
Sphaerodromia kendalli (Alcock & Anderson, 1894)
|
publication ID |
https://doi.org/10.5281/zenodo.5397969 |
|
persistent identifier |
https://treatment.plazi.org/id/C5482F17-901D-FFC5-C416-F99CFBFFFE2E |
|
treatment provided by |
Marcus |
|
scientific name |
Sphaerodromia kendalli (Alcock & Anderson, 1894) |
| status |
|
Sphaerodromia kendalli (Alcock & Anderson, 1894) View in CoL
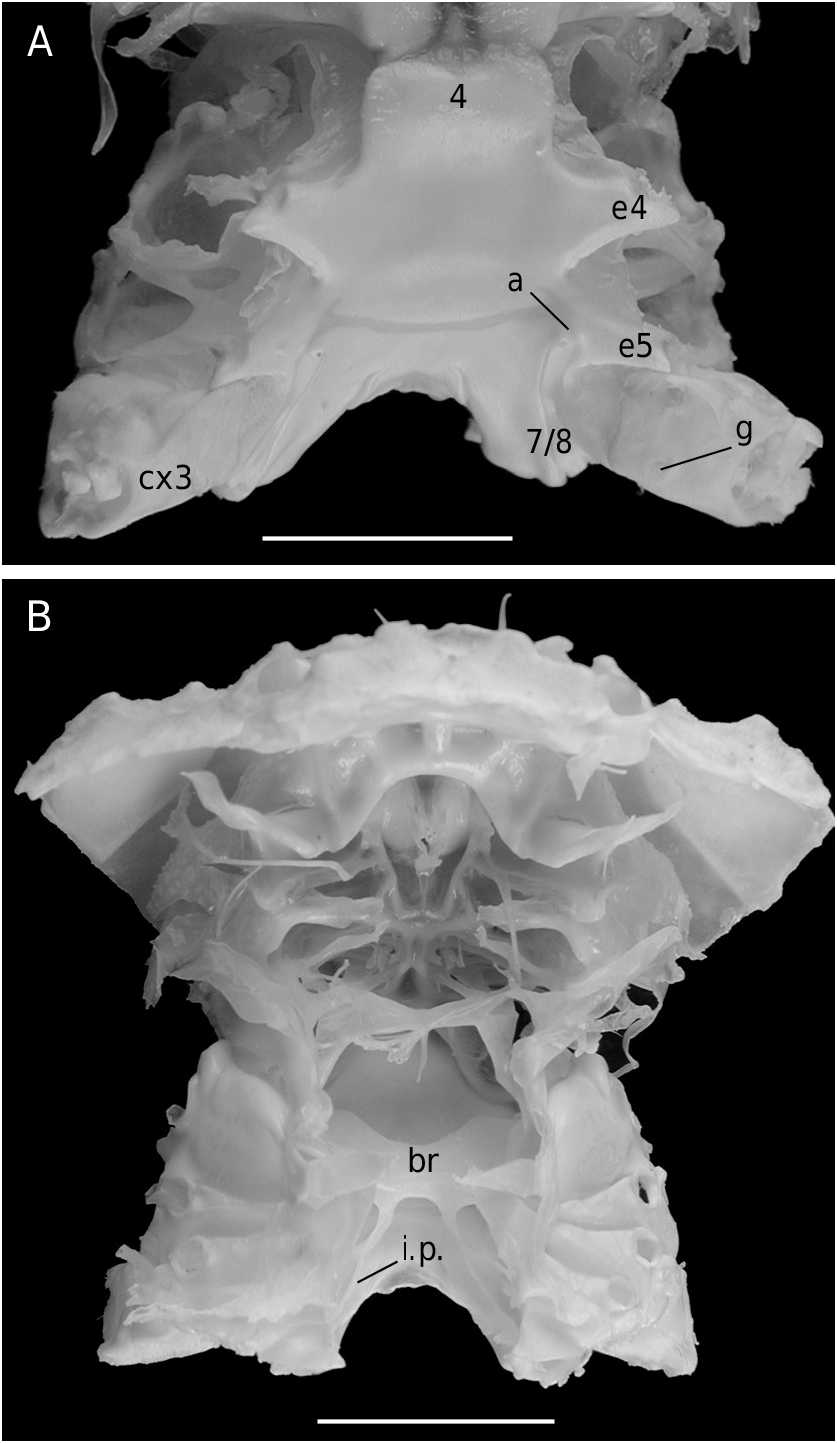
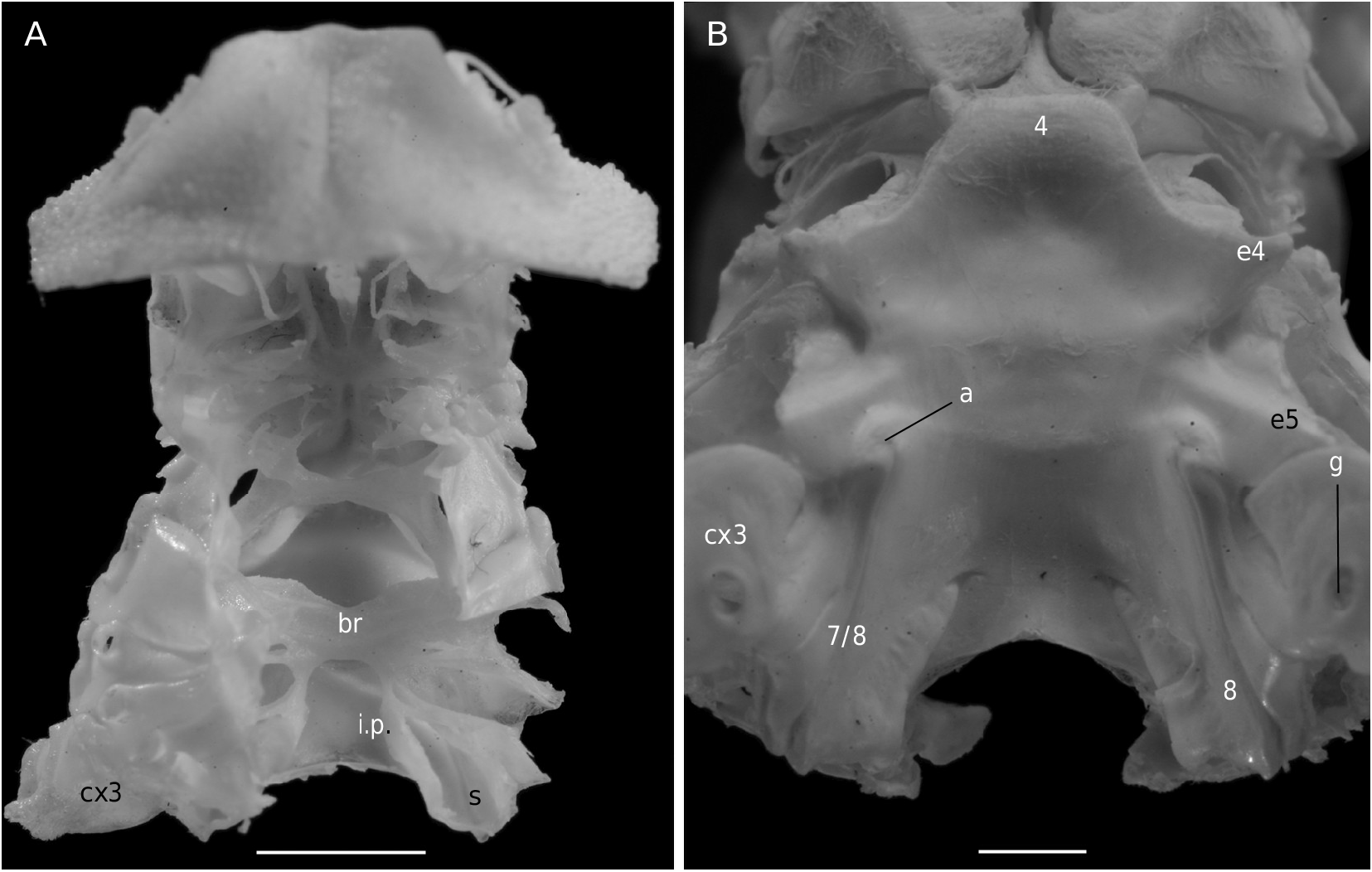
Ovigerous female 36.4 × 36.2 mm, Philippines, MUSORSTOM 3, CP 143 (MNHN-B 22543).
