
Ronzotherium romani Kretzoi, 1940
|
publication ID |
https://doi.org/ 10.5852/ejt.2021.753.1389 |
|
publication LSID |
lsid:zoobank.org:pub:8009DD3B-53B0-45C9-921E-58D04C9C0B48 |
|
DOI |
https://doi.org/10.5281/zenodo.4959528 |
|
persistent identifier |
https://treatment.plazi.org/id/C53FFD4D-D727-FF9E-FDAD-DC811956A6DB |
|
treatment provided by |
Felipe |
|
scientific name |
Ronzotherium romani Kretzoi, 1940 |
| status |
|
Ronzotherium romani Kretzoi, 1940
Figs 15–21 View Fig View Fig View Fig View Fig View Fig View Fig View Fig
Ronzotherium romani Kretzoi, 1940: 91 .
Hypsolophiodon csobánkanus Kretzoi, 1940: 94–95, fig. 6.
? Praeaceratherium kerschneri Spillmann, 1969: 241–253 , figs 4 (bottom), 6, 13 (left), 15, pls XX– XXII.
Diaceratherium massiliae Ménouret & Guérin, 2009: 314–323 , figs 10, 12a, 13a, b?, c–e, 14a?, 15a, 16a.
Acerotherium filholi – Roman 1910: 1559 (from La Ferté Aleps = La Ferté-Alais, La Comberatière and
Marseille); 1912a: 55–56, fig. 17, pl. V–3. Ronzotherium filholi – Bonis 1969: 1–8, pls 1–2. — Ginsburg 1969: 1267. Ronzotherium velaunum – Heissig 1969: 20–36, figs 5, 6d, 7, 8d–g, 9c, 10c–d (from St-André, St-Henri,
Marseille and Les Milles).? Ronzotherium filholi elongatum – Heissig 1969: 47, 53, 82 (from Cournon). Ronzotherium filholi romani – Heissig 1969: 55–90, figs 20–24, 26c–d, pls 1–3, 4(13). Ronzotherium romani – Brunet 1979: 155, figs 7a, 9b, d, 10b, pls XVIb–n, XIXg–l, o–p (from Ferté-
Alais, Etampes, Gaimersheim). — Brunet et al. 1981: 349. — Ginsburg & Hugueney 1987. —
Ménouret & Guérin 2009: 306–314, figs 2, 7–9. — Mennecart et al. 2012: 166–169, fig. 3(3, 4?,
5–7, 8?, 9?) (partim). — Ménouret et al. 2015: 245–248, figs 4–5a–d. Diaceratherium lamilloquense – Mennecart et al. 2012: 169, figs 3(10–11, 16), 4 (NMB-UM-2565)
(partim). Diaceratherium massiliae – Antoine & Becker 2013: 140. — Jame et al. 2019: 21. “ Diaceratherium ” massiliae – Becker et al. 2018: 401. “ Diaceratherium massiliae ” – Blanchon et al. 2018: 219. Non Ronzotherium filholi – Lavocat 1951: 116, pl. 19 fig. 3, pl. 26 fig. 1 (from Vendèze).
Non Ronzotherium romani – Brunet 1979: 154, fig. 15, pls XVII–XVIII (from Vendèze).
Non Ronzotherium romani – Mennecart et al. 2012: fig. 4 (NMO-K3/13, NMOI10/103) (partim).
Historical diagnosis
Kretzoi (1940) did not provide any proper diagnosis when he named the species in a footnote. The only mentioned characters are that “the molars have higher crown and less forward inclined, more verticallystanding lophs” than Ronzotherium filholi (translated by the authors).
Emended diagnosis
The I1 is oval in cross-section and the crochet and crista are sometimes present on the upper molars. The lingual cingulum of the upper cheek teeth is usually absent. The protoloph of P2 is mostly interrupted and disconnected from the ectoloph. The posterior valley of d2 is usually open. The lingual cingulum of the lower premolars is usually absent. The radius and ulna are in contact or fused and there is a single distal contact facet. The gutter for the musculus extensor carpi is weak on the radius and the proximal ulna facets are not always separated. The trapezium facet is small on the scaphoid. The transverse diameter/height ratio of the astragalus is above 1.2 and the posterior stop on the cuboid facet is absent. The Cc1 facet of the astragalus is nearly flat. The proximal border of the anterior side of the MtIII is concave.
It differs from R. velaunum by the deep median constriction of the distal humeral articulation and from R. filholi by the absence of i1, the single-rooted d/p1, the reduced paraconid on p2 and the high posterior expansion of the scaphoid facet on the radius.
Type material
Lectotype (designated by Heissig 1969)
FRANCE • right lower i2; Essone, La Ferté-Alais; MNHN.F.OBP63.
Paralectotypes (designated by Brunet 1979)
FRANCE • 4 lower molars; same collection data as for lectotype; MNHN.F.OBP72, MNHN.F.OBP76, MNHN.F.OBP78, MNHN.F.OBP79 .
Additional material
FRANCE – La Ferté-Alais, Essone • 1 P2; same collection data as for holotype; MNHN.F.OBP55 • 1 P 4; same collection data as for holotype; MNHN.F.OBP56 • 1 lingual fragment of P4; same collection data as for holotype; MNH.F.OBP57 • 1 M1; same collection data as for holotype; MNHN.F.OBP59 • 3 M 2; same collection data as for holotype; MNHN.F.OBP58 , MNHN.F.OBP60 , MNHN.F.OBP61 • 1 M 3; same collection data as for holotype; MNHN.F.OBP62 • 1 d1; same collection data as for holotype; MNHN.F.OBP86 • 1 p 2; same collection data as for holotype; MNHN.F.OBP65 • 2 p 3; same collection data as for holotype; MNHN.F.OBP66 , MNHN.F.OBP67 • 2 p 4; same collection data as for holotype; MNHN.F.OBP68 , MNHN.F.OBP69 • 6 additional lower molars excluding the paralectotypes; same collection data as for holotype; MNHN.F.OBP70 , MNHN.F.OBP71 , MNHN.F.OBP73 , MNHN.F.OBP74 , MNHN.F.OBP75 , MNHN.F.OBP77 . – St-Henri / St-André / Les-Milles • 1 complete maxilla P1- M2 (left) and P1-M3 (right) with subcomplete mandible with p2–m3 (left) and p3-m3 (right); FSL- 8547 • 1 fragment of right maxilla with P1–3; FSL-520275 (not found in collection) • 1 left I1; FSL-8835 • 2 right I2 and 1 left I2, with the same inventory number; NMB-Mar-354a (not found in collection) • 1 left i2; FSL-9445 (not found in collection) • 2 right I2; FSL-9524 (not found in collection) and FSL-9448 • 1 right P1; FSL-9519 • 1 right D3; FSL-8557 • 1 left D4; FSL-9530 • 1 right P2; FSL-8834 • 1 left P2; FSL-8833 • 1 left P3; FSL-8832 • 1 right P4; NMB-Mar-844 • 2 left M3; FSL-8828 , NMB-Mar-862 • 2 right M3; FSL-520290 (not found in collection), NMB- Mar-862 • 1 subcomplete mandible with p3–m3 (right) and p2–4 and m2–3 (left); FSL-8545 • 1 right hemimandible with p3–m3; NMB-Mar-843 , NMB-Mar-861 • 3 i 2; NMB-Mar-862 (right and left), FSL-9524 • 1 d1; FSL-9521 • 2 rows with d2–3; FSL-9520 , FSL-9518 (right), FSL-9517 , unnumbered specimen (possibly FSL-9519?) (left) • 1 left p3; FSL-8831 • 1 right P4; FSL-520277 (not found in collection) • 1 right m1/2; FSL-520277 (not found in collection) • 3 left m2; FSL-8827 (not found in collection), FSL-8829 (not found in collection), FSL-8830 • 1 right m2; FSL-520278 (not found in collection) • 1 right m3; NMB-Mar-1 • 1 left scapula; AIX.1979-2 • 1 distal humerus; FSL-9523 • 1 radius in two fragments; FSL-520279 , FSL-520280 • 1 scaphoid; FSL-520285 • 2 trapezoids; FSL-9501 , FSL-520283 • 2 unciforms; FSL-520289 , FSL-520282 • 1 unciform; NMB-Mar-865 • 1 left McIII; UPM 13667 • 2 fragments of McIII; FSL-9505 , FSL-520281 • 1 McIV; NMB-Mar-863 • 1 proximal fragment of McIV; NMB-Mar-864 • 1 McIV; FSL-520287 • 1 distal femur; NMB-Mar-828 • 1 navicular; NMB-Mar-847e • 1 cuboid; FSL-9528 • 1 cuboid; NMB-Mar-847d • 1 MtII, originally identified as a McII by Ménouret & Guérin (2009); NMB-Mar-847a • 1 MtIV; FSL-520286 .
GERMANY – Gaimersheim • 1 axis and several fragments of vertebrae, 1 complete radius, 1 complete scaphoid and a fragmentary one, 1 partial magnum, 1 broken McIII in articulation with a well preserved McIV as well as an incomplete tibia, 1 almost complete astragalus, and all dental specimens already attributed to R. romani by Heissig (1969); BSPG (unnumbered) • 1 left MtIII; BSPG- 1952-II .
SWITZERLAND – Jura Canton, Poillat • 1 fragment of squamosal; MJSN-POI-007-59 • 1 maxilla with P1–M3; MJSN-POI-007-3219 • 1 isolated I2; MJSN-POI-007-168 • 1 isolated P4; MJSN-POI-007-346 • 1 juvenile hemimandible with i2 –p1–m1 and erupting m2; MJSN-POI-007-174 • 1 isolated i2; MJSN-POI-007-937 • 1 isolated p4; MJSN-POI-007-211 • 2 scapulae; MJSN-POI-007-306 , MJSN-POI-007-222 • fragments of lumbar vertebra IV; MJSN-PRC-005-1 • 1 sacrum; MJSN- BEU-001-280 • 1 left femur; MJSN-POI-007-80 . – Zürich Canton,Rickenbach • 1D3/4; NMB-UM-971 • 1 P 2; NMB-Ri-24 • 1 P 4; NMO-H9-13 • 1 M 1; NMB-UM-972 • 2 M 1; NMO-K11/250 , NMO-I12/13 • 1 M 2; NMO-I12-24 • 1 M 2; NMB-Ri27 • 1 maxilla fragment with P4–M1; NMB-UM-1840 • 1 maxilla fragment with M2–3 (unnumbered in SMNS collection); SMNS • 1 mandible with left and right p2–m3; NMB-UM-3832 • 1 p 3; NMB-H.R.2 • 1 p 4; NMO-L6/25 • 1 m 1; NMB-UM-806 • 1 broken humerus; NMB-UM-973 • 1 scaphoid; NMO-I5-62 • 1 lunate; NMB-Ri-21 • 1 lunate; NMO-I7-115 • 1 pyramidal; NMO-I11-82 • 1 magnum; NMO-H10-110 • 1 McIV; NMO-I8-117 • 1 MtII; NMB-UM-2565 .
