
Odontomachus clarus Roger, 1861
|
publication ID |
https://doi.org/ 10.11646/zootaxa.3802.4.6 |
|
publication LSID |
lsid:zoobank.org:pub:4A83AAAE-6F0B-4173-A066-DBBCFBC3BDD |
|
DOI |
https://doi.org/10.5281/zenodo.6129410 |
|
persistent identifier |
https://treatment.plazi.org/id/C66787A5-5B27-FF83-9AE0-F9E6FEF0FBF0 |
|
treatment provided by |
Plazi |
|
scientific name |
Odontomachus clarus Roger, 1861 |
| status |
|
Odontomachus clarus Roger, 1861 View in CoL
( Figs. 2 View FIGURE 2 , 4 View FIGURE 4 , 5 View FIGURE 5 , 6 View FIGURE 6 , 9 View FIGURE 9 , 10 View FIGURE 10 , 14 View FIGURE 14 , 15 View FIGURE 15 , 24 View FIGURE 24 , 25 View FIGURE 25 , 26 View FIGURE 26 )
Odontomachus clarus Roger, 1861: 26 View in CoL (worker) Texas, USA; Patton, 1894: 618 (male); Wheeler, 1908: 407 (queen); Petralia & Vinson, 1980: 381 (larva). Subspecies of O. haematodus: Emery, 1911: 115 View in CoL ; Wheeler, 1914: 40; Smith, 1939: 129. Revived status as species: Taylor & Wilson, 1962: 142; Brown, 1976: 136.
