
Elysia asbecki, Wägele, Heike, Stemmer, Kristina, Burghardt, Ingo & Händeler, Katharina, 2010
|
publication ID |
https://doi.org/ 10.5281/zenodo.199203 |
|
DOI |
https://doi.org/10.5281/zenodo.6204676 |
|
persistent identifier |
https://treatment.plazi.org/id/C72BF37D-E823-8751-05AD-ECC5FEFFA8D7 |
|
treatment provided by |
Plazi |
|
scientific name |
Elysia asbecki |
| status |
sp. nov. |
Elysia asbecki View in CoL sp. nov.
Type material. All specimens were collected in the intertidal reef flat of South Island, Lizard Island, North Queensland, Australia. Animals have been discovered in trays after collecting algae and coral rubble from these reef flats. Type material is deposited at the Australian Museum Sydney. Holotype (AM C.464069): 11th July 2006 (length of preserved specimen 3 mm); one paratype partly dissected (AM C.464070, including SEM preparation of radula): 11th July 2006 (length of preserved specimen 2 mm). For further material see Table 5.
Date of collection Date and kind of preservation Type of investigation
11.07.2006 12.07.0 6 EtOH Holotype AM C.464069
11.07.2006 12.07.0 6 EtOH Paratype AM C.464070
Gene analysis: CO1
Radula preparation (LM, SEM)
11.07.2006 12.07.0 6 EtOH Gene analyses (partial 28S, 16S and CO1 gene sequences)
(publ. in Händeler et al. 2009 as Elysia spec . 1) 11.07.2006 12.07.0 6 FSW Histology ( ZSM 20100676)
11.07.2006 Died after PAM measurement PAM (9 days; see Fig. 12)
13.09.2004 13.09.2004 FSW animal lost during radula preparation
Distribution. Up to now this species has been recorded from South Island, Lizard Island on top of the reef flat in the intertidal zone. A few specimens of this species have also been found by one of the authors (IB) in the Samoan Islands in 2005 (unpublished data). Gosliner et al. (2008) spotted this species in different places in the Indopacific ( Papua New Guinea, Indonesia, Philippines, Japan, Guam and the Hawai’ian Islands) and figured it under the designation Elysia sp. 16.
Etymology. This species is dedicated to Dr. Frank Asbeck (SolarWorld AG, Bonn) for his continuous sponsorship to the Zoologisches Forschungsmuseum Alexander Koenig, Bonn, Germany.
Description. External morphology and color of living specimens ( Fig.7 View FIGURE 7 ). Size up to 8 mm. Body elongate when crawling ( Fig. 7 View FIGURE 7 B) or more compact when resting ( Figs. 7 View FIGURE 7 A, C, D). Prominent parapodia not fused anteriorly; usually covered with tiny tubercles, which change in height within the same animal (compare Figs. 7 View FIGURE 7 B, C, D). Margin of parapodium forming rather irregular lobes. Rhinophores rolled and rather short. No distinct propodial tentacles at anterior foot present. Pericardial prominence oval with narrow posterior elongation ( Fig. 7 View FIGURE 7 A). Dorsal vessels indistinct.
Background color of animals appearing whitish, due to accumulation of white dots; large amounts of yellow to orange spots scattered over body, sometimes densely arranged in stripe-like patterns ( Fig. 7 View FIGURE 7 D). Tiny black spots evenly distributed on outer side of parapodia. Inner surface of parapodia and foot-sole translucent, with less numerous white spots, no black spots and with green digestive gland shining through. Head with similar coloration as body, in the neck region with a lighter spot. Eyes clearly visible behind rhinophores. Rhinophores translucent whitish with darker ring, followed by a yellow ring towards the tip. Two pink elongate patches on both sides of tail ( Fig. 7 View FIGURE 7 C), as well as at frontal base of parapodia at the conjunction with head ( Figs. 7 View FIGURE 7 B, C). Similar patches present at margin of parapodia, mainly in median part.