The specimens from St-Henri/St-André/Les-Milles have previously been attributed to“ Diaceratherium ” massiliae .
Type horizon and locality
La Ferté-Alais (Essonne, France), MP24 (latest early Oligocene).
Stratigraphical distribution
?MP23 (early Oligocene) to MP30 (latest Oligocene).
Geographical distribution
France: Aubenas-les-Alpes, La Bénissons-Dieu, Brons, La Comberatière, Cournon, Étampes, Gannat?, Itteville, Pech Desse, Sainte-Quitterie, St-Henri/St-André/Les-Milles (= ‘Marseille’), Vodable. Germany: Gaimersheim. Hungary: Csobánka. Switzerland: Poillat, Rickenbach, Rüfi bei Schänis.
Description
Material from the type locality Part of this material was already described ( Heissig 1969; Brunet 1979) but we provide here some short updated descriptions. Only isolated teeth are preserved from La Ferté-Alais.
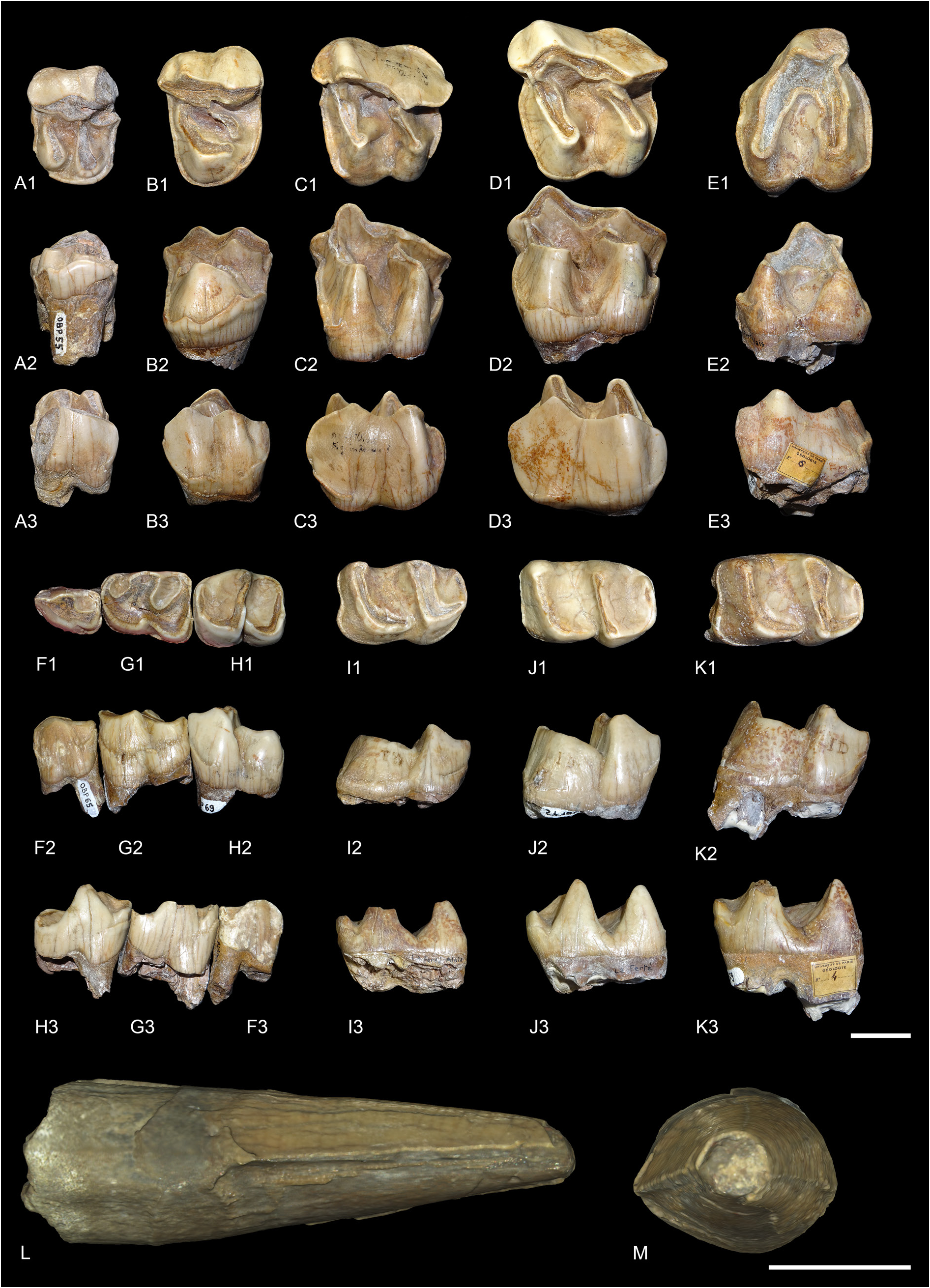
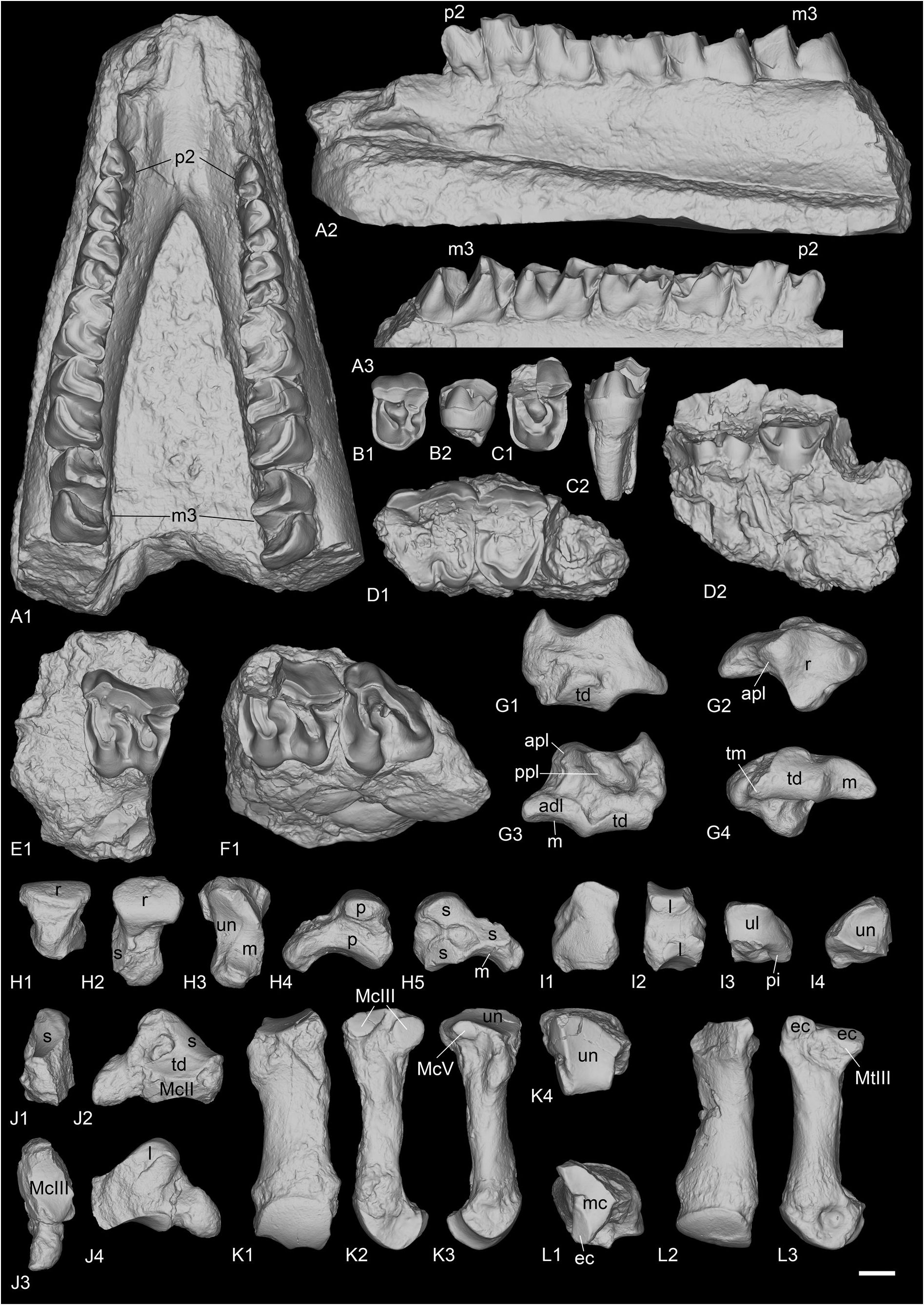
ANTERIOR DENTITION. The lectotype right lower i2 (MNHN.F.OBP63) is large and tusk-like ( Fig. 15L– M View Fig ). The root and the tip of the crown are broken, and the enamel is thin. The wear facet for the upper I1 is probably absent, either because of the absence of contact between these two teeth or because the tooth was not completely erupted if it belonged to a young individual. The transverse outline of the crown is drop-shaped, whereas the root is oval-shaped. There is a sharp mesial crest on the mesial border of the crown as well as a weaker crest on the lateral border. There is also a distomesial cingulum.
UPPER CHEEK TEETH. Seven isolated upper cheek teeth are preserved in La Ferté-Alais ( Fig. 15A–E View Fig ), but no upper incisors.
UPPER PREMOLARS. Only P2 (MNHN.F.OBP55) and P4 (MNHN.F.OBP56) are preserved. A lingual fragment of P4 (MNH.F.OBP57) is also preserved but is not informative. The lingual cingulum is strong and continuous on upper premolars and is deeply rippled in lingual view. The labial cingulum is fainted between the paracone and metacone of P4 and completely absent on P2. Crochet and antecrochet are completely absent and there is no protocone constriction. On P2, the protocone and hypocone are separated, but they are united by a bridge at the base of the tooth. The protocone is as strong as the hypocone and the protoloph is separated from the ectoloph. The metaloph is transverse. The paracone and metacone folds are wide and strong, whereas the parastyle is rather weak. On P4, the protocone and hypocone are fused, there is no lingual groove separating them and the protoloph is L-shaped. It is only weakly connected to the ectoloph. The metaloph is weak, S-shaped, directed postero-lingually and does not join the protocone nor the metacone. It joins however the wide and shallow crista at the base of the paracone. The paracone and metacone folds are very strong and separated by a deep groove of the ectoloph. The parastyle is large and the metastyle short. The postfossette is long and narrow.