Odontomachus texana Buckley, 1867: 355 . Junior synonym of O. insularis: Emery, 1895: 268 View in CoL ; of O. clarus: Wheeler, 1902: 26 View in CoL .
Odontomachus haematodus View in CoL subsp. coninodis Wheeler, 1915: 391 ; Brown, 1976: 103. Junior synonym of O. clarus: Brown, 1976: 103 View in CoL .
Odontomachus haemataoda subsp. clarionensis Wheeler, 1934: 141. Junior synonym of O. clarus: Taylor & Wilson, 1962: 142 View in CoL .
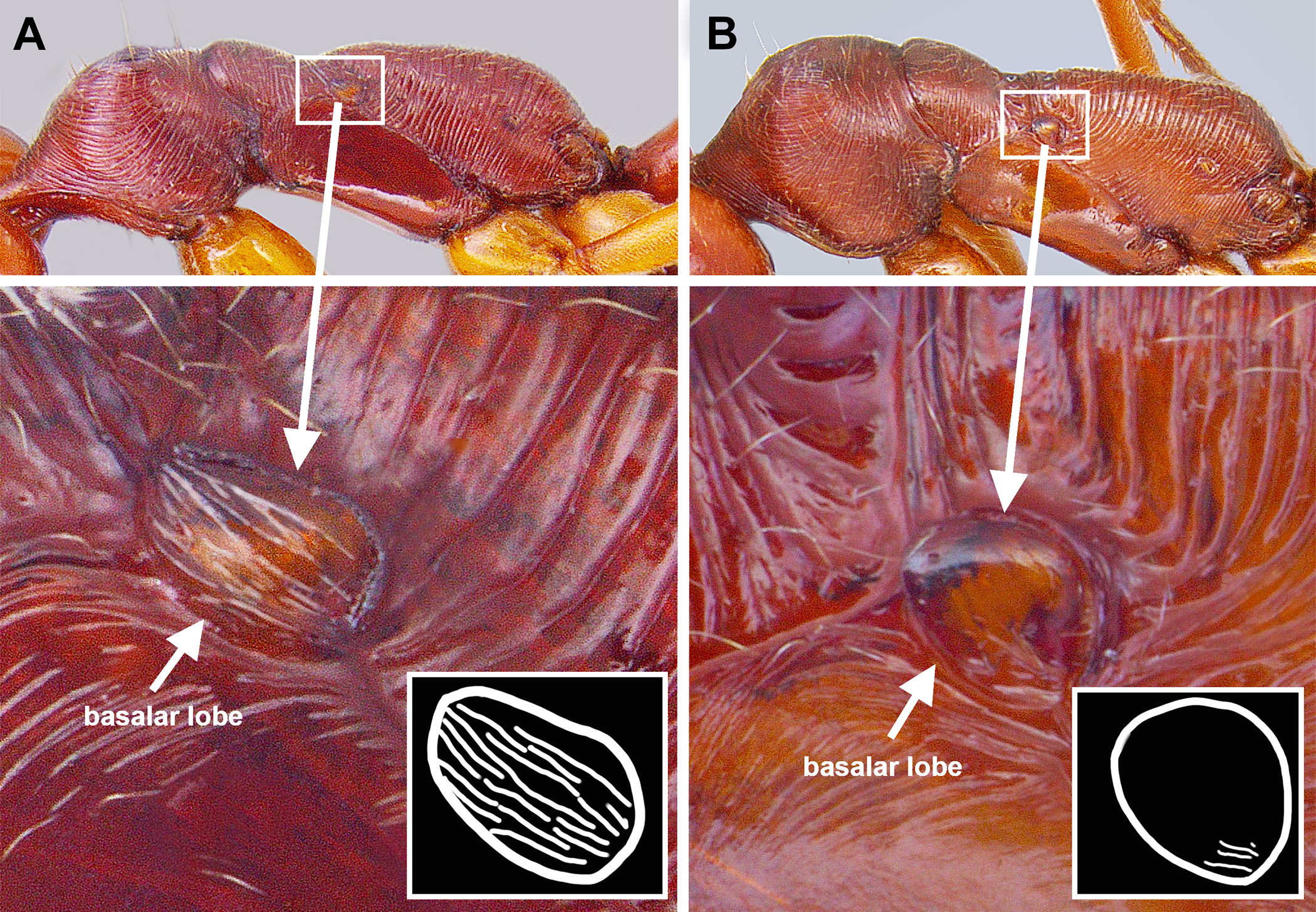
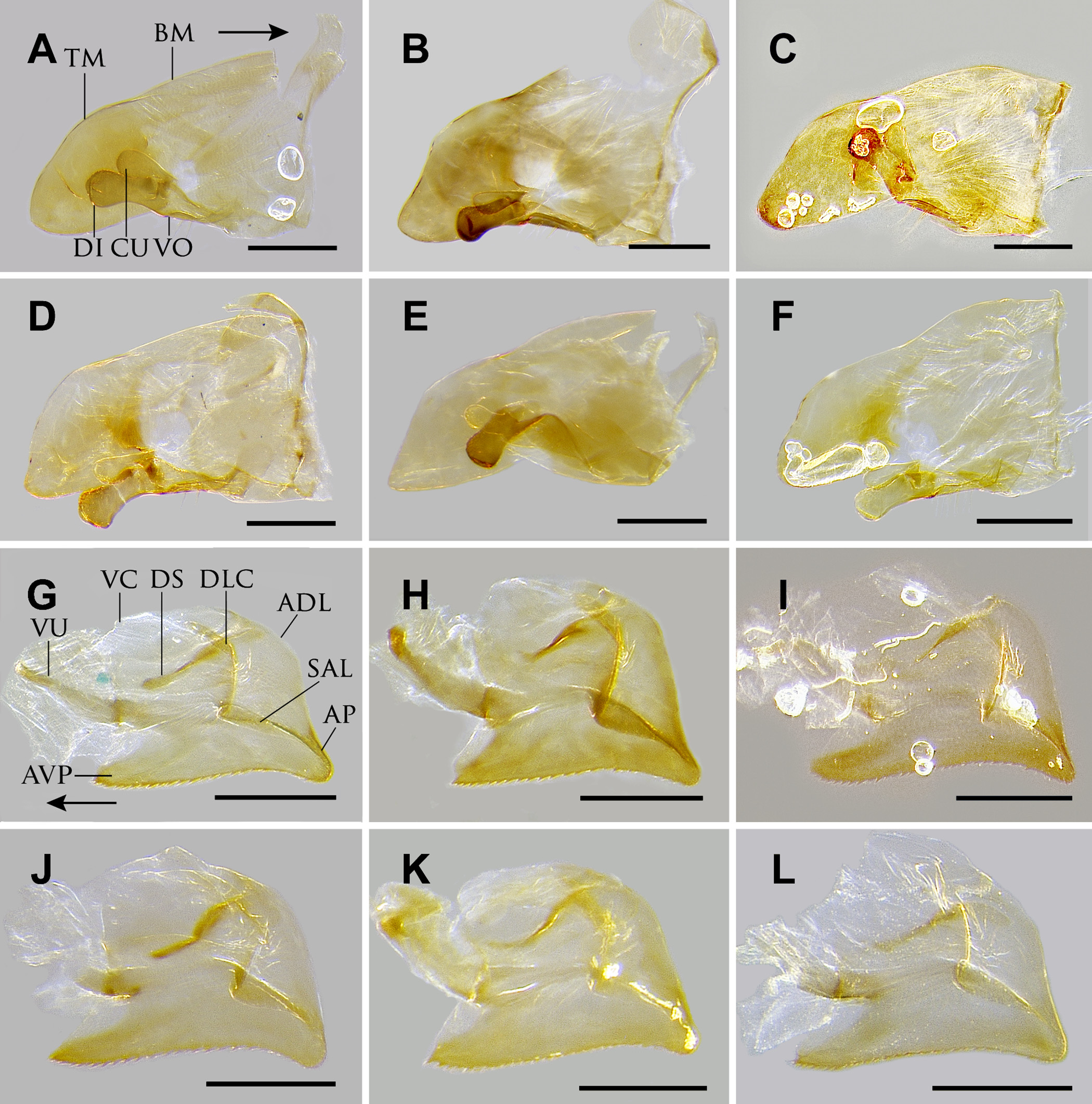
Diagnosis. Workers of O. clarus are most similar to O. relictus and O. desertorum , but are distinguished from O. relictus by their smooth basilar lobes (striate in O. relictus ), and from O. desertorum by the following: Size smaller (HL <2.2 mm, WL <2.8 mm); dorsomedian petiolar process nub-like or well-differentiated; propleurae almost entirely smooth and polished-looking; pronotal cervical lobe longer than broad; and antennal scapes relatively shorter (SL/HW = 0.99–1.07 vs. 1.11–1.15). Odontomachus clarus is further differentiated from O. relictus by geography: O. clarus only occurs west of the Mississippi river; whereas, O. relictus only occurs east of the Mississippi river. Males are uniquely identifiable among the Nearctic fauna by, at minimum, the following combination of characters: 1) dark brown coloration; 2) ocelli small, weakly bulging past posterior head margin; 3) mesoscutum longitudinally striate (striae may be posterolaterally divergent); 4) posterior margin of petiolar sternum lacking an angular process, but with paired setae on tubercles; and 5) propodeum finely striate. The ninth abdominal sternum of O. clarus is unique, having a short, broad apical portion of abdominal sternum IX ( Fig. 24 View FIGURE 24 B). The genitalia are otherwise most similar to those of O. desertorum , differing primarily in having a more strongly sclerotized digitus and anteroventral process shorter, sharply triangular, and not upturned.