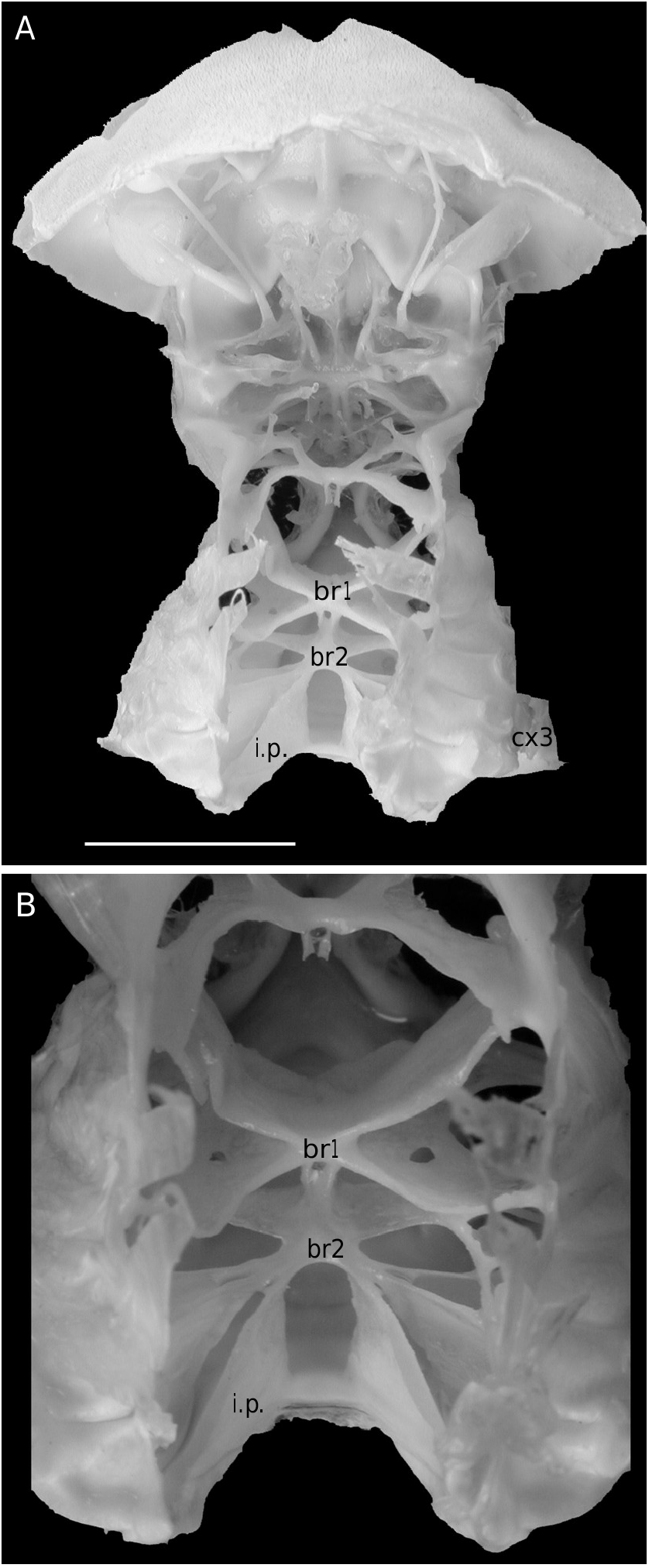
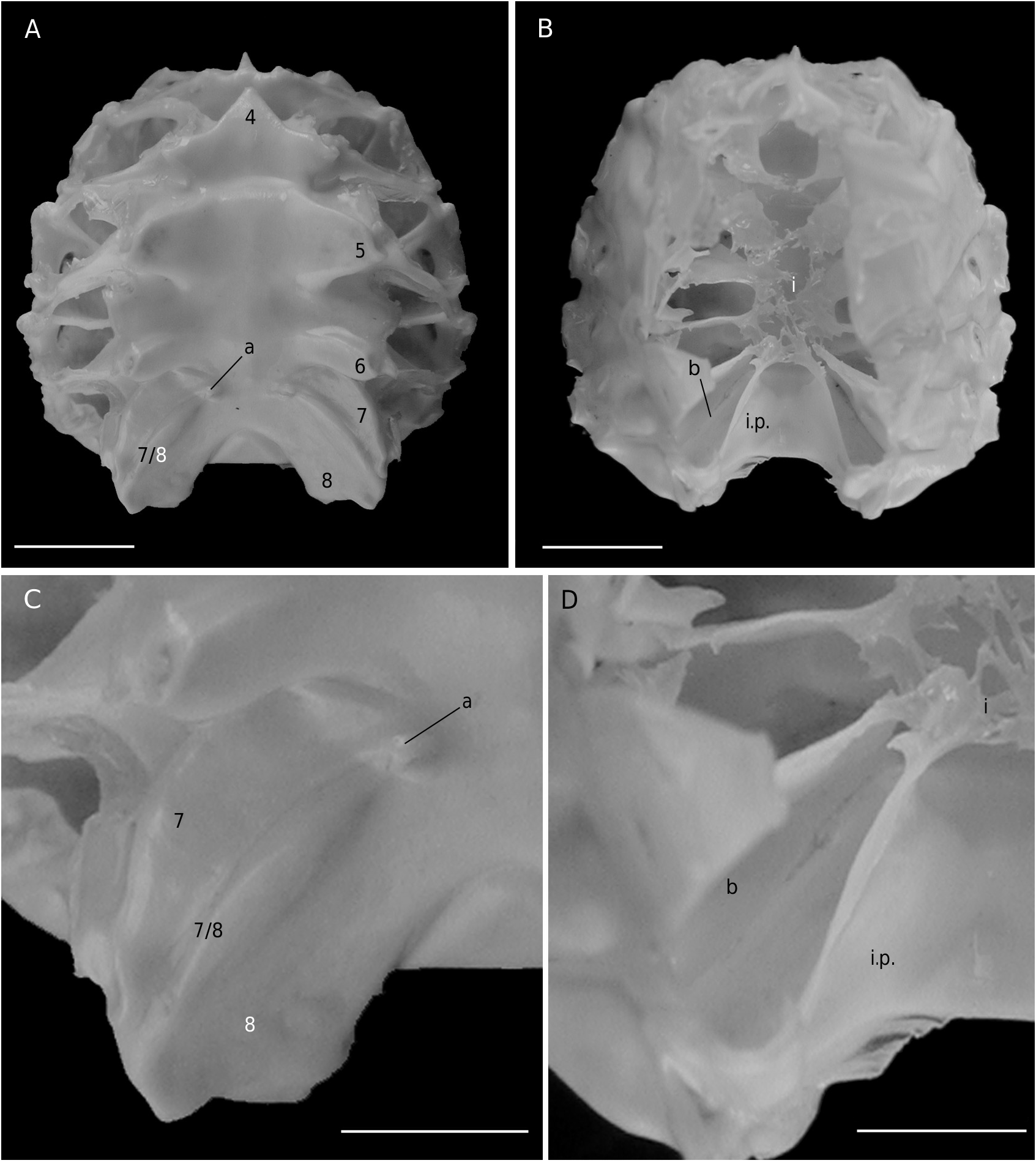
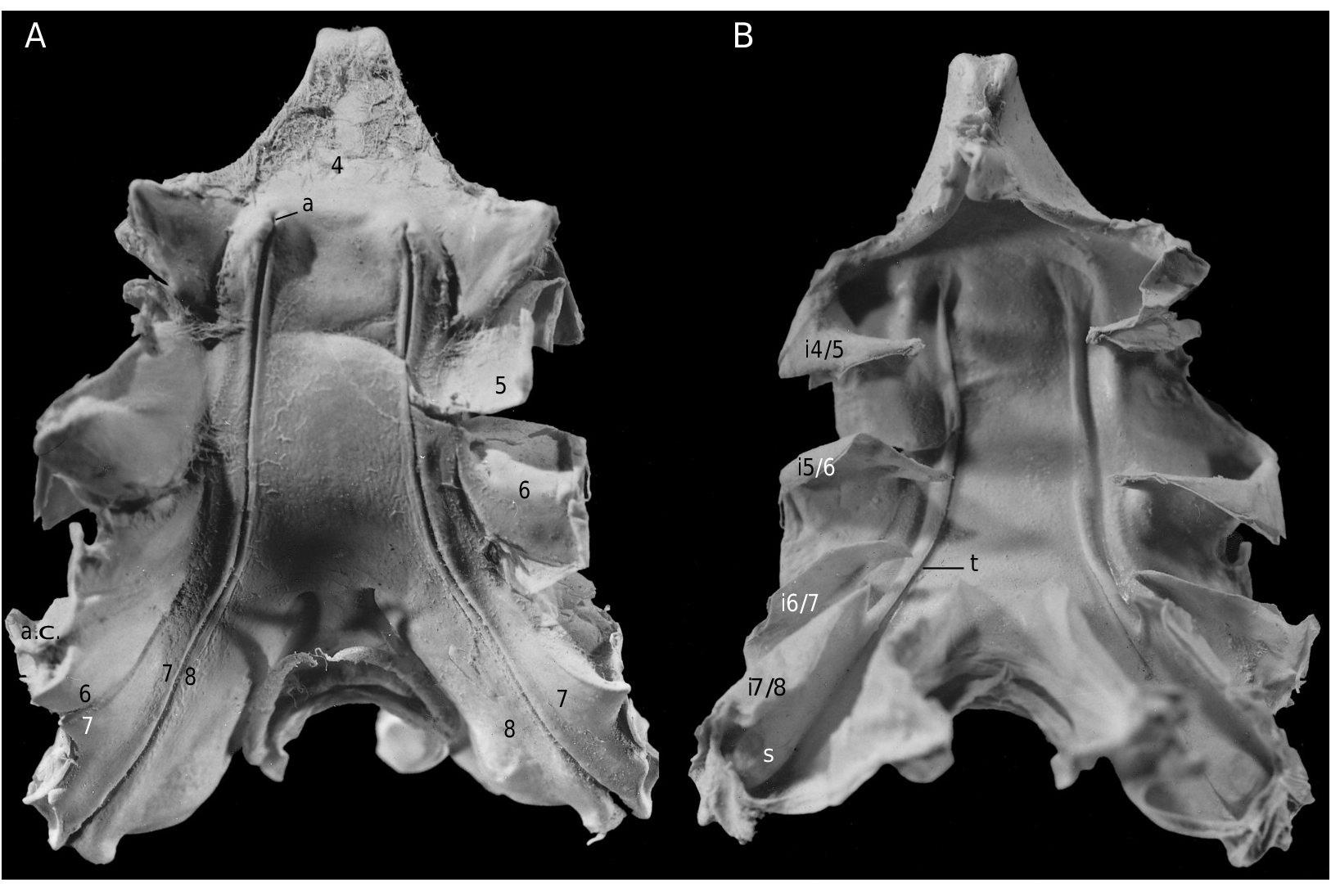
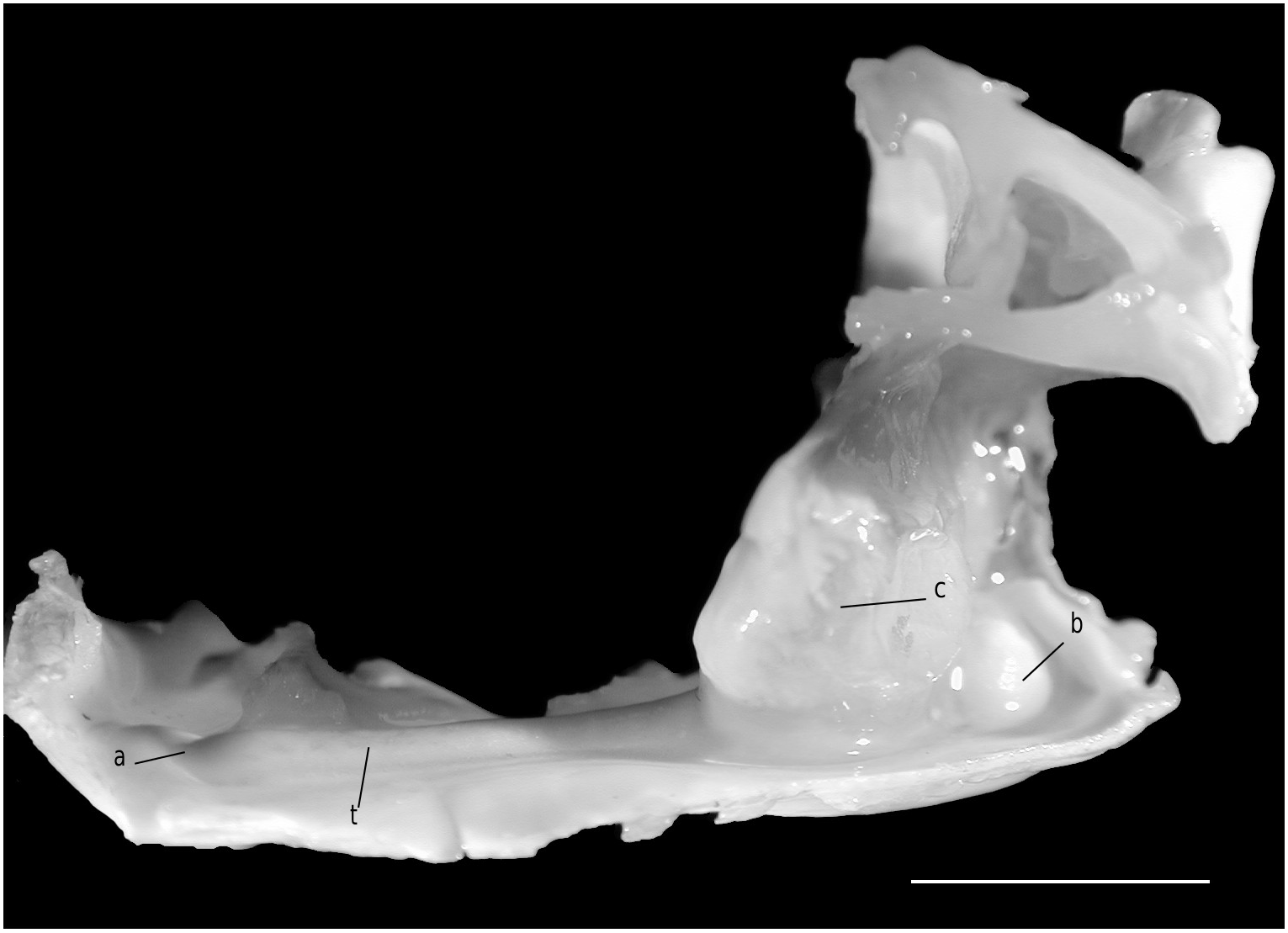
