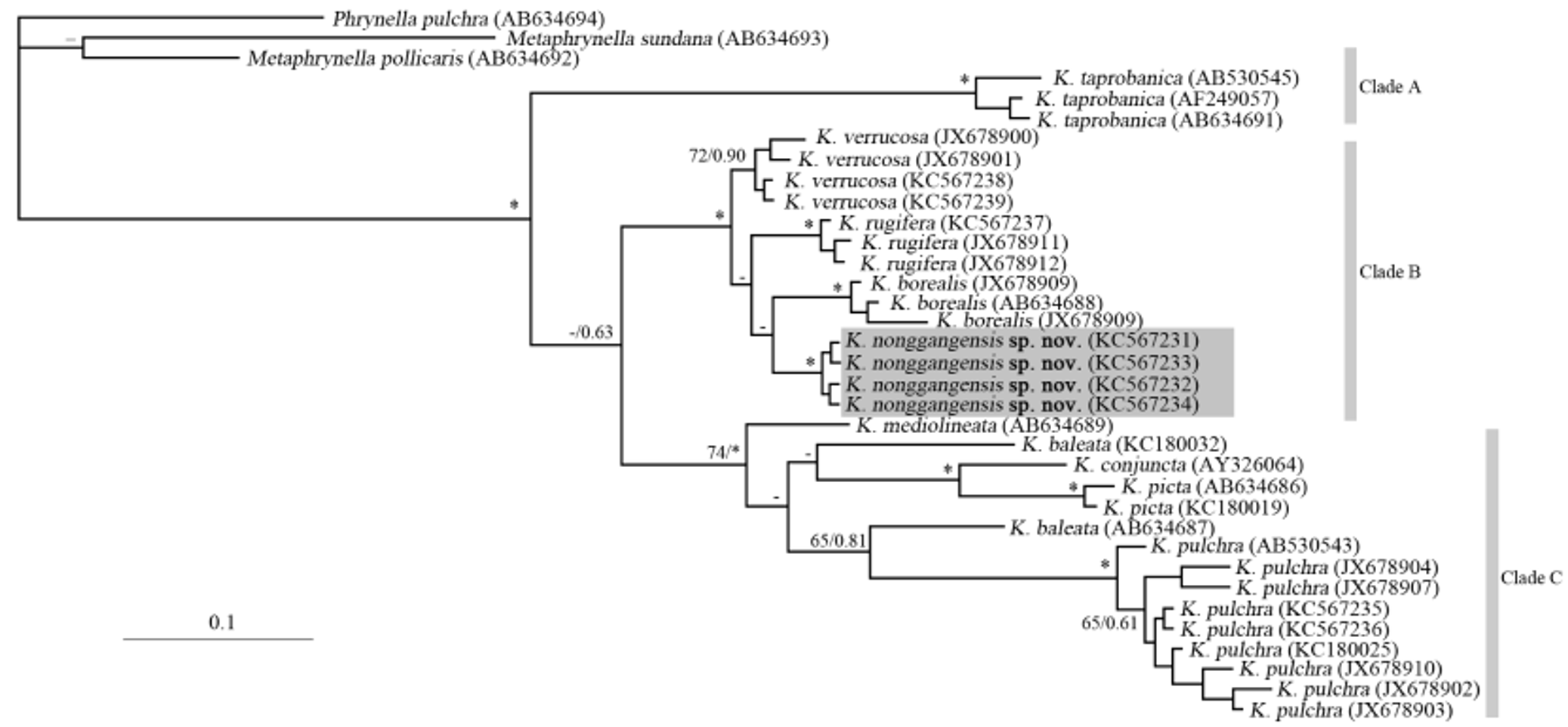
Preliminary hypothesis of phylogenetic relationship. Among 15 Kaloula species (Frost 2013), only 9 species' sequences (including K. baleata , K. borealis , K. conjuncta , K. mediolineata , K. picta , K. pulchra , K. rugifera , K. taprobanica , K. verrucosa ) are available from GenBank. Based upon our preliminary molecular data, the genus Kaloula formed a monophyletic group with well-supported values (BP=98; BBP=1.00), and was divided three clades ( Figure 4 View FIGURE 4 ). Clade A consists of Kaloula taprobanica Parker, 1934 ; Clade B consists of K. borealis , K. rugifera , K. verrucoca , and K. nonggangensis sp. nov. and Clade C consists of K. baleata van & Muller, 1836, K. conjuncta Peters, 1863 , K. mediolineata Smith, 1917 , K. picta Duméril & Bibron, 1841 , and K. pulchra Gray, 1831 .