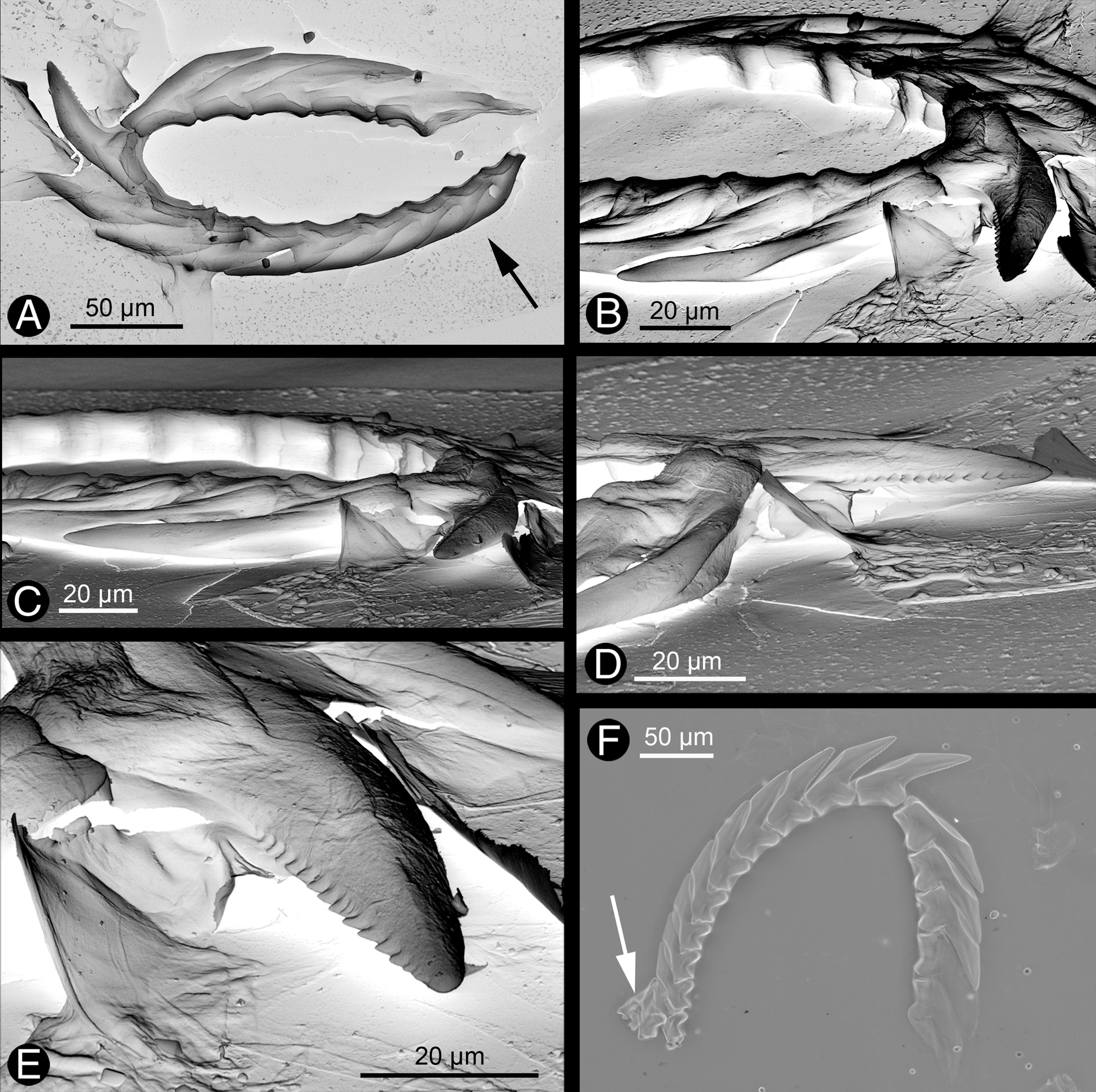
Description of anatomy and histology of preserved specimen ( Figs. 8–10 View FIGURE 8 View FIGURE 9 View FIGURE 10 ). Digestive tract. Oral tube in the beginning without any subepithelial glands, but with ciliated cells. More posteriorly, oral tube surrounded by subepithelial glands with acid mucopolysaccharides (oral glandular layer) ( Fig. 8 View FIGURE 8 A). Pharynx muscular, with a dorsal muscular pump. Radula with five teeth in descending limb and nine in ascending limb ( Figs. 9 View FIGURE 9 A, F). Four smaller teeth and two pre-radula teeth in ascus present ( Fig. 9 View FIGURE 9 F). Teeth with a denticulate cutting edge ( Figs. 9 View FIGURE 9 B, D, E). Salivary glands lobate, located postero-ventrally to pharynx, as well as lateral and ventral to oesophagus and stomach ( Fig. 8 View FIGURE 8 C, Fig. 10 View FIGURE 10 ). Gland composed of large secreting cells with granules staining bluish to dark violet and surrounding a tiny duct. Duct running anterior along pharynx, but entrance into pharynx not verified. Oesophagus starting from posterior part of pharynx and entering stomach a short distance behind on dorsal side. Epithelium of oesophagus highly folded and heavily ciliated ( Fig. 8 View FIGURE 8 B). Few violet stained glandular cells (acid mucopolysaccharides) interspersed. Posterior part of oesophagus surrounded by thick muscle layer, but not differentiated into distinct bulb.