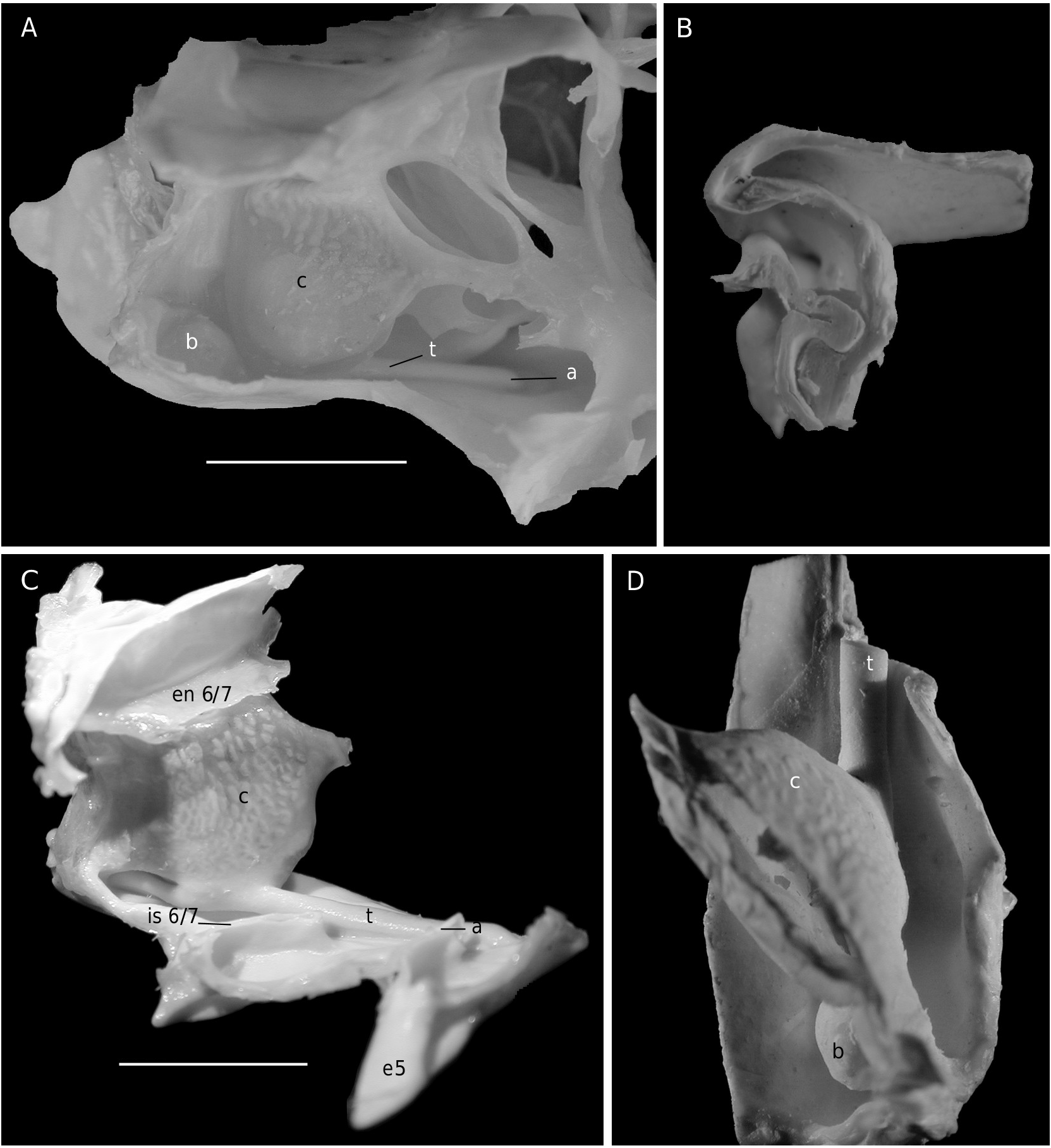
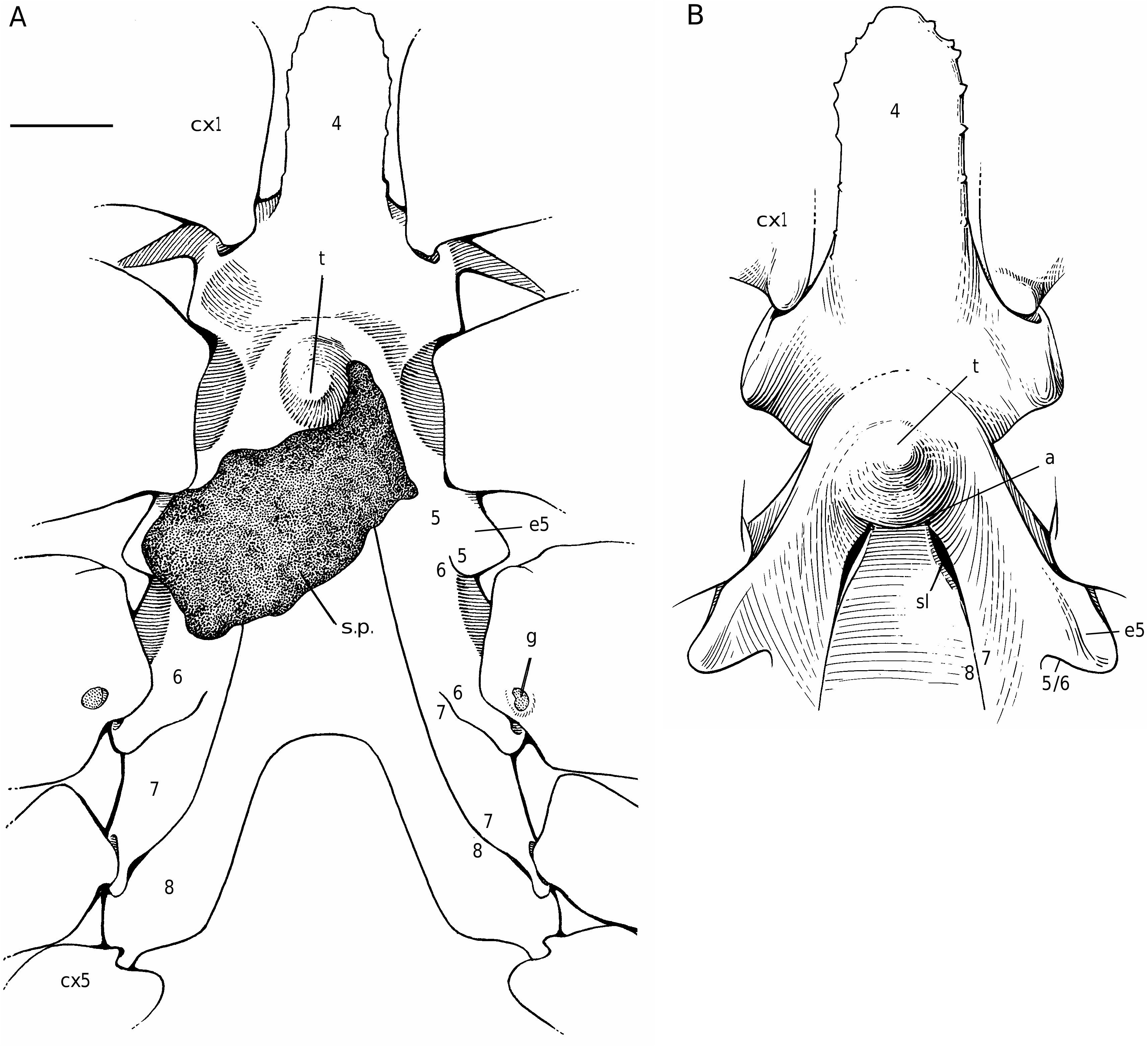
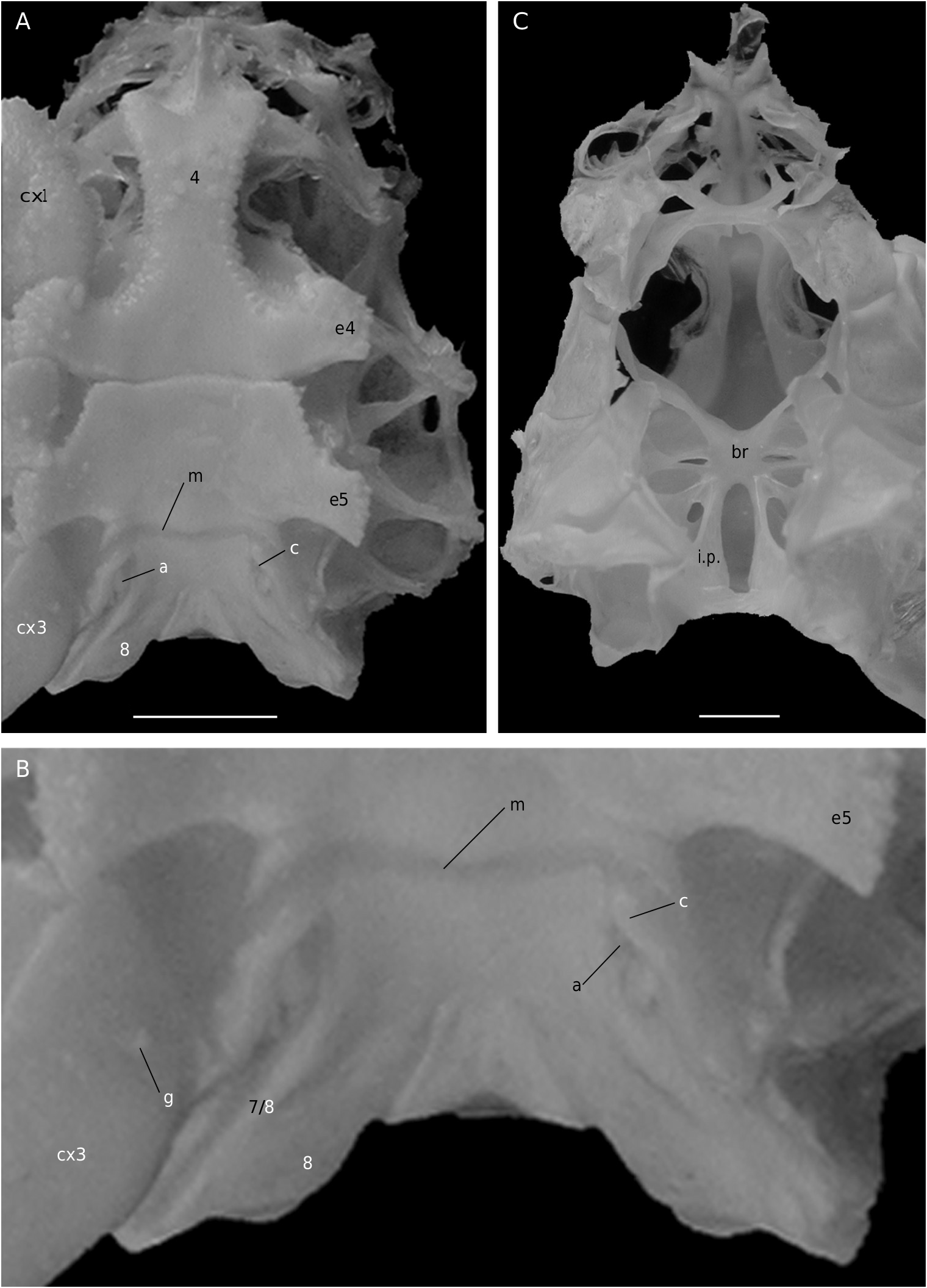
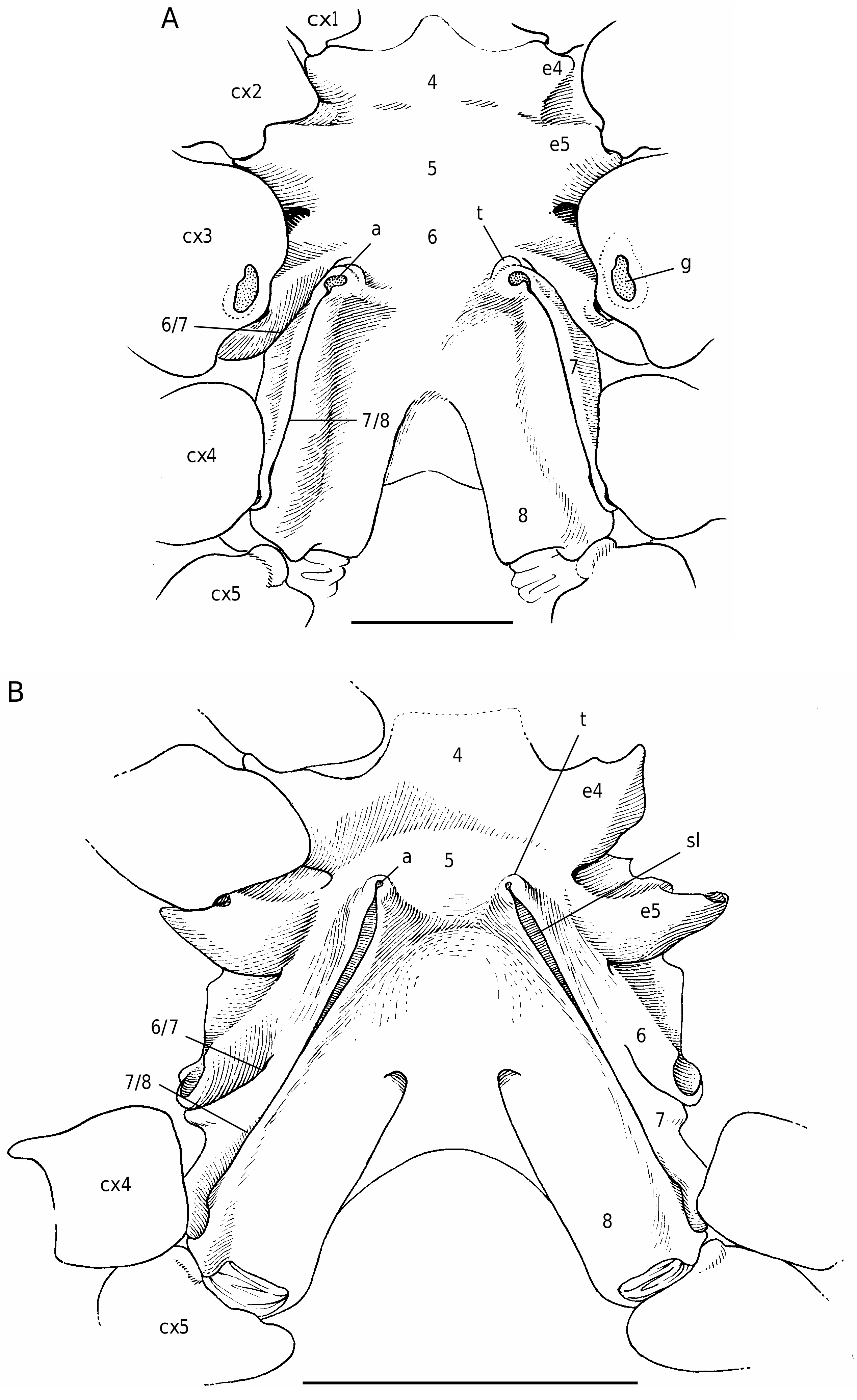
The axial skeleton, with two transverse bridges similarly to that of S. lamellata , was not completely dissected, but it was possible to partially examine the endosternite 7/8. Spermathecal apertures are concealed. Despite that suture 7/8 is slightly longer than in S. lamellata , a tube does not exist, however.