Comparisons: K. nonggangensis sp. nov. differs from all other species of Kaloula by having smooth or slightly rough dorsum without rough tubercles, finger tips widely expanded and truncated, two side protuberant osseous tubercles on the upper surface of the tips of fingers in male, throat and chest with some small lemon spots in male, and larvae with some yolk-yellow spots in body and tail. K. nonggangensis sp. nov. is most similar in appearance to K. borealis , K. rugifera and K. verrucosa ( Figure 2 View FIGURE 2 ). However, K. nonggangensis sp. nov. differs from K. borealis by having widely expanded terminal digital disks (vs. lacking expanded terminal digital disks in the latter); by two side protuberant osseous tubercles on the upper surface of the tips of fingers in male (without osseous tubercles in the latter, Figure 3 View FIGURE 3 D); by having nearly full webbing (vs. 1/2 webbing in the latter); by throat and chest beige with some small lemon spots (vs. distinct black spots in the throat area in the latter) (Fei et al. 2009). Furthermore, in life, the tadpole of K. borealis has dark brown dorsal body and tail, without pigment and venter white or hoar (vs. the tadpole of K. nonggangensis sp. nov. has brownish head and body with some faint yellow pigments and dorsal fin near the tail body-junction has a yolk-yellow line) (Fei et al. 2009; Zhou et al. 2011). K. nonggangensis sp. nov. differs from K. rugifera by having a relatively bigger body size (SVL 41.4–52.7 mm in male vs. 35.5–43.0 mm in male for K. rugifera ) (Table 3). It can be further distinguished from K. rugifera by finger tips widely expanded and truncated (slightly expanded in K. rugifera , Figure 3 View FIGURE 3 F); by two side protuberant osseous tubercles on the upper surface of the tips of fingers in male (having two clusters of osseous tubercles instead of two side protuberant osseous tubercles in the latter) ( Figure 3 View FIGURE 3 F); by throat and chest with some small lemon spots in male (vs. cream without pigments); by larvae with some yolk-yellow spots in body and tail (vs. dorsum and tail dark brown, no pigment) (Fei et al. 2009; Zhou et al. 2011). K. nonggangensis sp. nov. also differs from K. verrucosa by smooth or slightly rough dorsum without rough tubercles (rough dorsal skin and tubercles present in K. verrucosa ) ( Figure 2 View FIGURE 2 F); by finger tips widely expanded and truncated (non-expansion of terminal digital disks in the latter, Figure 3 View FIGURE 3 E); by two side protuberant osseous tubercles on the upper surface of the tips of fingers in male (4–6 free osseous tubercles in the latter) ( Figure 3 View FIGURE 3 E); by larvae with some yolk-yellow spots in body and tail (vs. dorsum and tail dark brown with black pigment in the latter) (Fei et al. 2009). The morphological differences in the shape of terminal disks ( Figure 3 View FIGURE 3 ) have been assessed in large numbers of specimens in the studies of 83 for K. borealis , 100 for K. verrucosa and 67 for K. rugifera , partly in specimens from different localities (Kunming and Dali in K. verrucosa ) (Appendix I, Fei et al. 2009).
K. nonggangensis sp. nov. differs from K. aureata Nutphand, 1989 , by having an olive dorsum without lateral bands (vs. bright yellow in median dorsum, and yellowish orange lateral bands along the back) (Pauwels & Chérot, 2006). K. nonggangensis sp. nov. differs from K. baleata by having a ventral adhesive gland in males (vs. venter without epidermal adhesive gland); by having olive dorsum without rough tubercles (vs. brownish dorsum with rough tubercles, and a bright yellow spots near axilla in the latter) (Pauwels et al. 1999). K. nonggangensis sp. nov. differs from K. conjuncta , K. kalingensis , K. kokacii and K. walteri by a larger body size (SVL 41.4–52.7 mm in males vs. 26.6–30.1 mm in K. conjuncta , 24.2–32.9 mm in K. kalingensis , 37.0– 39.1 mm in K. kokacii , 24.5–31.5 mm in K. walteri (Diesmos et al. 2002) . It also differs from K. conjuncta by the absence of rough dorsal tubercles (vs. tubercles distributed over the entire dorsum); by having olive dorsum (vs. brownish dorsum) (Diesmos et al. 2002). It also differs from K. kalingensis and, K. kokacii by its ventral adhesive gland in males (vs. venter without epidermal adhesive gland) (Brown et al. 2000; Diesmos et al. 2002). It also differs from K. walteri by the presence of a round, small, and pointed outer metatarsal tubercle (vs. absence, or presence of only a faint, very reduced outer metatarsal tubercle); by the presence of three subarticular tubercles on the fourth toe (vs. two); by having widely expanded terminal digital disks (vs. lacking expanded terminal digital disks in the latter) (Diesmos et al. 2002). K. nonggangensis sp. nov. clearly differs from K. assamensis , K. mediolineata , K. pulchra and K. taprobanica by absence (vs. presence) of dorsolateral bands. It also differs from K. assamensis by its inner metatarsal tubercle smaller (vs. larger) than first toe; lacking a vertebral stripe (vs. presence of a dark edged lemon vertebral stripe) (Das et al. 2004; Nath et al. 2011). It also differs from K. mediolineata by its finger tips widely expanded (vs. non expanded) and truncated (Diesmos et al. 2002). It also differs from K. pulchra by its smaller body size (SVL 41.4– 52.7 mm vs. 55.0–77.0 mm in K. pulchra (Fei et al. 2009) ; snout length about 1/3 head length (vs. half of the head length; by smooth venter (vs. granular venter) (Fei et al. 2009). It can be further distinguished from K. taprobanica by lacking (vs. having) large irregular shaped markings of yellow-red color in dorsum median area (Dutta & Manamendra-Arachchi 1996). K. nonggangensis sp. nov. differs from K. picta and K. rigida by absence (vs. presence) of supernumerary tubercles at the base of each digit of the manus. It also differs from K. picta by its inner metatarsal tubercle shorter (vs. equal to or longer) than first toe (Diesmos et al. 2002). Finally, K. macrocephala (originally treated as a synonym of K. pulchra ) differs from K. nonggangensis sp. nov. by having indistinct dorsolateral bands and mid-dorsum covered by large-sized irregular patches (vs. clearly absent dorsolateral bands in new species) (Bourret 1942; Ohler 2003; Pauwels & Chérot 2006).