Stomach large, lined by ciliated epithelium. Transition into right and left digestive gland facing each other on lateral sides of stomach ( Fig. 8 View FIGURE 8 B, Fig. 10 View FIGURE 10 ). Digestive gland ramifying heavily, with branches reaching into lateral parapodia as well as into foot area ( Fig. 8 View FIGURE 8 A). One tiny branch reaching half way into the rhinophores. Cells of these branches filled with chloroplasts. Intestine originating dorsally from stomach and opening to outside behind right rhinophore, between lateral foot side and parapodium. No typhlosole present in proximal part of intestine. Epithelium folded, with ciliated cells, but without any glandular cells. In some areas, chloroplasts present in lumen of intestine.
Genital system. The histologically investigated specimen was juvenile. Few gonad follicles present in parapodia, located dorsally in outstretched parapodia. Only spermatogonia in early stages recognizable. Female part of system not developed yet. A small penis present without cuticular structures ( Fig. 8 View FIGURE 8 E); epithelium of vas deferens inside penis formed by cuboidal cells with light bluish contents indicating secretory function.
Excretory and circulatory systems. Pericardial region in anterior third of body forming distinct hump lying above pharynx. Ventricle inside pericardium muscular. Kidney forming sac-like structure, starting within this hump and reaching into posterior part of body.
Sensory organs. Eye with homogeneously stained globular lens ( Fig. 8 View FIGURE 8 A); pigment cup of eye orientated to dorsolateral side. Statocysts large, lying between cerebral and pedal ganglion, containing one large otolith ( Fig. 8 View FIGURE 8 F).
Epithelia and glandular structures. Epidermis composed of flat cells ( Fig. 8 View FIGURE 8 B); few subepidermal glandular cells present only above pericardium and excretory system (dorsal hump) ( Fig. 8 View FIGURE 8 A). Foot characterized by a loose layer of subepidermal mucus glands similar to those of dorsal area and some regions of parapodia. In few areas of parapodia, agglomerations of specialized cells visible. Form and shape of these cells differ, in some with star-like contents, in others with a vacuole that is filled with particulate-stained contents ( Fig. 8 View FIGURE 8 D).
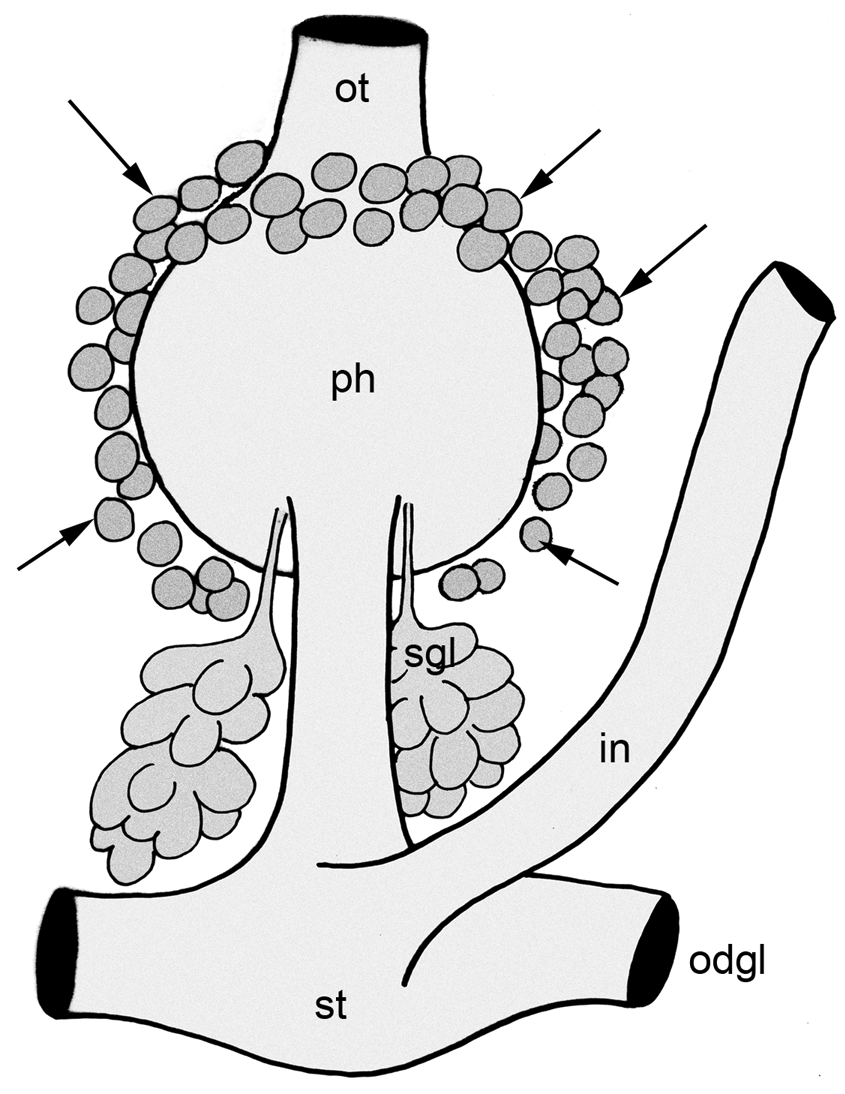
A special glandular layer present, starting at transition of oral tube into pharynx on dorsal and lateral sides ( Fig. 8 View FIGURE 8 A arrows, Fig. 10 View FIGURE 10 ). Further to the posterior, these glands are located only at lateral side, ending on ventral side near transition from pharynx into oesophagus ( Fig. 8 View FIGURE 8 C, Fig. 10 View FIGURE 10 ). This layer consisting of large drop-like glandular cells characterized by bluish granules; no connections to oral tube, pharynx or other parts of digestive system visible.