UPPER MOLARS. Five upper molars are preserved: one M1 (MNHN.F.OBP59), three M2 (MNHN.F.OBP58, MNHN.F.OBP60 and MNHN.F.OBP61) and one M3 (MNHN.F.OBP62). The upper molars have almost no lingual cingulum, except on one M2, where it is strong and continuous under the protocone. The labial cingulum is restricted to the posterior-most part of the ectoloph under the metacone. The antecrochet is strong on M1–2 but very weak on M3. There is a weak crochet on M1–2 that would disappear early with wear and the crista is always absent. There is no protocone constriction. The paracone fold is strong and the metacone fold and mesostyle are completely absent. The metaloph is long but the metastyle is quite short. There is a small hypostyle in the postfossette of M1, contiguous to the strong posterior cingulum. The posterior part of the ectoloph of M1–2 is very straight. The postfossette is deep, below the posterior cingulum. There is no lingual groove of the protocone. The ectoloph and metaloph of M3 are fused into an ectometaloph, and there is no posterior groove. It is quadrangular. The posterior cingulum is strong and continuous and the protocone is not constricted.
LOWER CHEEK TEETH. Sixteen lower cheek teeth are preserved in La Ferté-Alais, including six premolars and ten molars ( Fig. 15F–K View Fig ).
LOWER PREMOLARS. Only one left d1 is known (MNHN.F.OBP86). It is very simple and has two cuspids: a very large protoconid and a small posterior cusp, possibly the hypoconid. There is a small paralophid, weakly constricted, but no anterior valley. The posterior valley is more developed. There is only a very short anterior cingulum but no lingual or labial one. The root is broken. A left p2 (MNHN.F.OBP65) and a left p3 (MNHN.F.OBP66) could have belonged to the same individual, whereas the right p3 (MNHN.F.OBP67) and the right p4 (MNHN.F.OBP68) could have belonged to another. Another left p4 (MNHN.F.OBP69) cannot be attributed to any individual. The p2 and p3 bear labial vertical rugosities whereas p4 only has discontinuous cingulum. The lingual cingulum is weak and only present at the opening of the valleys. The ectolophid groove is angular on p4, but less developed on p2–3, and it always disappears before the neck. The metaconid is very slightly constricted. The entoconid is either completely absent or very weak. The posterior valley is wide and U-shaped on p4 but narrower on p2–3. The paralophid of p2 is not constricted and the anterior valley is absent. The paraconid is reduced. The posterior valley is narrowly open. The anterior branch of the paralophid is long on p3–4.
LOWER MOLARS. The isolated lower molars are difficult to differentiate from one another, so they will be discussed globally. The ectolophid groove is developed until the neck. The trigonid is angular, in right dihedron, while the talonid is rounded. The entoconid and metaconid are very slightly constricted. Lingual cingulum is only present in the posterior valley of one specimen, otherwise it is completely absent. However, the anterolingual cingulum is present in the opening of the anterior valley, though it is weak. The labial cingulum is usually present, anteriorly, labially and in the ectolophid groove, but it is always discontinuous and rather weak. The hypolophid and protolophid are slightly oblique. There is no lingual groove of the entoconid. The anterior branch of the paralophid is high and long. The opening of the anterior valley is higher than the posterior one. The posterior cingulum is always present, strong and continuous.
Material from other localities
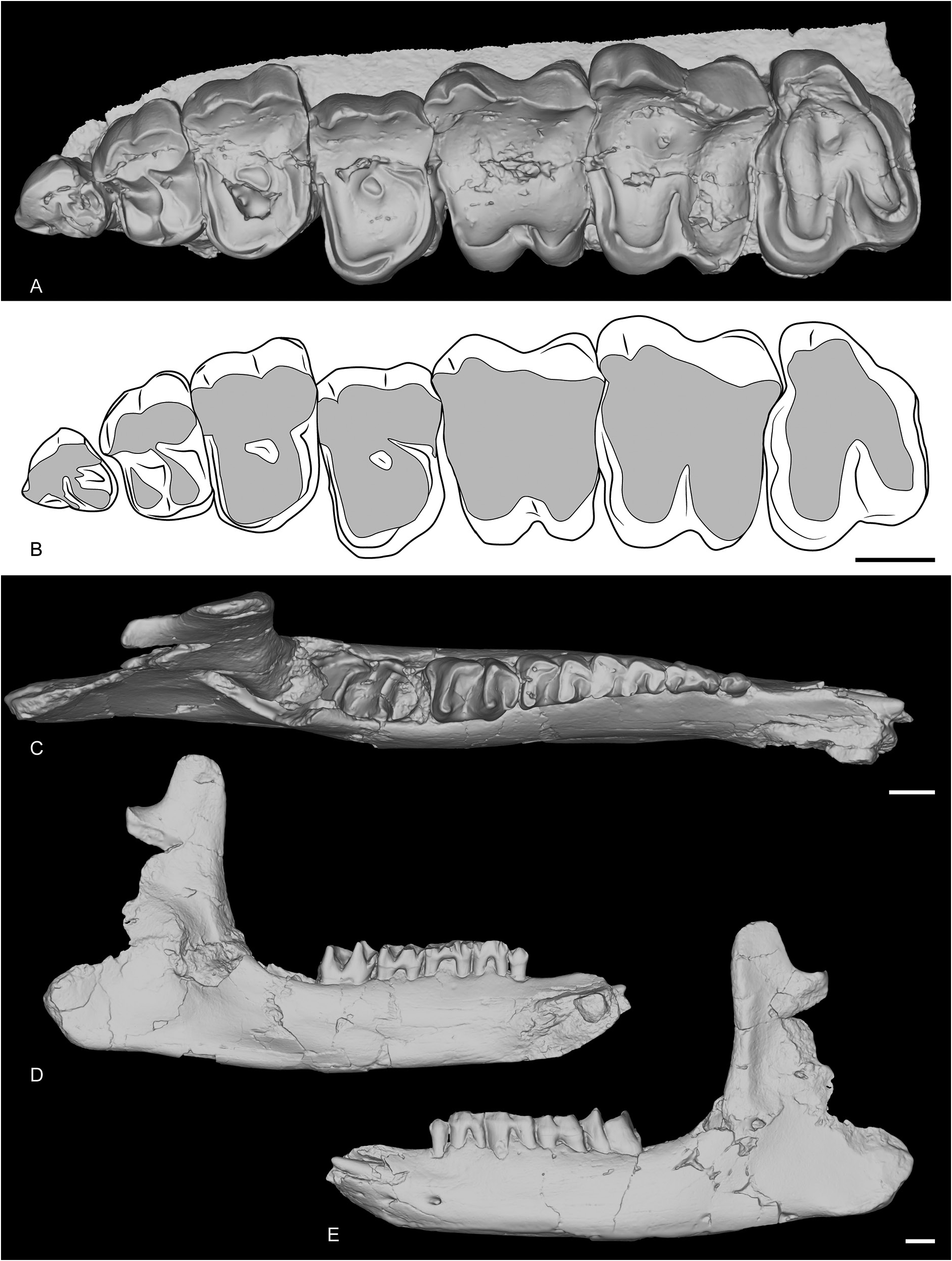
MAXILLA AND MANDIBLE FROM POILLAT. A complete upper tooth row (MJSN-POI-007-3219) and a very well-preserved juvenile mandible (MJSN-POI-007-174) are preserved ( Fig. 16 View Fig ) from the recently discovered locality of Poillat, near Delémont (Jura Canton, Switzerland), which also yielded a wellpreserved skull of Epiaceratherium delemontense Becker & Antoine, 2013 , another rhinocerotid ( Becker et al. 2013). The upper teeth are very worn, indicating a very old individual, but some characters can nonetheless be observed. The P1 is quite large, with a well-developed parastyle, and a single large lingual cusp. The ectoloph is convex. The paracone and metacone folds are strong on P2–4. There is almost no labial cingulum, but the lingual is subcomplete (it slightly faints below the protocone) and waved. The protocone and hypocone of P2 are connected by a lingual bridge. The protoloph is very short and does not connect to the ectoloph while the metaloph is oblique and connects to the paracone. The protocone and hypocone of P3–4 were either fused or connected. The molars have neither lingual nor labial cingulum, except below the metacone and at the opening of the median valley of M3. There is a posterior groove on the ectometaloph of M3.
The juvenile mandible is subcomplete. The symphysis is slightly upraised compared with the corpus mandibulae and its posterior border was just in front of d1. The foramen mentale is below p1 and there is no lingual groove for the sulcus mylohyoideus. The base of the corpus is completely straight, and the ramus is vertical. The coronoid apophysis is large and well-developed. The foramen mandibulare was below the teeth neck.
The i2 is partly unerupted. The d1 is single rooted and very simple. The posterior valley is very small. Lingual and labial cingulum are completely absent on d1–4 and m1 and there are no vertical rugosities on the ectolophid. The protoconid fold is present and the metaconid is slightly constricted on d3–4, but not the entoconid. The paralophid of d2–3 is double and the ectolophid folds are absent. The anterior groove of the ectolophid is present on d2 and its posterior valley is open lingually.