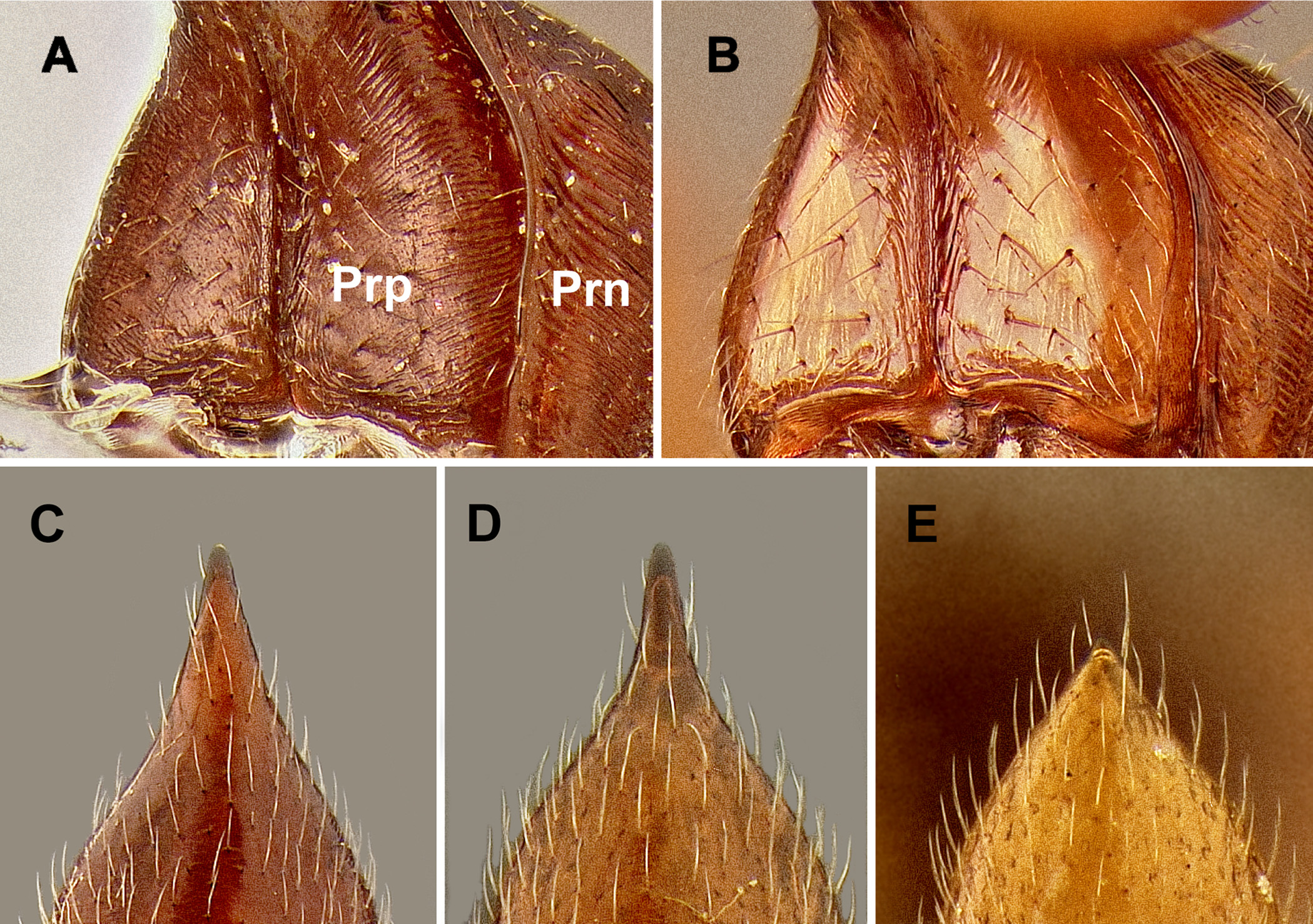
Description. Worker: HL 1.92–2.32, HW 1.57–1.90, SL 1.68–1.90, EL 0.31–0.36, ML 1.06–1.30, WL 2.43–2.83, PTH 0.81–1.10, PTL 0.42–0.48 (n=7). Entire body generally shiny except where obscured by dense pubescence; head, mesosoma, and petiole reddish-brown, legs yellowish-brown to orangish-brown, and gaster blackish. Head with fine, longitudinal striae covering much of the head in full-face view, striae beginning from frontal lobes and diverging toward posterior corners of head, fading at corners and sides; sides and underside of head lacking sculpture; with numerous, fine, appressed pubescence and scattered elongate, erect setae present dorsally. Pronotum with circular, slightly concentric striae that become transverse near rear margin; appressed pubescence abundant; 5–6 elongate, erect setae present. Mesonotum and propodeum with deep transverse striae; propleuron, mesopleuron, and basalar lobe lacking sculpture; abundant pubescence present dorsally. Metasternum lacking paired elongate, spiniform processes between hind coxae. Petiole widest at base, either gradually tapering apically to a short spiniform spine or stoutly conical (see Figs. 6 View FIGURE 6 D, E and 14F); mostly lacking striae with only faint striae present near base; subpetiolar process rounded triangular; appressed pubescence present anteriorly and laterally, but mostly absent posteriorly. Gaster mostly shiny beneath pubescence, lacking striae or other strong sculpture, but with fine coriaceous sculpture (seen at high magnification); slightly coarse, appressed pubescence somewhat sparse, hairs separated by at least 1/2 of their length; scattered erect, elongate setae present.