FIGURE 12. Elysia asbecki sp. nov.: PAM measurements (PSII maximum quantum yield, Φ IIe-max) of one specimen of Elysia asbecki sp. nov. plotted against starvation days. Two to four measurements were taken per day. Mean value of yield values and standard deviation is given (see also Table 7). The line represents the trend line as calculated in Excel.
Molecular investigation. Partial CO1 gene sequences were analysed for two specimens (see Tab. 6) in comparison with closely related Elysia species, according to the results based on the phylogenetic analyses (see Fig. 11 View FIGURE 11 , grey box). Two analysed Elysia asbecki sp. nov. sequences differ only in one nucleotide (sequence divergence 0.15%). The lowest interspecific divergence was found towards an undescribed species from Lizard Island ( Elysia sp. 5) with 10.5% (uncorrected distance), followed by E. macnaei Marcus, 1982 with 11.6% (uncorrected distance). The phylogenetic analysis based on three genes also reflects the close relationship of these three species, but also their distinctiveness.
E. asbecki sp. Elysia sp. 5 Elysia Elysia Elysia pratensis Elysia
nov. (paratype) macnaei subornata tomentosa
Elysia asbecki 0.15 10.52 11.65 13.53 14.66–14.85 15.22–15.41 sp. nov.
Notes on biology and photosynthetic activity. Figure 12 shows measurements of photosynthetic activity plotted against starvation days. Data are shown in Table 7. Maximum quantum yield values (Φ IIe-max) on the day of collecting (but after several hours of starving) start on a higher level of 0.6 and decrease to around 0.5 after nine days of starving. No information on algal food or development is available yet.
Discussion. Taxonomy of Elysia asbecki sp. nov.. The new species described here as Elysia asbecki sp. nov. was already illustrated in two varieties in Wägele et al. 2006b ( Figs. 4 View FIGURE 4 C and D) as Elysia sp.. This material has not been preserved at that time, but was only documented with a digital camera (see also Fig. 7 View FIGURE 7 C this study). Elysia asbecki sp. nov. was also recorded by Gosliner et al. (2008) as Elysia sp. 16. The animal depicted in the photograph clearly shows the black and white ribbon around the upper part of the rhinophores, the red patches along the edges of the parapodia in the middle part, the light stained dot in the neck area and the whitish appearance with the tiny yellow dots.