Postcranial remains
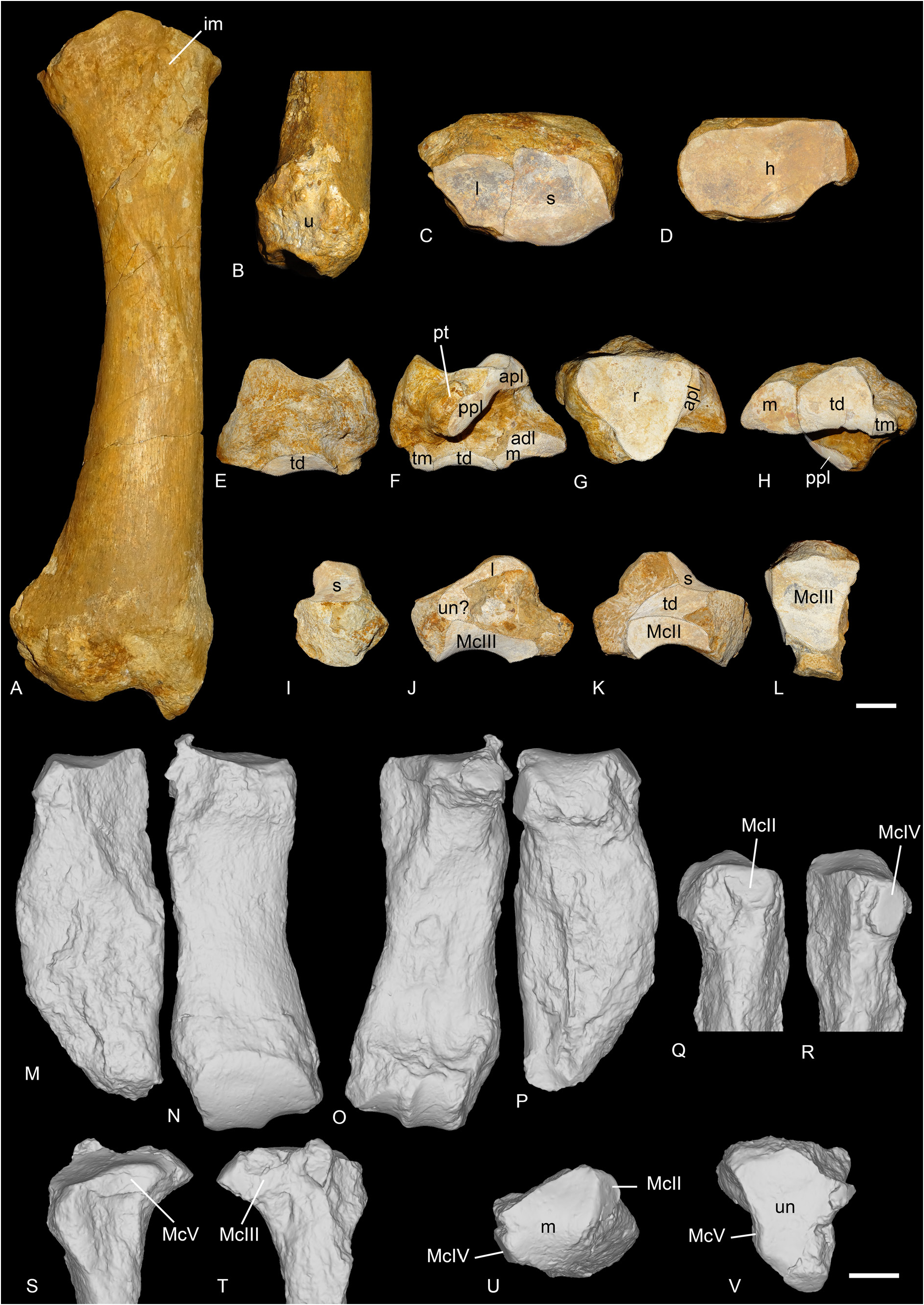
Until now, the postcranial skeleton of R. romani was almost completely unknown. No remains are preserved in the type locality of La Ferté-Alais and only a few bones were described from St-Henri/ St-André/Les-Milles ( Ménouret & Guérin 2009) and Rickenbach ( Mennecart et al. 2012): a scapula, a distal femur, a cuboid and an ectocuneiform from the former locality, and a distal humerus, two astragali and various metapods from the latter. However, after re-examination of the material from Gaimersheim, several postcranial remains can be assigned to R. romani : a complete radius, a complete scaphoid, a partial magnum, a broken McIII articulated with a well-preserved McIV as well as an almost complete astragalus and a MtIII. These specimens are of drastic importance because they can be confidently attributed to Ronzotherium , contrary to specimens from other localities such as Rickenbach
or ‘Marseille’, that were previously partly referred to as the co-occurring Diaceratherium , mostly because of their dimensions. Yet, based on fine anatomical comparisons, we now refer most of the specimens from ‘Marseille’ ( Figs 17–18 View Fig View Fig ), originally assigned to “ Diaceratherium ” massiliae (including the holotype McIV) by Ménouret & Guérin (2009), as well as new specimens from Poillat, Gaimersheim ( Figs 19–20 View Fig View Fig ) and Rickenbach ( Fig. 21 View Fig ), to R. romani . Furthermore, the two astragali previously identified as R. romani from Rickenbach ( Mennecart et al. 2012) should in fact be referred to as Diaceratherium , while the metatarsals, also identified as R. romani , should be referred to as Mesaceratherium ( Tissier et al. 2021) . These new attributions show that R. romani had a larger size than previously thought and that it was less cursorial than other species of the genus. Throughout the description, comparisons will be made with other ronzothere species as well as with Diaceratherium , and in particular D. tomerdingense , the type species of the genus, to validate the synonymy of R. romani and “ D.” massiliae .
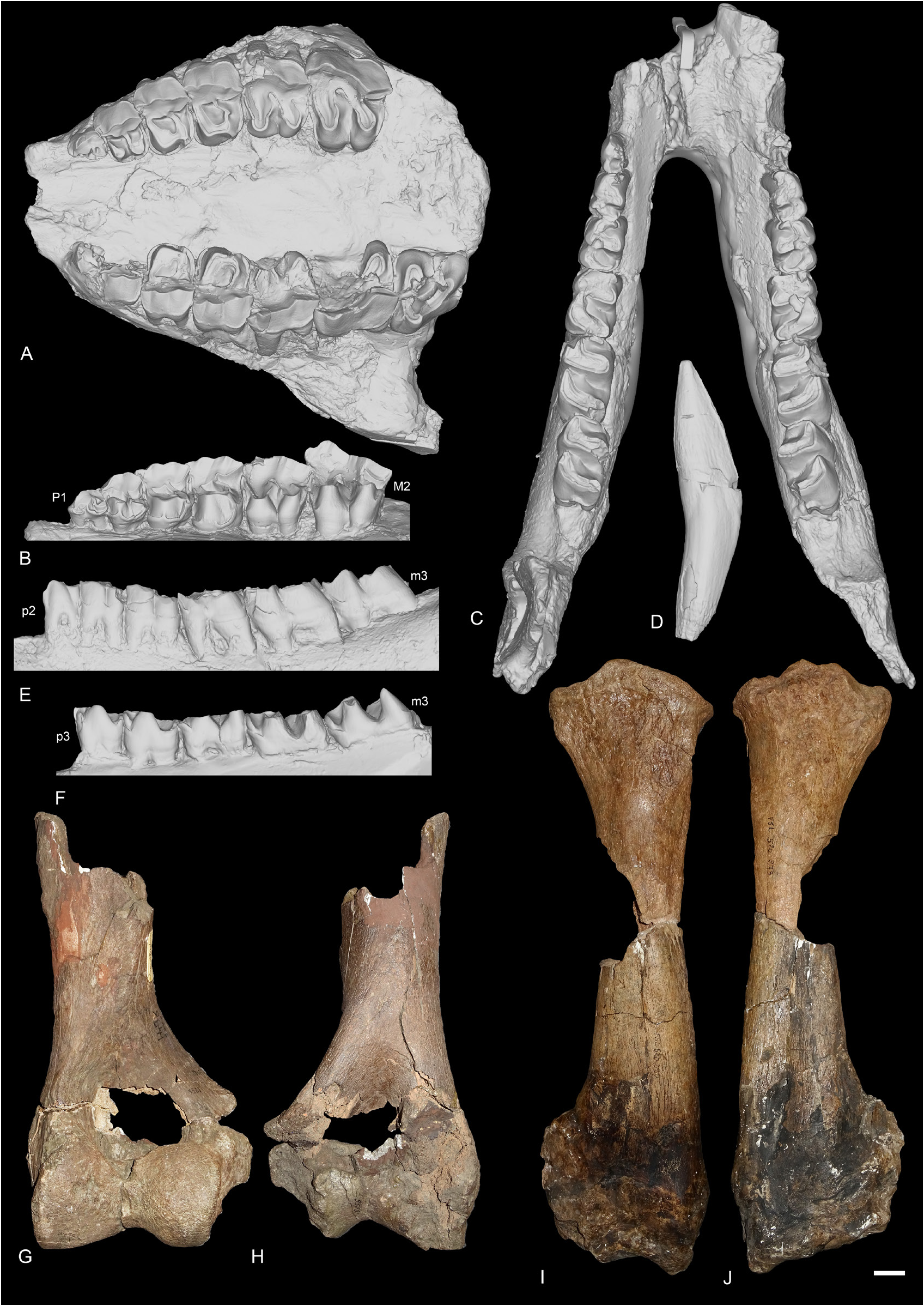
SCAPULA. Two scapulae are preserved from Poillat (MJSN-POI-007-306 and MJSN-POI-007-222), and one from ‘Marseille’ (AIX.1979-2). The two scapulae from Poillat are complete, whereas the one from ‘Marseille’ is not. It is very wide, compared to its height (= spatula-shaped, sensu Antoine 2002). In distal view, the medial border of the articulation is straight. The posterior border of the scapula and the glenoid cavity are very concave.
COMPARISON. The scapula of R. velaunum is unknown but it is preserved in Villebramar for R. filholi ( Brunet 1979) and it shares with R. romani the very concave posterior border of the scapula and glenoid cavity. The scapula of Diaceratherium aginense ( Répelin, 1917) from Laugnac ( Répelin 1917) widely differs by its reduced width compared to the height (being elongated, sensuAntoine 2002) and its slightly less concave distal border.
HUMERUS. It is known from ‘Marseille’ and Rickenbach ( Fig. 17G–H View Fig ). The distal fragment of humerus FSL-9523 from ‘Marseille’ is large-sized and incomplete. The fossa olecrani is low and wide in posterior view and the distal articulation is hourglass-shaped (or ‘diabolo-shaped’) in anterior view: there is a deep proximal incision between the two lips of the trochlea. However, there is no scar on the trochlea. The epicondylar crest is wide and laterally expanded. In anterior view, the articulation is oblique compared to the shaft of the humerus. The humerus NMB-UM-973 from Rickenbach is the most complete one but is very poorly preserved. The trochiter and the deltoid tuberosity are not preserved. The distal articulation is similar to that of other humeri from ‘Marseille’ in every aspect.
COMPARISON. The humerus of R. velaunum (PUY.2004.6.262.RON) differs by a smaller size and a higher fossa olecrani, as well as a distal articulation not medially constricted in anterior view. It further differs from the humerus from Rickenbach by a less developed lateral epicondyle. The largest humeri of R. filholi from Villebramar are more similar in size, in particular compared to the one from Rickenbach (NMB.UM-973). However, Ménouret & Guérin (2009) had referred the humerus from ‘Marseille’ FSL-9523 to Diaceratherium massiliae based on the size difference with R. filholi , as well as morphological differences, such as the wide coronoid and olecranon fossae. Yet, the humerus Vil.1970-225 of R. filholi from Villebramar ( Brunet 1979: pl. XXI, fig. a) is very similar to FSL-9523: the two fossae are wide and the epicondylar crest and lateral epicondyle are equally developed. It only differs by less constricted condyles in anterior view. Furthermore, we believe that another very large humerus from ‘Marseille’ (FSL-8546; but seeMénouret & Guérin 2009: fig. 10a–b, which is indeed FSL- 8546 and not FSL-9523, contrary to the legend of the figure) attributed to D. massiliae by Ménouret & Guérin (2009) could in fact come from another locality, based on its very different preservation. Finally, the humeri of D. tomerdingense (type species of the genus; SMNS-16154), D. lamilloquense Michel, 1987 (NMB-L.M.429) and even D. aginense ( Répelin 1917: pl.VIII, fig. 1) all have a rather high fossa olecrani in posterior view, and unconstricted condyles in anterior view.
RADIUS. It is preserved from ‘Marseille’ and Gaimersheim. The radius from ‘Marseille’ FSL-520279+ 520280 ( Fig. 17I–J View Fig ) is subcomplete, but the distal articulation is poorly preserved. In proximal view, the anterior border of the articulation is straight.The proximal facets for the ulna are separated in posterior view. The lateral one is large and concave. The medial border of the diaphysis is straight in anterior view. In anterior view, the insertion for the m. biceps brachii is marked and deep. The gutter for the m. extensor carpi is very shallow on the distal articulation. The radius from Gaimersheim (BSPG collection, Fig. 19A–D View Fig ) is complete and very well preserved. It shows that the radius was connected to the ulna over three quarters of the diaphyseal length. In proximal view, the medial articulation facet for the humerus is much larger than the lateral one and they are both concave. The medial articulation for the ulna is a thin lateromedially elongated band, whereas the lateral articulation is large and triangular. Distally, there are two poorly distinguished articulations: a large, medial one for the scaphoid and a smaller one, lateral, triangular and concave anteroposteriorly for the lunate. In posterior view, the extension of the distal articulation for the scaphoid is large and well developed, but wider than high. The distolateral contact area for the ulna is large.