Male: HL 0.91–0.97, HW 1.18–1.30, SL 0.17–0.25, EL 0.62–0.64, EW 0.35–0.37, OL 0.12–0.14, OES 0.25–0.26, PTH 0.66–0.73, WL 2.39–2.65, PTL 0.40–0.59, FWL 4.56–4.97 (n=3). Body generally shiny except where obscured by dense pubescence; head and mesosoma brownish-black, petiole and gaster dark reddish-brown to brownish-black, legs and antennae reddish-brown. Head and body with abundant semi erect, whitish pubescence. Eyes extremely large, maximum diameter of each eye at least 70% of the length of the head in fullface view. Ocelli small, the length of each ocellus about half the distance between lateral ocellus and eye margin; in full-face view, lateral ocelli do not noticeably protrude beyond posterior border of head. Mesosoma: pronotum lacking sculpture; mesoscutum with fine transversely arcuate striae anteriorly, striae becoming longitudinal posteriorly; mesoscutellum raised and convex, with longitudinal striae; propodeum and mesopleuron with weak to slightly strong longitudinal striae. Petiole bluntly rounded apically, with apically acute to apically rounded triangular subpetiolar process present anteriorly; densely pubescent anteriorly and laterally, with reduced pubescence posteriorly; petiolar sternum lacking angular process near posterior margin, but with paired setae on small tubercles. Abdominal sternum IX disc subrectangular, breadth almost twice length; posterior lobe length less than twice maximum width, basal half barely narrowed. Telomeral apex narrowly rounded; telomere length distinctly greater than height; valviceps ventral apex strongly produced and very narrow; apical margin of valviceps linear until abruptly curving posteriorly at apicoventral process; anteroventral process of valviceps sharply triangular, and not upturned; vertical portion of dorsolateral carina and lateral margin of subapical lamina curving into one another; apicodorsal lobe of valviceps narrow and dorsally elongate; subapical lamina broad.
Queen: No specimens measured, but similar to workers in color and general appearance except slightly larger, with mesosoma developed for wings.