CONCLUSIONS FOR THE DROMIACEA
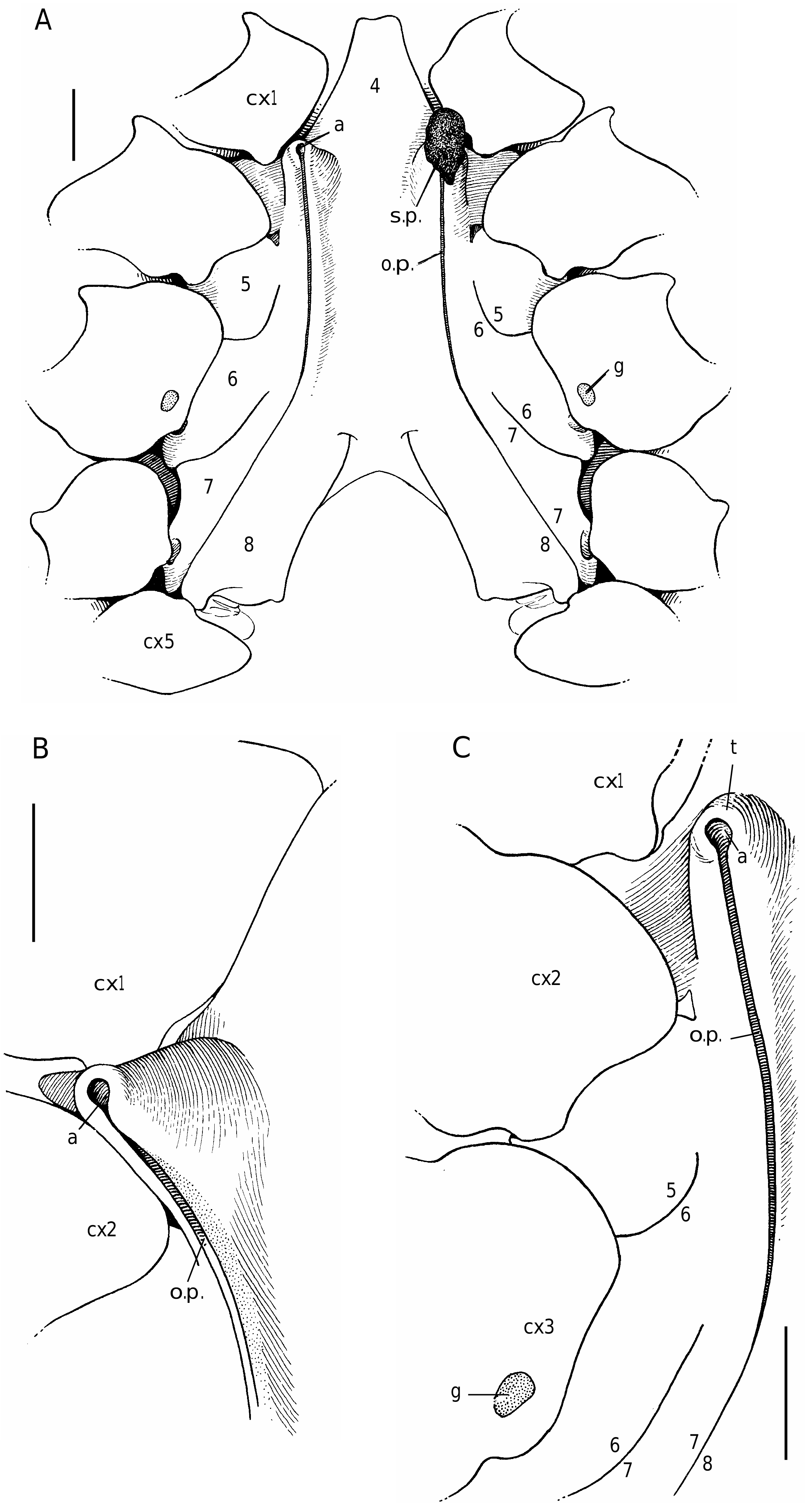
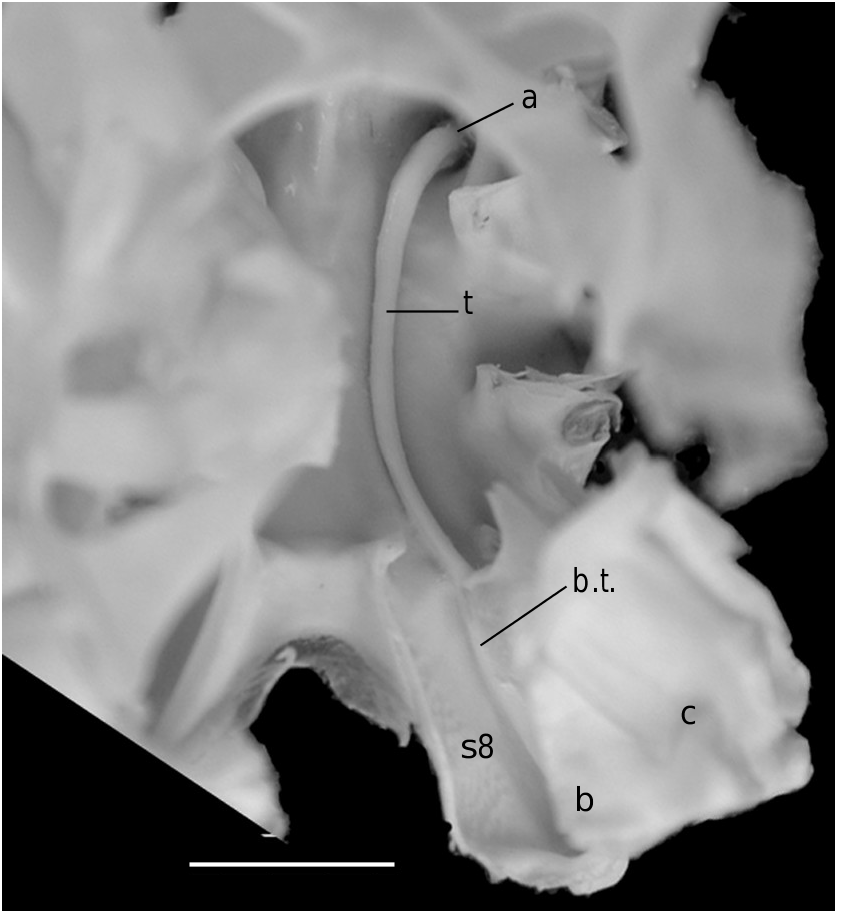
In all the Dromiacea that were examined, the spermatheca consisted of a bulb, which is formed by infoldings of the phragma 7/8, and a sort of pocket, herein called “chamber”, which corresponds to delamination of the two laminae of the endosternite 7/8, one belonging to the segment 7, the other to the segment 8. Another structure may be present along a part of the suture 7/8: an internal, closed tube, completely calcified, hollow and, in very most cases, perforated at its extremity, namely the spermathecal aperture. This tube is the forward prolongation of the chamber and continues well beyond it, sometimes along a great length of the suture 7/8 (in [ Dromia ] dormia, the tube is as long as the suture 7/8 of about 50 mm length). The tube, which is “doubtless formed by infoldings of sternites 7 and 8 along their sutural margins” ( Gordon 1950: 248), is the result of the fusion, on a short space, of the two laminae of the phragma 7/8. Its lumen forms a channel leading to the spermathecal aperture, which is generally terminal. Thus, the spermathecal tube corresponds to an entirely modified and elaborate structure: it is the result of a specialization. It is found only in the females. In dromiine males, the suture 7/8 is short, and the endosternite 7/8 is only a thin double septum, consisting of two coalescent sheets, not modified and without any prolongation. Gordon (1950: 245) observed that, in a small female Dromia “ already the anterior third of the suture 7/8 is wider and more conspicuous than it ever is in the male”. For Hartnoll (1979: 82) “the dromiid structure has been elaborated by the development of spermathecal canals along the line of suture 7/8 to link the openings with the spermathecae”.