COMPARISON. The radius of R. velaunum is unknown. The proximal facets for the ulna from the radii of R. filholi are like those of R. romani . The radius of R. filholi mostly differs from R. romani by its deep and wide gutter for the m. extensor carpi on the anterior side of the distal extremity, which is very shallow on the radii from ‘Marseille’ and Gaimersheim. This deep gutter is also present on a radius from Espenhain (BSPG- 2008-I- 44), also attributed to R. filholi , although the total length of the bone is much smaller than in Villebramar (around 30 cm versus 38 to 41 cm in Villebramar). However, a very deep gutter is also present on the radius of Diaceratherium tomerdingense (SMNS-16154), whereas it is shallow on a hand of D. lemanense (Pomel, 1853) from Gannat (MNHN-LIM-598). Therefore, there seems to be variation within this character, even among species of a same genus. Finally, Ménouret & Guérin (2009) referred the radius from ‘Marseille’ to Diaceratherium massiliae because they considered that in “ R. filholi it is the external humeral facet that is the most developed ( Brunet 1979: pl. 21)” [translated by the authors], which is actually incorrect. Brunet (1979: 138) in fact states in the description of the material from Villebramar that “[the proximal articulation surface] is composed of two glenoid cavities (a large internal one, long, weakly concave; a smaller external one, thinner and more concave, pl XXIc)” [translated by the authors], which is also the case in the radii described here. The radius of D. lamilloquense from Castelmaurou (UM CAM-22) differs by the very concave posterior border of the proximo-medial articulation surface in proximal view, whereas it is straight in Ronzotherium .
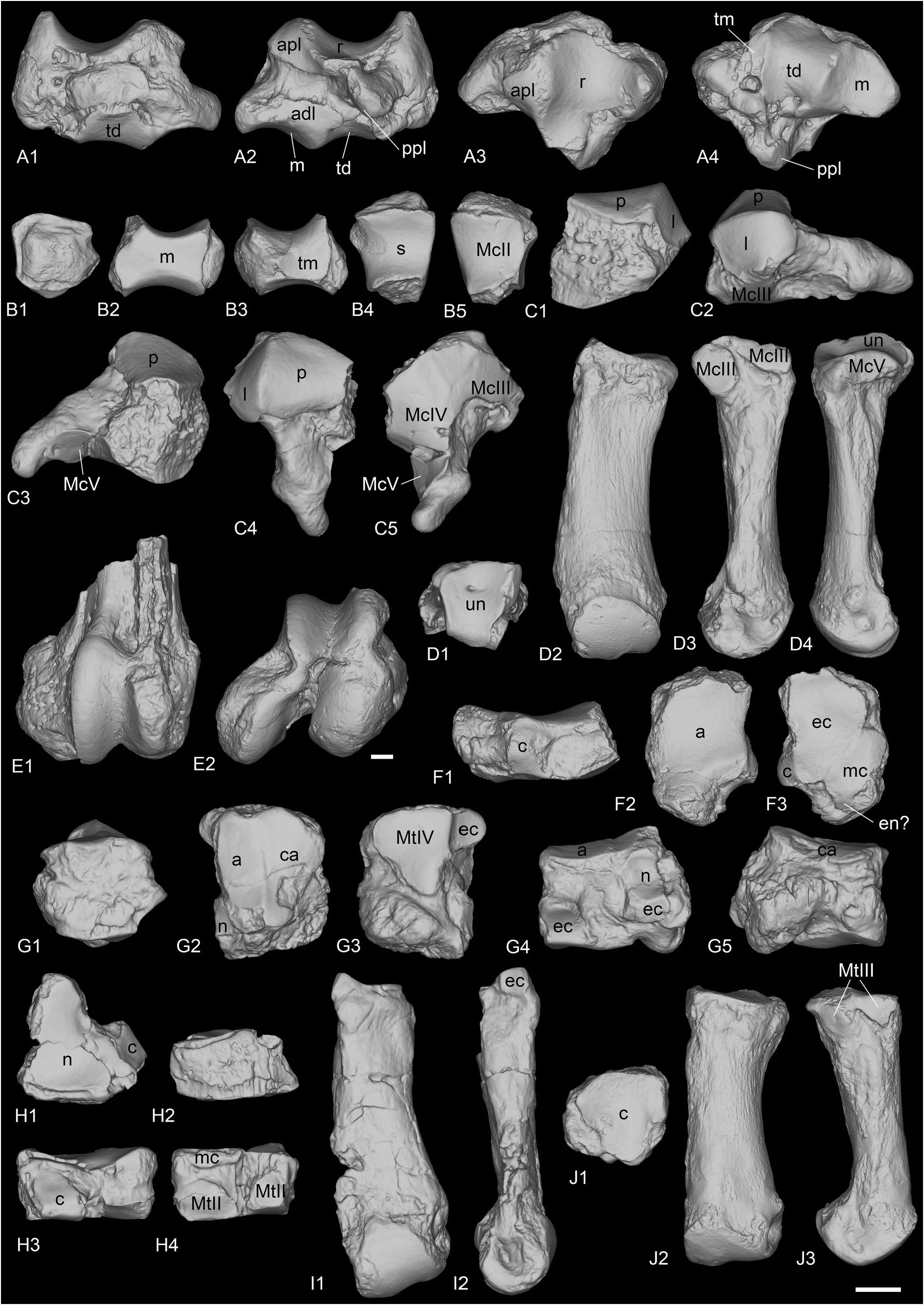
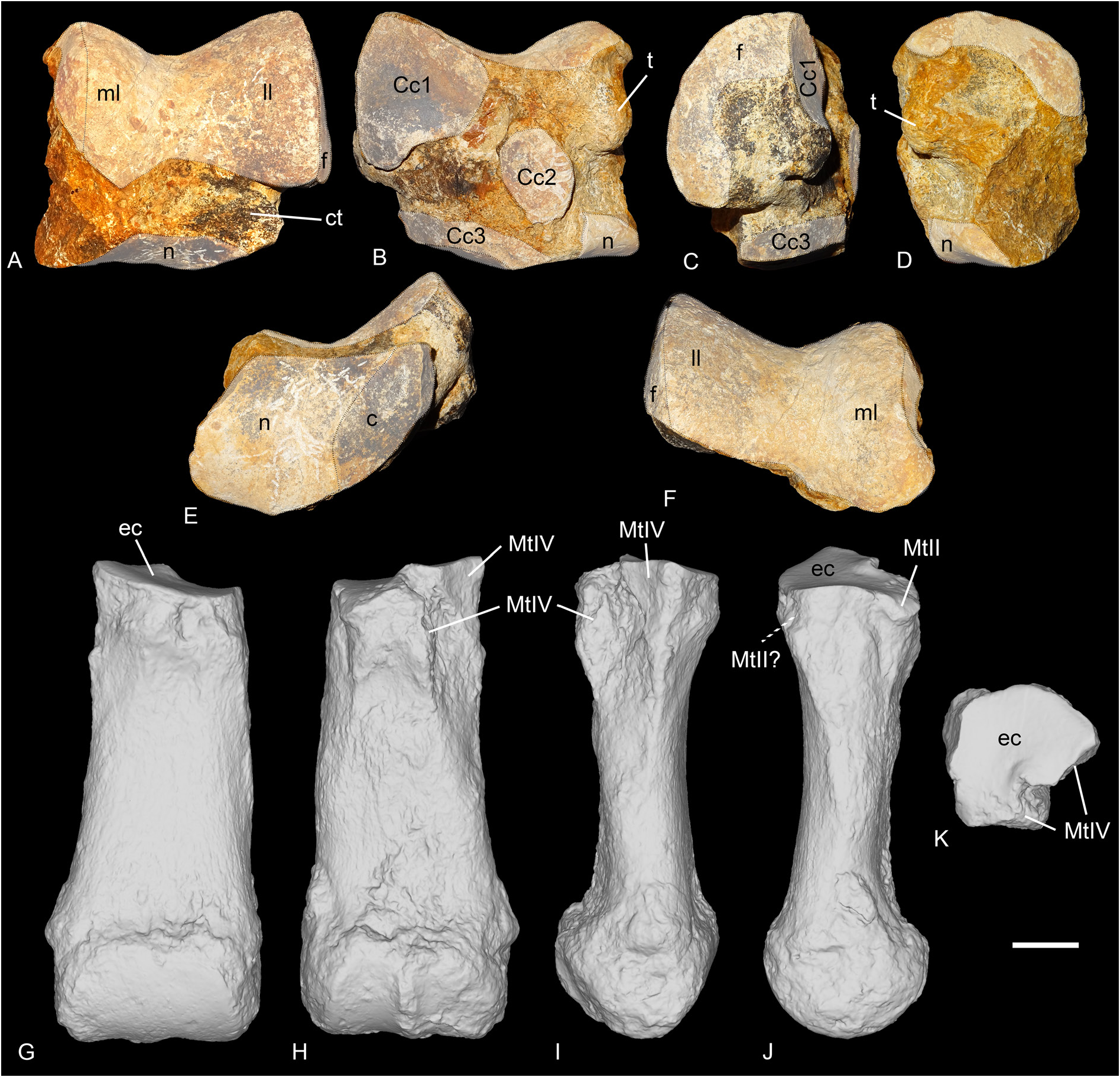
SCAPHOID. It is preserved in ‘Marseille’ (FSL-520285, Fig. 18A View Fig ), Gaimersheim (BSPG collection, Fig. 19E–H View Fig ) and Rickenbach (NMO-I5-62, Fig. 21G View Fig ). All three specimens are very well preserved and almost identical. The proximal articulation for the radius is triangular, and concave anteroposteriorly. Posterodistal to the proximal articulation, there is a large lateromedially elongated tuberosity on which occurs an articulation for the lunate (the postero-proximal articulation for the lunate sensu Antoine (2002). This articulation for the lunate is fused with the anteroproximal facet for the lunate on the scaphoids from Gaimersheim and Rickenbach, but they are partly separated by a shallow groove on the specimen from ‘Marseille’. The anteroproximal articulation for the lunate is band-shaped and separated from the anterodistal articulation for the lunate by a large and deep groove for ligaments, extending anteroposteriorly, below the proximal tuberosity. On the lateral side, the anterodistal articulation for the lunate is anteroposteriorly elongated, band-shaped and almost fused with the distal magnum facet. The distal articulations for the magnum (anteriorly) and the trapezoid (median) are concave and almost equal-sized. Posterior to these two facets, there is a small articulation facet for the trapezium on the specimens from ‘Marseille’ and Gaimersheim, that seems to be absent or fused with the trapezoid facet on the specimen from Rickenbach. The anterior and posterior heights of the scaphoid are equal. In medial view, the trapezoid facet is prominent and high, whereas the other facets are not visible.