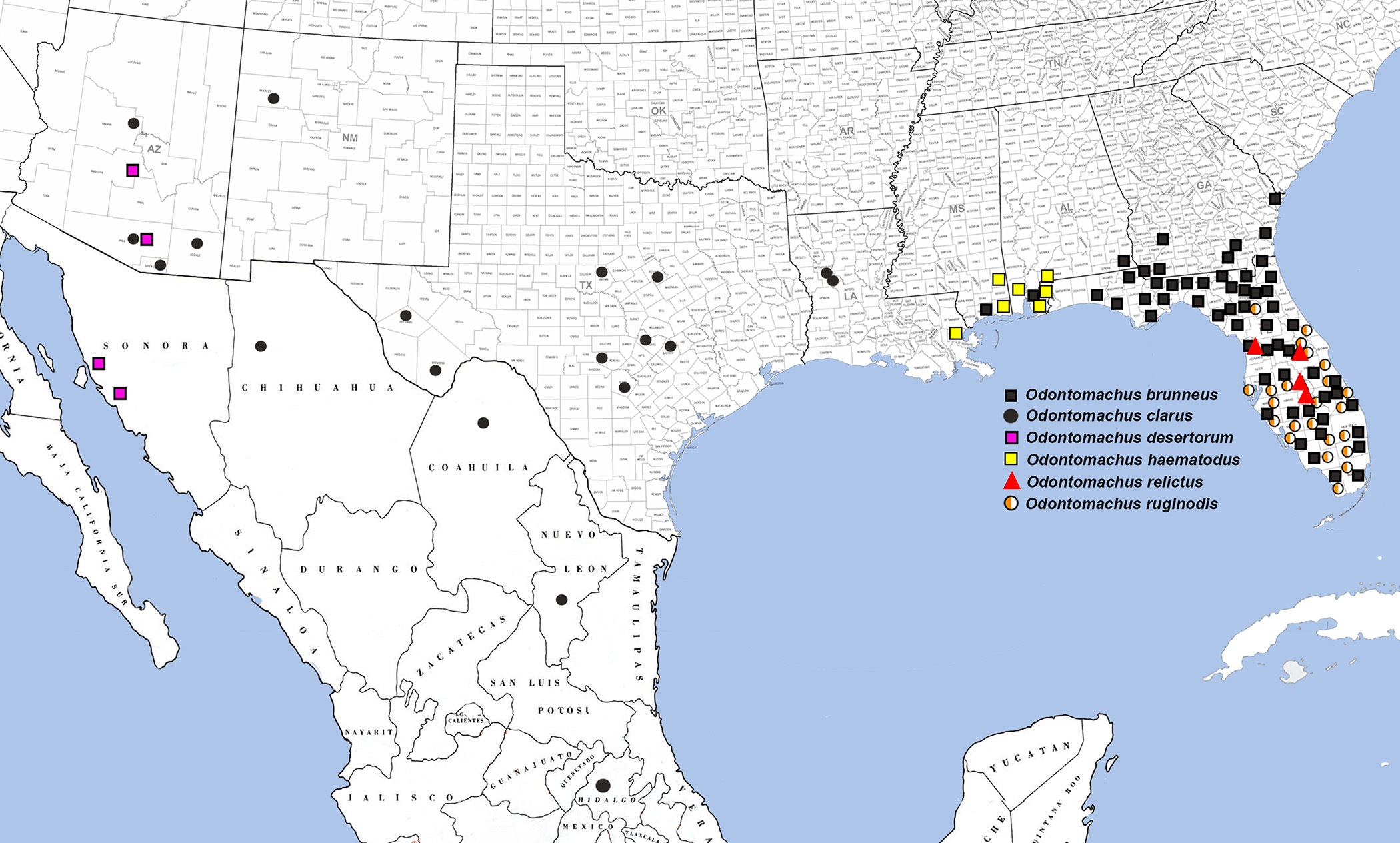
Distribution ( Fig. 26 View FIGURE 26 ). Mexico. Chihuahua, Coahuila, Hidalgo, Nuevo Leon, and Clarión Island. USA. Arizona: Cochise, Pima, Santa Cruz, and Yavapai Counties. New Mexico. McKinley County. Texas: Bastrop, Bexar, Bosque, Brewster, Brown, Denton, Jeff Davis, and Travis Counties. Louisiana: Natchitoches Parish. Distributional information from AntWeb (2013), Anonymous (2013), Brown (1976), Adams et al. (2010), Deyrup and Cover (2004), and examination of specimens from ABS, JTLC, LACM, LSAM, MEM, UCDC, UGA, and UTEP.