In addition, genetically, except for K. borealis , K. rugifera and K. verrucosa , the uncorrected sequence divergences between K. nonggangensis sp. nov. 16S rRNA sequences and all homologous sequences available on GenBank (Table 2) were greater than 3%, a value usually representing differentiation at the species level in frogs (Vences et al. 2005; Fouquet et al. 2007).
Discussion
Fei et al. (2009) divided Chinese Kaloula species into two groups: the K. pulchra group (consisting of K. pulchra ) and the K. verrucosa group (consisting of K. borealis , K. rugifera and K. verrucosa ), being consistent with our phylogenetic trees. K. nonggangensis sp. nov. embedded within the K. verrucosa group. K. nonggangensis sp. nov., K. borealis , K. rugifera and K. verrucosa formed a monophyletic group with high supported values ( Figure 4 View FIGURE 4 ). Preliminary molecular data indicated that K. nonggangensis sp. nov. and K. borealis were sister species. However, due to weak inter-nodes support values (<60%), their phylogenetic positions must be considered as unresolved. We found that the genetic distances among lineages in the K. verrucosa group are lower than the values usually representing differentiation at the species level in frogs (Vences et al. 2005; Fouquet et al. 2007) (Table 2). According to the results of these authors, in most cases species are differentiated by uncorrected p -distances in 16S rRNA of 3% or higher. If the criterion was strictly applied in this group, then K. nonggangensis sp. nov., K. borealis , K. rugifera and K. verrucosa could also be considered as conspecific populations. However, the morphological differences separating these four taxa, including those in larval morphology, are quite distinct ( Figure 2 View FIGURE 2 , 3 View FIGURE 3 and Table 3), and extend to characters that do not usually show intraspecific variability in anurans. For instance, K. borealis differs by lacking osseous tubercles on the upper surface of the tips of fingers in male, and the finger not being dilated. The digital tips of K. verrucosa are not dilated either. Tadpoles of K. borealis , K. rugifera and K. verrucosa have relatively dark colored in tail muscles and tail fins (vs. having some yolk-yellow spots in K. nonggangensis sp. nov. tadpole) (Fei et al. 2009). Besides, although the diploid number among K. borealis , K. rugifera and K. verrucosa was of 2n=28 chromosomes, their karyotypes are different. The K. borealis karyotypes consisted of eleven metacentric (1, 2, 4–11, 13) and three submetacentric (3, 12, 14) pairs. The K. rugifera karyotypes consisted of eleven metacentric (1, 2, 4–12), one submetacentric (pair 3), one subtelocentric (pair 13) and one telocentric (pair 14) pairs. The K. verrucosa karyotypes consisted of eleven metacentric (1, 2, 4–7, 9–11, 13, 14) and three submetacentric (3, 8, 12) pairs (Wu & Yang 1981; Zeng & Wu 1990; Zhang et al. 2006). Despite the low genetic distances among group, considering their distinct morphology and karyotype, we suggest that they are separated species. In fact, in Asian microhylid genera, a low genetic differentiation among some species has also been reported by Matsui et al. (2011). For instance, the uncorrected p -distance of Micryletta inornata (GenBank no.: AB611968 View Materials ) and Micryletta steinegeri (GenBank no.: AB634696 View Materials ) in 16S rRNA sequence (~540 bp) is 2.8%. Some 16S rRNA sequences of Asian microhylid (data mainly coming from the paper of Matsui et al. (2011) (Appendix II) together our data indicate that the branches in Kaloula are relatively short (figure no showed). This may either indicate that Kaloula are very young and evolved in a recent rapid radiation, or that mitochondrial substitution rates have become decelerated in this genus. To conclude, a re-evaluation of the species boundaries and the phylogenetic relationships within the genus, using larger samples and bioacoustics is warranted.

No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.