COMPARISON. The scaphoid of R. velaunum shows many similarities with that of R. romani : the anterior and posterior heights are equal, the proximal articulation for the radius is triangular, the anterodistal facet for the magnum is very concave and the facet for the trapezoid is extended anteriorly. However, it differs by a smaller size, a better development of the trapezium facet and a less well developed tuberosity below the proximal articulation. The scaphoid of R. filholi is also similar to the scaphoid of R. romani , especially in its development of the distolateral apophysis (bearing the magnum facet). The scaphoid from Quercy attributed to? R. filholi is almost identical, and also shows the typical fusion of the anteroproximal and postero-proximal facets for the lunate. The scaphoids of Diaceratherium asphaltense ( Depéret & Douxami, 1902) from Pyrimont (FSL213008), D. lamilloquense ( Michel 1983; Duranthon 1990), D. aginense from Laugnac (MHNM.1996.17.94) and D. aurelianense (Nouel, 1866) from Neuville-aux-Bois (MHNB41.2018.0.282, -.384 and -.866) differ from R. romani by a higher posterior height compared to the anterior, a more convex dorsal border in proximal view, a flattened articulation for the magnum, a much more concave articulation for the trapezoid and a larger articulation for the trapezium. Furthermore, D. aurelianense also greatly differs by the deep and wide groove separating the anteroproximal and postero-proximal facets for the lunate, as in Pleuroceros blanfordi ( Antoine et al. 2010: fig. 6) or Teleoceras aepysoma ( Short et al. 2019: fig. 45), whereas in Ronzotherium they are either completely fused or partly connected, although in D. lamilloquense from La Milloque and Castelmaurou they also seem to be fused ( Michel 1983; Duranthon 1990). In D. asphaltense , this postero-proximal facet seems absent.
LUNATE. It is only known from Rickenbach (NMO-I7/115 and NMB-Ri-27, Fig. 21H View Fig ). One is complete (NMO-I7/115) but the other is broken. The proximal articulation for the radius is large and convex anteroposteriorly. It occupies the whole anteroproximal border, there is no articulation with the ulna. In proximal view, there is a drop-like posterior extension of the radius facet on the medial border. In anterior view, the proximal border is much wider than the distal part. The anterior side is deeply keeled. There are two medial articulations, two lateral and two distal. In lateral view, there is only one proximal articulation facet for the scaphoid, which occupies the whole proximal border, formed by the fusion of the anteroproximal and postero-proximal facets, as on the scaphoid. A shallow groove separates the proximal facet from the distal one. This distal facet for the scaphoid is high, almost triangular and restricted to the anterior portion of the lunate. In medial view, the proximal and distal articulations for the pyramidal are rather small, but they are not in the same plane, the proximal one is more medially displaced. The proximal one is a half oval, whereas the distal one is band-shaped and posteriorly displaced. In distal view, there are two large articulation facets: an anterior one for the unciform, and a distal one for the magnum, very concave, with a thin band-shaped anterior elongation separating the
scaphoid facet from the unciform facet. The unciform facet occupies almost all of the distal border of the bone in anterior view and is nearly horizontal. The magnum facet is very small in anterior view and makes a very weak angle with the distal scaphoid facet.
COMPARISON. Although the lunate of R. velaunum from Ronzon is not fully extracted from the sediment, the visible part of the bone is similar to the lunate from Rickenbach. The proximal articulation for the radius is very wide and has a posterior extension. The posterior tuberosity is larger and wider than in Rickenbach. On the medial side, the two facets for the pyramidal are not in the same plane either, though on the specimen from Ronzon, the distal facet is much larger. These characters are also found in the lunate of R. filholi from Villebramar ( Brunet 1979). However, the lunates of R. velaunum and R. filholi (both from Villebramar and Quercy) differ by the presence of a shallow groove separating the anteroproximal facet for the scaphoid from the postero-proximal one. The lunates of Diaceratherium tomerdingense (SMNS-16157c), D. aurelianense ( Cerdeño 1993) , D. aginense (MHNM.1996.17.21) and D. asphaltense (FSL-213008) differ by their reduced posterior tuberosity in proximal view, the reduced or absent posterior extension of the proximal facet for the radius and the much more proximo-distally compressed anterior side. They also mostly differ by the wide groove separating the anteroproximal facet for the scaphoid from the postero-proximal one as well as the larger anterior portion of the facet for the magnum (it is almost as large as the distal pyramidal facet). In D. lamilloquense from Castelmaurou (TLM.PAL.2014.0.2571), this postero-proximal facet for the scaphoid is either absent or separated from the anterior by a large groove, as in other diaceratheres, and the magnum facet is also rather large anteriorly. On the preserved hand of D. lemanense from Gannat (MNHN-LIM-598), the anterior portion of the magnum facet is also very large, as in other diaceratheres but the scaphoid facets are not visible.
PYRAMIDAL. It is only known from Rickenbach (NMO-I11-82, Fig. 21I View Fig ) and almost complete. The proximal ulna facet is large, saddle-shaped, concave anteroposteriorly, transversally convex and medially elongated. It contacts the long band-shaped postero-proximal pisiform facet. There are two lateral facets for the lunate: the proximal one is half-oval, and the distal one is asymmetrical. They are separated by a wide and shallow groove, and are not exactly in the same plane, in the same way as the two corresponding facets on the lunate. Furthermore, their size, shape and position also fit with it. The distal articulation for the unciform is triangular in distal view and concave anteroposteriorly.
COMPARISON. This specimen is almost identical to the pyramidal of R. velaunum MNHN.F.RZN. 502, both in size and morphology. It only differs by a larger distal facet for the lunate, and a deeper groove between the two facets for the lunate. The pyramidal of R. filholi is overall also very similar but shows a deeper groove between the two lunate facets, as well as a tubercle on the posterior side, below the unciform facet, which is absent in R. romani . The pyramidals of D. tomerdingense (SMNS-16157d) and D. aginense (MHNM.1996.17.20) differ however by very different proportions: the anterior side is lower and more anteroposteriorly elongated, and the medial side, corresponding to the lunate, is extremely reduced proximo-dorsally. They also differ by their less elongated and drop-shaped facet for
the pisiform and a laterally reduced facet for the ulna. On the medial side, the two facets for the lunate are very small, band-shaped and anteroposteriorly elongated, contrary to the pyramidal of Ronzotherium .
TRAPEZOID. Two trapezoids are preserved from ‘Marseille’ (FSL-9501 and FSL-520283, Fig. 18B View Fig ). In anterior view, they are wider than high.The proximal border is sigmoid on the specimens from ‘Marseille’. The magnum facet occupies the whole lateral side, while the medial side is partly occupied by the extension of the scaphoid facet, and by a subtriangular medio-distal articulation for the trapezium. The proximal side is fully occupied by the anteroposteriorly concave scaphoid facet. The distal articulation for the McII is anteroposteriorly concave.
COMPARISON. The only other known trapezoid of Ronzotherium belongs to R. filholi from Villebramar ( Brunet 1979). It differs by a flattened distal articulation for the McII and a concave proximal border in anterior view. This trapezoid is also smaller than those that we refer here to R. romani , especially anteroposteriorly. Because we lack comparative specimens, especially with the type species R. velaunum , we can only tentatively attribute these trapezoids to R. romani .
MAGNUM. It is preserved from Gaimersheim (BSPG collection, Fig. 19I–L View Fig ) and Rickenbach (NMO-H10/110, Fig. 21J View Fig ). The specimens are partly broken. In anterior view, the proximal border is straight. The anterior side is wider than high. The proximal apophysis is wide, high and very convex. This apophysis is laterally bordered by a long band-shaped articulation for the lunate, that completely fuses anteriorly with the small unciform facet. The proximomedial facet for the scaphoid is larger and concave anteroposteriorly. This facet is very poorly distinguished from the medial facet for the trapezoid. This latter facet is longer than high, and its morphology would fit the shape of the corresponding facet on the trapezoid from ‘Marseille’. The trapezoid facet is separated from the medio-distal McII facet by a ridge and by a very short and shallow notch anteriorly. This facet is much longer than high, flat and its distal border is very concave in medial view. On the distal side, the McIII facet is large, trapezoidal, longer than wide and very concave anteroposteriorly. The posterior tuberosity of the magnum is short and straight in Rickenbach.
COMPARISON. The magnum of R. velaunum PUY. 2004.6.263.RON differs from the specimens from Rickenbach and Gaimersheim by its narrower proximal apophysis. The magnum of R. filholi also differs from R. romani by its higher and narrower anterior side. The magnum of Diaceratherium asphaltense (FSL-213008) only differs by a slightly longer and straighter posterior tuberosity, and by a shorter proximal contact between the trapezoid and scaphoid facets.
UNCIFORM. Three unciforms (FSL-520289, FSL-520282 and NMB-Mar-865, Fig. 18C View Fig ) are preserved from ‘Marseille’ according to Ménouret & Guérin (2009) but we only could recover the specimen NMB-Mar-865. It is almost complete, only a small part of the anterolateral side is missing. In anterior view, the two proximal facets for the pyramidal and the lunate are visible. In proximal view, the posterior expansion of the pyramidal facet was probably absent, and the pyramidal and McV facets were probably separated. The McV facet is large, very concave and located posteriorly. The posterior apophysis is thin, curved and ‘hook-shaped’. In distal view, the McIII and McIV facets are almost undistinguishable, forming a single large convex facet.
COMPARISON. Only one other unciform of Ronzotherium is known, from Ronzon, but it is very incomplete. However, from the remaining part, the dimensions are very similar to those of R. romani , and no characters permit to distinguish them. In contrast, the unciform of Diaceratherium tomerdingense (SMNS-16157e) strongly differs from that of R. romani by its very wide and flattened posterior apophysis, its larger McV facet contacting the pyramidal facet, the much thinner and elongated McIII facet that is well distinguished from the McIV facet and the anteroposteriorly concave McIV facet. The unciform of D. lemanense (MNHN-LIM-598) also has a very wide posterior apophysis and a connection between the McV and pyramidal facets. The unciform of D. aginense (MHNM.1996.17.98) shows a similar wide posterior apophysis, but the contact between the pyramidal and the McV facets is absent.
MCIII. The McIII is overall very badly preserved. In ‘Marseille’, UPM-13667 is incomplete and poorly preserved, whereas FSL-9505 and FSL-520281 are two proximal extremities (none found in collection). In Gaimersheim (BSPG collection, Fig. 19M, P–R, U View Fig ), it is very broken and incomplete. The anterior McII facet is large and semi-circular. The posterior McII facet seems absent. The magnum facet is convex anteroposteriorly. On the lateral side, only the posterior McIV facet is preserved, but it was separated from the anterior by a shallow groove.