Discussion. Odontomachus clarus has had four specific synonyms until the present paper. While the Buckley name may be safely ignored as superfluous and O. desertorum is here recognized as a valid species, the speciesstatus of the other two names stands to be reassessed. Odontomachus clarus is variable across its range, with a broad longitudinal gradient from Texas and Louisiana South to Hidalgo, Mexico, and a very narrow shift from Texas into the mountains of Arizona. The US-Mexico cline is manifested in a North-to-South trend of increasing intensity of gastral sculpturation, erection of gastral pubescence, and anteroposterior compression of the petiolar node. The Mexican High Plateau population is otherwise identical to those of the Great and South Central Plains. The montane Arizonan populations, located in the Chiricahua, Huachua, and Santa Rita Mountains, as well as the Colorado Plateau Transition Zone, have previously been provided with the subspecific name O. clarus coninodis ( Wheeler 1915) due to their nub-like dorsomedian petiolar process (see Fig. 6 View FIGURE 6 E). Upon careful examination of material across the range of O. clarus it is clear that this is the only distinguishing character for these populations, and that the conical dorsomedian petiolar process merely represents a foreshortening of the process as the rest of the petiole is otherwise unmodified. Moreover, males attributable to “ coninodis are inseparable from those of O. clarus except for reduced size. These observations strongly suggest that the name O. clarus coninodis should remain in synonymy with O. clarus . It would still be valuable to conduct a sampling transect from Texas through the southeastern Arizona mountains to detect intergradation of petiolar node form as Brown (1976) suggested.
Finally, the status of the name O. clarus clarionensis remains to be absolutely determined. The name was provided by Wheeler (1934) for an insular population on Clarión Island (about 700 km WSW from the tip of Baja) of what he considered to be O. haematodus . Wheeler erected the subspecies based on the larger size, broader head, and smoother posterolateral head corners than compared to O. haematoda (an artificial assembly not recognized as such at the time). Taylor and Wilson (1962) synonymized the subspecies with O. clarus without discussion, and the population has been noted as “typical for O. clarus by Brown (1976). Examination of one of the cotypes at the MCZ does not reveal any salient distinctions of the Clarión Island population from mainland O. clarus . The synonymy is thus tentatively confirmed here; however, it would be valuable to sequence genetic material of the Clarión Island population of O. clarus to compare to material from across the species’ range.
The first report of Odontomachus clarus from Louisiana was based on specimens collected in Provencal in Natchitoches Parish ( Smith 1939). Unfortunately, we were unable to locate Smith’s specimens to verify this record. Subsequent authorities implicitly questioned this disjunct record: Brown (1976) omitted it entirely from his lengthy monograph, and this record was later omitted by Deyrup and Cover (2004). Smith's efforts were later vindicated, however, as specimens of O. clarus were collected nesting under large rocks in sandy soil in the Kisatchie National Forest in Natchitoches Parish just 20 km SW of where Smith reported this species ( Adams et al. 2010). Similarly, in Texas, nesting sites are typically in dry sites. In Arizona, its nesting habits appear more flexible, as it has been collected under rocks and grass tussocks in dry to mesic habitats ( Deyrup and Cover 2004). These differences in nest site selection may yet reveal whether the “ coninodis populations represent a distinct species, although this seems unlikely. Regardless of the results of increased sampling and molecular work on the Arizona populations, the Louisiana populations of O. clarus would certainly retain that specific epithet as the type series was collected from Texas.







No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.
|
Kingdom |
|
|
Phylum |
|
|
Class |
|
|
Order |
|
|
Family |
|
|
Genus |
Odontomachus clarus Roger, 1861
| Macgown, Joe A., Boudinot, Brendon, Deyrup, Mark & Sorger, D. Magdalena 2014 |
Odontomachus haemataoda
| Taylor 1962: 142 |
| Wheeler 1934: 141 |
Odontomachus haematodus
| Brown 1976: 103 |
| Brown 1976: 103 |
| Wheeler 1915: 391 |
Odontomachus texana
| Wheeler 1902: 26 |
| Emery 1895: 268 |
Odontomachus clarus
| Petralia 1980: 381 |
| Brown 1976: 136 |
| Taylor 1962: 142 |
| Smith 1939: 129 |
| Wheeler 1914: 40 |
| Emery 1911: 115 |
| Wheeler 1908: 407 |
| Patton 1894: 618 |
| Roger 1861: 26 |