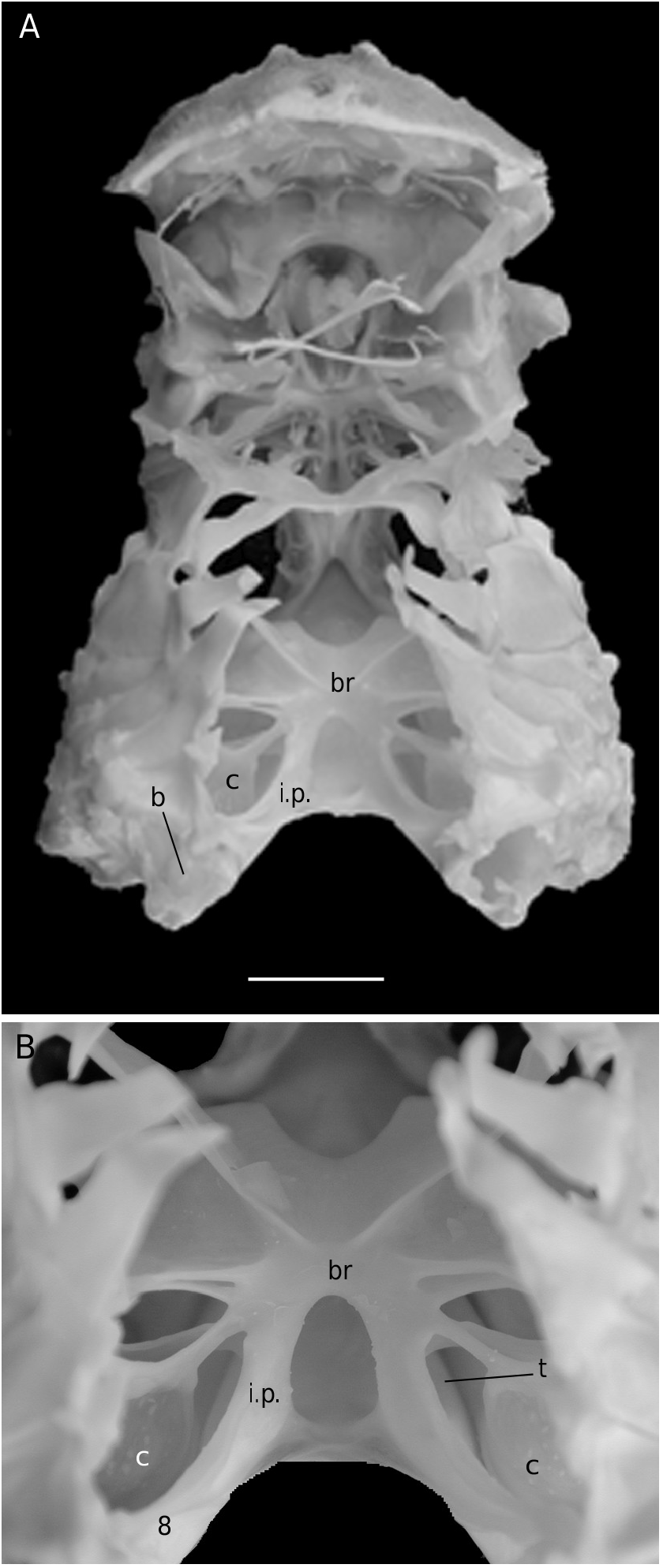
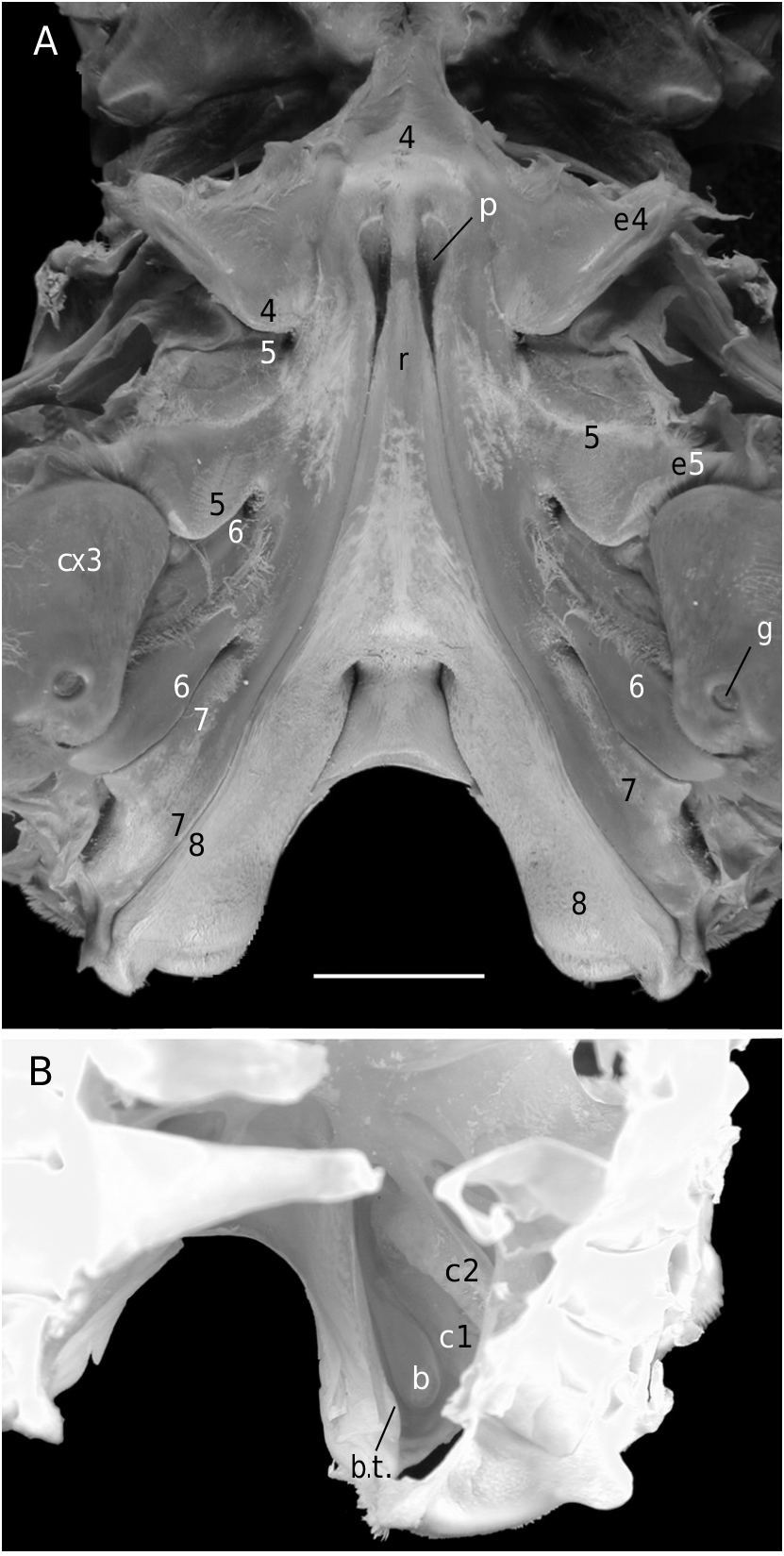
Considering the material available in the MNHN collections that was used to prepare the skeletons, the Dromiacea exhibit two main patterns concerning the spermathecal tube: 1) the tube is absent and the chamber communicates directly with the exterior through a spermathecal aperture: Dynomenidae ( Fig. 4 View FIG ) and Sphaerodromiinae ( Fig. 17 View FIG ); or 2) a more or less long tube is present and leads to an external aperture: Homolodromiidae ( Fig. 3 View FIG ), Dromiinae ( Figs 1 View FIG ; 2 View FIG ; 6-11 View FIG View FIG View FIG View FIG View FIG View FIG ; 13-15 View FIG View FIG View FIG ), Hypoconchinae ( Fig. 16 View FIG ), and the enigmatic genus Frodromia ( Fig. 12 View FIG ).
The visible but short tube is present even in the pattern with short sutures 7/8 ( Homolodromiidae ). Gordon’s (1950: 250, fig. 1) hypothesis about Dicranodromia baffini , a suture 7/8 of the dromiid type and “in all probability” a very short “channel leading to the spermathecal pit”, is thus verified.
The Homolodromiidae , which shows a unique combination of plesiomorphic morphological characters (pediform mxp3, abdominal pleurae, exopod present on G2, uropods showing as ventral lobes, elongated coxae of P5, skeletal connections by interfingering), exhibits the same spermathecal tube pattern as in the Dromiinae , in contrast to the Sphaerodromiinae (at least in the only ovigerous dissected female, Sphaerodromia lamellata ). If we consider the tube a more advanced condition, it corroborates the basal place of the Sphaerodromiinae within the Dromiidae , as previously pointed out ( Guinot & Tavares 2003). The presence of a (short but well visible) tube in the Hypoconchinae supports the placement of this subfamily in the Dromiidae . The specialized Hypoconcha is perhaps not as primitive as indicated until now.
The absence of a spermathecal tube in the Dynomenidae is of particular interest. The Dynomenidae shares several characters with the Dromiidae (dorsal uropods, skeletal junctions occuring by fusion), but it shows a peculiar mix of characters. Plesiomorphic are notably the presence of vestigial pleopods on the somites 3-5 as (generally) biramous vestiges, and elongated male coxae of P5. Apomorphic are a widened thoracic sternum and innovation in some genera ( Dynomene , Hirsutodynomene ) of a sternal differentiation on the thoracic somite 5 (corresponding to P2) for the holding (although not efficient, see Guinot & Bouchard 1998) of the abdomen.
To examine phylogenetic relationships between Homolodromiidae , Dromiidae and the Dynomenidae , spermatological characters (exemplified by Homolodromia kai , Sphaerodromia lamellata , and Metadynomene tanensis , respectively) were used. They strongly endorse monophyly of the Dromioidea , but without a clear recognition of the three families (Guinot et al. 1998).