COMPARISON. The McIII of R. velaunum is unknown. One McIII of R. filholi is preserved in Möhren 7 (BSPG-1969-XXIV) and differs by the much smaller anterior McII facet. However, the posterior McIV facet is similar and also separated by a shallow groove from the anterior. This groove is larger in Villebramar ( Brunet 1979). The McIII of Diaceratherium cannot be distinguished based on these characters.
MCIV. It is preserved from ‘Marseille’ (FSL-520287, NMB-Mar-863 and NMB-Mar-864, Fig. 18D View Fig ), Gaimersheim (BSPG collection, Fig. 19N–O, S–T, V View Fig ) and Rickenbach (NMO-I8/117, Fig. 21K View Fig ). In proximal view, the proximal side is lozenge to triangular-shaped. The articulation for the unciform is almost flat anteroposteriorly, but slightly concave lateromedially. On the lateral side, the articulation for the McV is long and low, except on the specimen from Rickenbach where it is reduced and circular. The rugosity of the contact surface for the McV on the lateral border occupies almost half of the diaphysis proximally. On the medial side, two large facets articulate with the McIII (broken on the specimen from Gaimersheim): one is band-shaped and anteroposteriorly elongated on the anteroproximal border, and the other oval-shaped, posterior and separated from it by a groove. These two facets are almost in the same vertical plane. In posterior view, the specimen from Rickenbach differs from the others by its very deep fossa, just above the distal articulation.
COMPARISON. The McIV of Diaceratherium tomerdingense (SMNS-16155b) strongly differs by its reduced length (only 9.5 cm), its convex medial border of the diaphysis with a prominent rugose tuberosity on the anteroproximal part of the diaphysis, the anterior McIII facet contacting the posterior one and the deep incision of the posterior border of the unciform facet in proximal view. The McIV of D. asphaltense (FSL-213012) also differs by its convex medial border of the diaphysis with a prominent rugose tuberosity on the anteroproximal part of the diaphysis, but the two McIII facets are separated, and the unciform facet is not incised.
SACRUM. A sacrum is preserved from Poillat (MJSN-BEU-001-280). It is quite well preserved and is formed by the fusion of five sacral vertebrae. The neural spines are not fused together and there are four dorsal and ventral sacral foramina on each side.
COMPARISON. Because of the rarity of the preservation of the sacrum, no comparison can be made, either with other ronzotheres or with Diaceratherium .
FEMUR. Only a very poorly preserved but subcomplete femur is known from the locality of Poillat (MJSN-POI007-80). The distal articulation is only known in ‘Marseille’ (NMB-Mar-828, Fig. 18E View Fig ). The head of the femur is rounded, and the fovea capitis is deep. The smaller trochanter is only preserved on the specimen from Poillat, and it is very small. The third trochanter, the medial condyle and the medial lip of the trochlea are not preserved. The lateral condyle is protruding posteriorly, far behind the diaphysis and the lateral epicondyle is present but not very developed laterally.
COMPARISON. There are almost no characters preserved that permit to distinguish the femur of R. romani from other ronzotheres, or from Diaceratherium but it is overall very similar to the femur of R. velaunum from Ronzon.
TIBIA. It is only known from Gaimersheim (BSPG collection), and only its medial half is preserved. The medial articulation surface is circular and concave in proximal view and the medial intercondylar tubercle is present. The medial border of the diaphysis is slightly concave. In the distal part, the mediodistal gutter is not preserved and the posterior apophysis is broken. The ridge delimitating the two distal condyles is wide and low. The fibula is unknown.
COMPARISON. Based on what is left from this tibia, it only seems to differ from Ronzotherium velaunum (PUY.2004.6.260.RON and PUY.2004.6.261.RON) in having a larger size. However, it is slightly shorter than the tibiae of R. filholi from Villebramar. There are too few characters visible on the tibia from Gaimersheim to compare it with those of Diaceratherium , which also have a very similar size.
ASTRAGALUS. It is preserved from Gaimersheim only (BSPG collection, Fig. 20A–F View Fig ) and slightly eroded but complete. It is wider than high (TD>H) and its APD/H ratio is high (around 0.78). On the lateral side, the fibula facet is large, flat and vertical. In anterior view, the lateral lip is larger than the medial one, and the groove between the two lips is wide. The collum tali is very high, the two lips of the trochlea do not contact the distal articulation at all. The distal articulation for the navicular is concave in anterior view. In distal view, this articulation is a parallelogram, and it bears a proximal extension on the posterior side of the astragalus. Lateral to the articulation for the navicular, there is a smaller, almost flat and anteroposteriorly elongated facet for the cuboid. This facet is posteriorly broken, and the posterior stop is thus not preserved. In distal view, the trochlea is oblique compared to the distal articulation. In proximal view, the posterior border of the trochlea is sinuous. In posterior view, the three facets for the calcaneum, Cc1, Cc2 and Cc3, are distinct. The Cc1 facet is the largest and it bears a low and wide distal extension on the lateral side. It is rather triangular, and almost flat in lateral view. It is separated from the Cc2 by a deep proximal fossa, and from the Cc3 facet by a wide groove. The Cc2 facet is almost contacting the Cc3 facet by a very thin bridge and it is oval-shaped and slightly proximodistally elongated. There is a strong, rounded tuberosity medial to this Cc2 facet and separated by a large and deep proximodistal groove. Distally, the Cc3 facet is low and band-shaped, but partly eroded. The medio-distal tubercle of the astragalus is broken.
COMPARISON. The astragalus of R. velaunum (PUY.2004.6.1770.RON) shares with the astragalus of R. romani the very high collum tali and the absence of contact between the trochlea and the distal border, the large lateral lip compared to the medial one, the wide groove between the two lips of the trochlea, the large and flat fibula facet, the transversally concave distal navicular facet and the oblique trochlea compared with the distal articulation, in distal view.These same characters are also found on the astragalus of R. filholi from Villebramar. Unfortunately, the posterior side of the astragalus of R. velaunum is still in sediment. Another astragalus (MNHN.LIM7) attributed to R. filholi from Bournoncle-Saint-Pierre is also very similar but it shows an even more laterally offset lateral lip of the trochlea. It shares, however, the deep proximal fossa separating the Cc1 and Cc2 facets, the oval-shaped and proximodistally elongated Cc2 facet, and the band-shaped Cc3 facet. However, on this specimen, the Cc1 facet is very concave in lateral view and the Cc2 facet is connected to the Cc3 by a very wide band, contrary to the specimens from Gaimersheim and Villebramar (flattened sagittally). Also, the distal extension of the Cc1 facet is long, thin and drop-shaped. The astragalus of Diaceratherium lemanense from Gannat (NMB-Gn-158), as well as the astragali of D. aginense from Laugnac (MHNM.1996.17.41, -.55 and -.77) differ from the astragalus of R. romani in having a more visible and more concave facet for the navicular in anterior view, a lower height, a lower collum tali, more rounded lips of the trochlea, a larger and circular Cc2 facet, completely independent Cc2 and Cc3 facets, a concave Cc1 facet in lateral view and a reduced distal extension of the Cc1 facet.
The calcaneum, meso- and entocuneiform remain unknown for R. romani .
NAVICULAR. It is only preserved in ‘Marseille’ (NMB-Mar-847e, Fig. 18F View Fig ). It is quite large, longer than wide and pretty high. The proximal articulation for the astragal is slightly anteroposteriorly concave and occupies the whole anterior side. The distal side is occupied by two poorly distinguished facets: a large, anterolateral and almost triangular one for the ectocuneiform, and a smaller one, rectangular and located postero-medially, for the mesocuneiform. There is possibly a third very small facet for the entocuneiform, but it cannot be distinguished from the mesocuneiform facet. On the lateral side, there is a single posterior and convex articulation for the cuboid. The cross-section of the navicular is lozengeshaped.
COMPARISON. The navicular of Ronzotherium velaunum is not preserved from Ronzon, but one specimen is known in Haag 2 (unnumbered in BSPG collection). It shares a very similar shape in proximal view, with a distinct posterior notch, as well as the absence of an anterior cuboid facet and a similar concavity in lateral view. The only other known navicular of Ronzotherium belongs to R. filholi from Villebramar ( Brunet 1979). Its morphology is very similar and it basically only differs by its slightly smaller size. The navicular of D. lamilloquense ( Michel 1983) differs by its shape, it is as long as wide, and by the presence of an anterolateral facet for the cuboid. It also differs by its distal facets: the mesocuneiform facet is triangular and slightly convex, the entocuneiform facet is oblique, and the three cuneiform facets are distinguishable and separated. The navicular of D. aginense is also as wide as long and differs by its distal articulation surfaces. The navicular of D. aurelianense ( Cerdeño 1993) also differs by its overall shape, by the two facets for the cuboid, and by a strong angle between the distal ento- and mesocuneiform facets.
ECTOCUNEIFORM. Only one ectocuneiform is known for R. romani , from ‘Marseille’ (NMB-Mar-735, Fig. 18H View Fig ). The proximal articulation for the navicular is roughly triangular, concave and longer than wide. The postero-lateral process is absent. The lateral side bears two facets for the cuboid, a large and oblique anterodistal one, and a smaller postero-proximal one. The groove separating these two is rather deep. On the medial side, the mesocuneiform facet is thin, low, elongated and located posteroproximally whereas the two distal articulations for the MtII are rather large. The anterior one is concave whereas the posterior one is larger and convex. The distal articulation for the MtIII is triangular. In anterior view, the distal border is sinusoidal.
COMPARISON. The ectocuneiform of R. velaunum (PUY.2004.6.577.RON) slightly differs by its smaller and vertical anterodistal facet for the cuboid and its smaller posterior facet for the MtII. The ectocuneiform of R. filholi from Villebramar ( Brunet 1979) only differs by its slightly smaller size. The ectocuneiform of Diaceratherium greatly differs from Ronzotherium . The ectocuneiform of D. lamilloquense from La Milloque ( Michel 1983) differs by the presence of a facet for the MtIV below the anterior facet for the cuboid and by a less elongated and triangular facet for the mesocuneiform, that is located more anteriorly than in Ronzotherium . The ectocuneiform of D. lamilloquense from Castelmaurou ( Duranthon 1990) differs by the presence of a third articulation facet for the cuboid. The one of D. aurelianense from Artenay differs by the fusion of the two distal facets for the MtII ( Cerdeño 1993).
CUBOID. It is preserved in ‘Marseille’ (FSL-9528 and NMB-Mar-847d, Fig. 18G View Fig ). The anterior side is approximately as high as wide. In anterior view, the proximal articulation is posteriorly elevated. In proximal view, the posterior apophysis is almost not visible, the proximal side is occupied almost exclusively by the two articulation surfaces, for the astragalus on the medial side, and for the calcaneus laterally. The proximal side is trapezoid and the astragalus and calcaneal facets are almost equal-sized. On the medial side, the postero-proximal and elongated facet for the navicular is concave and contacts the small, square and postero-mesial facet for the ectocuneiform. The navicular facet bears a thin extension up to the anterior border, bordering the astragalus facet. The small anterodistal facet for the ectocuneiform is separated from the posterior one by a wide groove. On one specimen (NMB-Mar- 847d), this anterodistal facet is very developed and deeply concave, with a strong medial extension, that is not visible on the other specimen. There is no articulation facet on the lateral side, but a large and deep groove, obliquely and forward oriented, which serves as a ‘slideway’ for the tendon of the m. fibularis longus and isolates the posterior apophysis of the cuboid from the main body of the bone. In distal view, the distal articulation for the MtIV is almost flat and triangular.