In addition to features of the carapace during the course of this investigation, the Homolodromiidae , Dynomenidae and the Sphaerodromiinae share the following features, considered plesiomorphic: 1) male pleopodal formula complete or nearly complete (vestigial pleopods on abdominal somites 3-5, biramous in the Sphaerodromiinae and most Dynomenidae , uniramous in the Homolodromiidae ) (the Hypoconchinae maintains various vestiges of these pleopods, while most Dromiinae lack male Pl3-Pl5; see Guinot & Tavares 2003: table 1); 2) exopod present on G2 (in the Sphaerodromiinae , present only in the genus Sphaerodromia , probably absent in Eodromia ); G2 exopod absent in the Dromiinae and Hypoconchinae ; and 3) a totally modified male P5 coxa, which assumes the shape of a hard and elongated extension, resulting in a dimorphic male P5 coxa (the Dromiinae and the Hypoconchinae share a never dimorphic P5 coxa, bearing in males a long movable, “articulated”, sclerotized tube, the penial tube; see Guinot & Tavares 2003: 115, fig. 28).
The enigmatic genus Frodromia combines dromiine and sphaerodromiine characters, and its status remains uncertain ( Guinot & Tavares 2003: 110). Its sphaerodromiine features include short female sternal sutures 7/8 and spermathecal apertures lying close to the female gonopores on the P3 coxae. The present study permits to add two dromiine features to Frodromia : the presence of a spermathecal tube and skeleton with only one transverse bridge. The most important features which still conflict with the inclusion of Frodromia in the Dromiinae are the elongated male P5 coxa and the absence of a mobile penial tube.
All the Dromiacea, either basal ( Homolodromiidae , Dynomenidae , Spharodromiinae) or more advanced ( Dromiinae , Hypoconchinae ), share a similar spermathecal aperture, that is generally a small orifice. For the moment, only five cases differ from the typical condition.
1) In the Homolodromiidae , only one species, Dicranodromia spinulata Guinot, 1995 , shows suture 7/8 occupied by a long subterminal slit, while other homolodromiids have rounded or ovate terminal apertures.
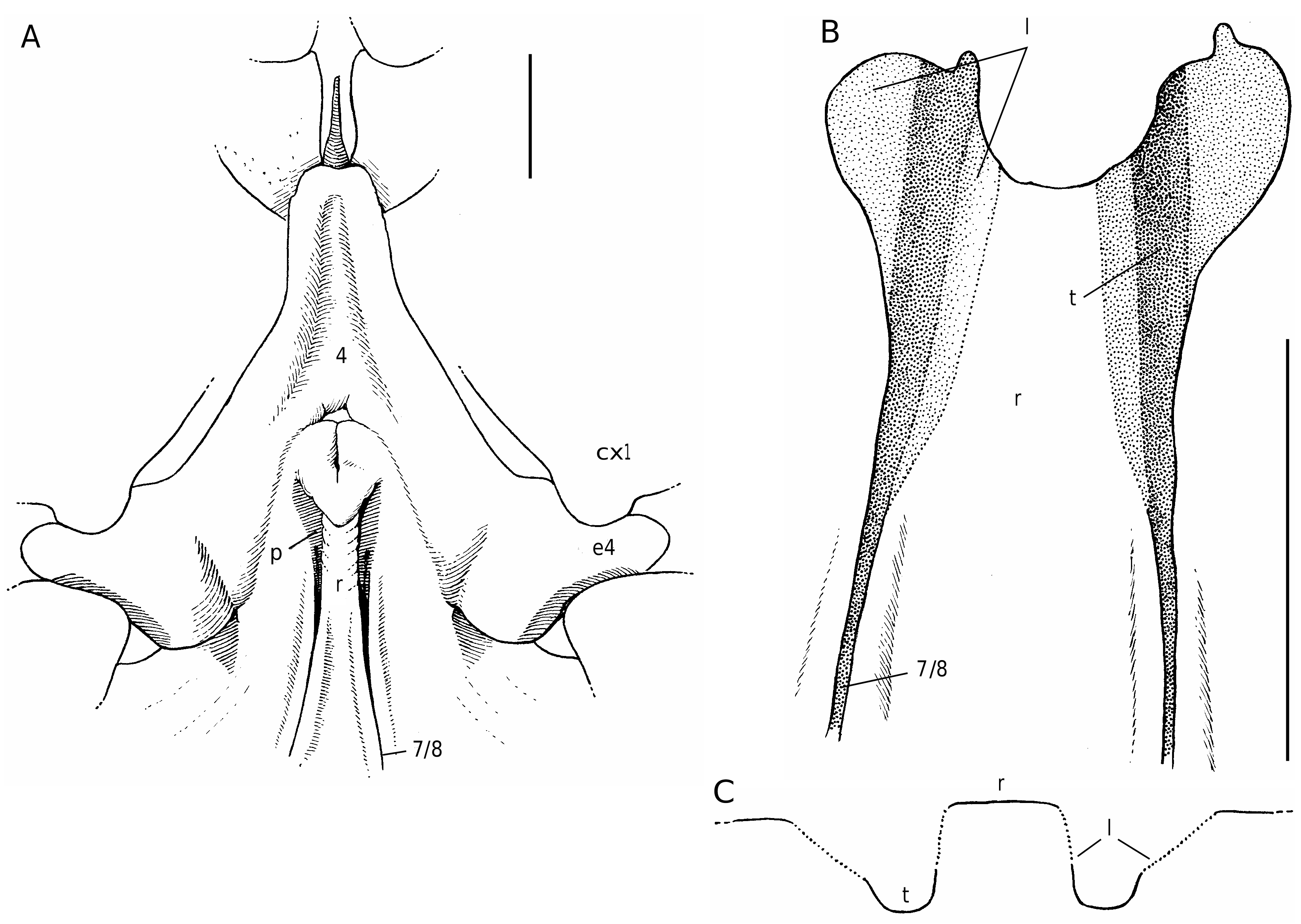
2) The dromiine genus Sternodromia ( Fig. 15 View FIG ) shows very tiny terminal apertures but they are replaced (or complemented) by subterminal slits, long ( S. spinirostris ) or shorter ( S. monodi ), the bottom of the tube being partly exposed and opened along its subterminal part so that the underlying spermathecal channel is connected with the exterior (Guinot unpubl. data).
3) The dromiine species [ Dromia ] dormia (a species that belongs to a different genus [Guinot unpubl. data]) is devoid of the usual circular terminal spermathecal aperture ( Figs 8A View FIG ; 9 View FIG ). Each suture 7/8 opens in a partly decalcified and unperforated pit; at its extremity, the thick spermathecal tube is no longer closed, and the bottom of the phragma 7/8 itself is opened and becomes exposed at the sternal surface to form an ovate pit, which is the spermathecal aperture.
4) In Lauridromia the two sheets of the phragma 7/8 are not completely joined subdistally, but it does not prevent the presence of a terminal aperture; sperm may penetrate in the space between the two separated sheets ( Fig. 13 View FIG ).
5) I n t h e g e n u s C r y p t o d r o m i a S t i m p s o n, 1858 sensu lato, the two phragmae of the endosternite 7/8 do not join, being sometimes significantly opened in their half distal part. In C. tuberculata Stimpson, 1858 ( Fig. 27B View FIG ), the margins of the suture 7/8 are distinctly raised, there is a wide, long slit, and the rounded aperture is located on the summit of a strong tubercle.
In the Dromiacea, due to the shortness or the lengthening of sutures 7/8, the spermathecal apertures are situated either just below or slightly behind the female gonopores on the P3 coxae ( Homolodromiidae pro parte, Dynomenidae ) or approximately opposite the P3 coxae ( Sphaerodromiinae , Stebbingdromia and Frodromia ). Or they extend slightly beyond and are not very far from the female gonopores ( Homolodromiidae pro parte, Hypoconchinae ) or extend far forward in leaving a great gap between the spermathecal apertures and the female gonopores and also the storage chambers (most Dromiinae ).


No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.