COMPARISON. The cuboid of R. velaunum (PUY.2004.6.1309.RON and PUY.2004.6.268.RON) differs from that of R. romani by its smaller and oval-shaped distal articulation for the MtIV, by its shallow groove separating the proximal calcaneal facet from the astragalus one, and by its slightly shorter proximal articulation. All other characters are overall very similar to those of R. romani . The cuboid of R. filholi from Villebramar is poorly preserved, but it differs nonetheless by its slightly shorter posterior height, at the level of the posterior apophysis. The cuboid from the Quercy (NMB-QE-362) tentatively referred to? R. filholi differs by its very different morphology of the anterior side, the absence of ridge separating the proximal astragalus and calcaneal facets and its more posteriorly elevated proximal articulation, but resembles R. romani by its very similar medial articulations for the ectocuneiform and navicular. The cuboid of Diaceratherium asphaltense (FSL-213014) from Pyrimont-Challonges greatly differs from R. romani by its proportions, dimensions and morphology (see Depéret & Douxami 1902: pl. XXIX, fig. 7). It differs by the presence of an isolated anteroproximal facet for the lunate. The height of the anterior side is much smaller than in R. romani , whereas its width is similar. However, it is much higher posteriorly than the cuboids of R. romani , because of the very high apophysis, and the strong posterior elevation of the proximal surface. The proximal articulation is rectangular in proximal view and the posterior apophysis is very visible posterior to this articulation. The distal articulation for the MtIV is transversally convex and concave anteroposteriorly. The lateral groove for the tendons is very shallow. Another cuboid from Castelmaurou (TLM.PAL.2014.0.2563) attributed to D. lamilloquense ( Duranthon 1990) also shares the same characters as D. asphaltense , and especially the isolated anteroproximal facet for the lunate, which is always absent in ronzotheres. The presence of this facet thus seems to be a diagnostic character differentiating Diaceratherium from Ronzotherium .
MTII. One MtII of R. romani is preserved from ‘Marseille’ (NMB-Mar-847a, Fig. 18I View Fig ) but was originally attributed to a McII of “ Diaceratherium ” massiliae and another from Rickenbach (NMB-UM-2565, Fig. 21L View Fig ). It is partly broken proximally. In anterior view, the proximal articulation for the mesocuneiform is concave. The diaphysis is curved towards the medial side and is very widened distally.Antero-laterally, there is no anterior articulation for the MtIII, only a single small facet for the ectocuneiform. A groove separates this facet from the two posterior facets (not preserved on the specimen from ‘Marseille’): one is for the ectocuneiform, the other below, is for the MtIII. The ectocuneiform facet is large and oblique, whereas the MtIII facet is thin and elongated.
COMPARISON. The MtII of R. filholi from Villebramar ( Brunet 1979) and Möhren 7 (BSPG-1969- XXIV-73) differ by the presence of an anterior facet for the MtIII, below the ectocuneiform facet, and by their gracility. The MtII of D. lemanense from Wischberg ( Jame et al. 2019) differs in being more gracile, but also in having a very large posterior facet for the ectocuneiform, an anterior facet for the MtIII and an elongated posteromedial entocuneiform facet.
MTIII. It is only preserved from Gaimersheim (BSPG- 1952-II, Fig. 20G–K View Fig ). It differs drastically from MtIII of R. filholi from Villebramar by its robustness. The proximal part is slightly broken medially and laterally. The proximal articulation for the ectocuneiform is roughly trapezoid, with a lateral notch separating the two facets for the MtIV, and it is as wide as long. It is slightly bulged at the level of this notch. There is no facet for the cuboid. In anterior view, the proximal border is straight and oblique and there is a marked distal widening of the diaphysis towards the distal articulation. In medial view, the anterior articulation for the MtII is broken but may have been absent, and the posterior is small and poorly differentiated from the proximal articulation. In lateral view, the anterior articulation for the MtIV is large and triangular, whereas the posterior is poorly preserved. They are separated by a deep groove. The distal keel is quite smooth but still visible in anterior view, and there is no distal tubercle on the posterior side. The insertions of the m. interossei are long on the medial and lateral sides (they extend beyond the middle of the diaphysis).
COMPARISON. The MtIII of R. velaunum is poorly preserved, and it differs from that of R. romani by its greater length, even though its width is quite similar. It also shares with R. romani a distal widening of the diaphysis and a smooth distal keel of the articulation. The MtIII of R. filholi from Villebramar differs from that of R. romani by its higher gracility, but it shows a similar distal widening of the diaphysis. As in R. romani , the proximal border is straight and oblique in anterior view and the distal keel is smooth. Another MtIII from Möhren 7 (BSPG-1969-XXIV-156) is quite similar to that of R. romani , as it shares the distal widening of the diaphysis, the absence of a posterior facet for the MtII, the presence of a posterior articulation for the MtII and the similar shape of the proximal side. Although their length is almost equal, the MtIII of R. romani is much wider. The MtIII of Diaceratherium asphaltense (FSL-213016) differs by its smaller size and its reduced width, compared to the MtIII of R. romani . The shape of the proximal side in anterior view is also quite different, it is triangular. It also differs by its shorter insertion for the m. interossei, the absence of a posterior facet for the MtII and the absence of distal widening of the diaphysis. In proximal view, the anterior border of the proximal articulation is straight, and it is also slightly less oblique in anterior view. The MtIII of D. lamilloquense from Castelmaurou (TLM.PAL.2014.0.2564) differs from that of R. romani by its much thinner diaphysis, its concave proximal articulation in anterior view, the absence of a posterior facet for the MtII, and its proximal side being much wider than long in proximal view.
MTIV. It is only preserved from ‘Marseille’ (FSL-520286, Fig. 18J View Fig ), and it is complete. As for the MtIII, it is also more robust than the MtIV of R. filholi from Villebramar. The proximal articulation for the cuboid is roughly triangular, with a small notch on the medial side between the two facets for the MtIII. The postero-proximal tuberosity is pad-shaped and continuous. On the medial side, the two facets for the MtIII are rather large, and separated by a narrow groove, than runs from the proximal side to the anterior side. The anterior MtIII facet is triangular while the posterior one is less proximal, and oval-shaped. There is no posterior tubercle. The MtV facet is absent. By virtually articulating the 3D models of this MtIV to the MtIII from Gaimersheim, their morphologies would both match very well: the length of the anterior MtIII/MtIV facet is identical, and the groove is located at the same position; on the diaphysis, the insertions for the m. interossei extend up to the same level.
COMPARISON. The MtIV of R. velaunum from Ronzon is lost. The MtIV of R. filholi from Villebramar ( Brunet 1979) differs by its dimensions, the concave proximal facet for the cuboid, and the much wider groove separating the two MtIII facets. The MtIV of Diaceratherium lamilloquense ( Duranthon 1990) also differs by its dimensions, by the concave proximal facet for the cuboid, by the much wider groove separating the two MtIII facets and by the 90° angle between these two. The MtIV of D. aginense from Laugnac (de Bonis 1973: fig. 34a) further differs by the presence of an anterior ectocuneiform facet, by a reduced postero-proximal tuberosity and by the very different shape (triangular) of the proximal side.
FINAL REMARKS. All these newly identified postcranial remains considerably change our view of the species Ronzotherium romani . Prior to this study, only scarce remains were identified, and this species was believed to resemble its closely-related species R. filholi , by being rather medium-sized, gracile and cursorial. Based on this wrong premise, large and robust postcranial rhinocerotid remains from ‘Marseille’ were not assigned to the co-occurring R. romani . Indeed, the species, “ Diaceratherium ” massiliae was named based on these short and robust postcranials, as it was not conceivable to consider they would document any representatives of Ronzotherium ( Ménouret & Guérin 2009) .Yet, by comparing postcranial remains to other remains of Ronzotherium , especially to those of R. velaunum for which the postcranial skeleton is well preserved, we show that all of them can be assigned to Ronzotherium , instead of Diaceratherium . In particular, the postcranial skeleton of Ronzotherium romani differs from Diaceratherium by:
- the lower fossa olecrani of the humerus in posterior view and the constricted condyles in anterior
view;
- the equal posterior and anterior heights of the scaphoid, a less convex dorsal border in proximal
view, a concave distal articulation for the magnum, a less concave articulation for the trapezoid and a
smaller articulation for the trapezium;
- the large posterior tuberosity of the lunate in proximal view, the developed posterior extension of the
proximal facet for the radius, the higher anterior side in anterior view, the shallower groove separating
the anteroproximal facet for the scaphoid from the postero-proximal, as well as the reduced anterior
portion of the facet for the magnum (it is almost as large as the distal pyramidal facet);
- the higher anterior side of the pyramidal, the more elongated facet for the pisiform, the developed
facet for the ulna and the larger facets for the lunate;
- the shorter and straight posterior tuberosity of the magnum and a shorter proximal contact between
the trapezoid and scaphoid facets;
- the thin, curved and ‘hook-shaped’ posterior apophysis of the unciform, the larger McIII facet, poorly
distinguished from the McIV facet and the convex McIV facet;
- the absence of rugose tuberosity on the anteroproximal part of the diaphysis of the McIV, the straighter
medial border of the diaphysis and its more reduced robustness;
- the less concave facet for the navicular on the atragalus, the higer collum tali, the smaller Cc2 facet,
the contact between the Cc2 and Cc3 facets, a flatter Cc1 facet in lateral view and the large distal
extension of the Cc1 facet;
- the global size and shape of the navicular, longer than wide, and its single posterior articulation for
the cuboid;
- the disposition of the facets of the ectocuneiform;
- the absence of anteroproximal facet for the lunate on the cuboid and the trapezoid proximal side;
- the more robust MtII, without anterior facet for the MtIII;
- the wider diaphysis of the MtIII and the different shape of its proximal articulation;
- the dimensions of the MtIV and the shallower groove separating the two MtIII facets.
Accordingly, Diaceratherium massiliae Ménouret & Guérin, 2009 should be considered as a junior synonym of Ronzotherium romani Kretzoi, 1940 .
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Ronzotherium romani Kretzoi, 1940
| Tissier, Jérémy, Antoine, Pierre-Olivier & Becker, Damien 2021 |
Diaceratherium massiliae Ménouret & Guérin, 2009: 314–323
| Menouret B. & Guerin C. 2009: 323 |
Ronzotherium romani
| Brunet M. 1979: 154 |
Praeaceratherium kerschneri
| Spillmann F. 1969: 253 |
Ronzotherium romani
| Kretzoi M. 1940: 91 |
Acerotherium filholi
| Roman F. 1910: 1559 |