TABLE 7. PAM measurements (PSII maximum quantum yield, Φ IIe-max) of one specimen of Elysia asbecki sp. nov. during nine starvation days. Two to four measurements were taken per day. Mean value of yield values (see also Händeler et al. 2009) and standard deviation is given. For graphic illustration of the data see Figure 12. The family Plakobranchidae are shell-less sacoglossans with a flattened body, with leaf like lateral expansions (parapodia), which are usually folded up on the dorsal side. The pericardium is located mediodorsally just behind the head. A number of pericardial vessels are found branching from the pericardium along the dorsal surface of the body and parapodia. Marcus d. B.-R. (1980) described the branching patterns of the vessels as species-specific. According to anatomical data, we include the new species to the genus, since the presence of blade-shaped radula teeth with a median denticulate cutting edge is confirmed, and no pharyngeal pouch was found. Molecular data also give evidence for this assignment.
Roughly 80 species of Elysia are described worldwide ( Jensen 2007), more than half of them from the Indopacific region. Additionally, many undescribed species are recorded in this region as well. Around 30 species have been recorded from the Mediterranean Sea and the Atlantic Ocean ( Jensen 2007). Except of E. timida and E. verrucosa Jensen, 1985 , none of these are similar in their external features and coloration to the here newly described Elysia asbecki sp. nov.. Elysia timida differs by the distinct red dots, which are missing in E. asbecki sp. nov., and never shows the distinct red patches at the junction of the parapodia, along the edges of the parapodia and along the end of the foot (pers. observation). The leading tooth is more elongate in E. timida , than in E. asbecki sp. nov.. Elysia verrucosa shows white patches and black spots abundant on the entire body surface ( Jensen 1985). Living animals can be easily distinguished by the green and white irregular patterns in E. verrucosa , as well as the lack of the distinct dark and orange ring on the rhinophores in the latter. The leading radula tooth in E. verrucosa is roundish, where as it is acute in our new species. Elysia asbecki sp. nov. clearly shows short u-shaped rhinophores, similar to those in E. trisinuata Baba, 1949 and E. pusilla ( Bergh, 1872) , but not in E. timida or in E. verrucosa .
The new species will be discussed with the following whitish colored and similar shaped species known from the Pacific Ocean: Elysia mercieri ( Pruvot-Fol, 1930) , E. tomentosa Jensen, 1997 , E. trisinuata and E. pusilla . In terms of color, the new E. asbecki sp. nov. can be distinguished from other species by its whitish appearance due to many tiny white dots, the larger orange spots and tiny dark dots covering mainly the outer parapodia and dorsal body parts. Elysia pusilla differs by the color pattern, the reduced parapodia and the cryptic appearance on their food source Halimeda sp. ( Jensen 1992). E. trisinuata appears similar in shape but the plain green color and the specific three raised folds along the parapodial edge distinguish it from our new species here. Also the radula is very similar, but the teeth in E. trisinuata appear more elongate ( Jensen 1992) than those of the new species described here. E. mercieri is distinguished by the elaborate parapodial margin with structures similar to branched papillae, and the rhinophores show several brownish patches or bands. Elysia tomentosa has distinct papillae, which may even form branched processes. That species never exhibit the typical color patterns of the rhinophores or the red markings of our new species. Furthermore, the radula teeth appear more elongate in E. tomentosa than in E. asbecki sp. nov. ( Jensen 1997).
Comparison of histological results on several sacoglossan species shows that the special glands described here for the first time for Elysia asbecki sp. nov. are typical for members of the Plakobranchidae . A reinvestigation of E. crispata , E. ornata ( Swainson, 1840) , E. timida , E. viridis ( Montagu, 1804) , as well as Plakobranchus ocellatus van Hasselt, 1824 and two Thuridilla species ( T. carlsoni Gosliner, 1995 , and T. hopei (Verany, 1853)) revealed similar glands in the dorso-anterior to lateral parts of the pharynx, additionally to separate salivary glands, which usually lie ventrolaterally and posterior to the pharynx. So far, these glands are absent in non-plakobranchoidean species investigated up to now ( Oxynoe viridi s ( Pease 1861), Alderia modesta , Ercolania annelyleorum sp. nov. and E. kencolesi ) (unpublished results of HW).
Wägele et al. (2006a) mentioned special glandular structures in the parapodia of Elysia ornata . These differ to the cells mentioned here in the parapodia of E. asbecki sp. nov. in so far as there seems to be a dense core composed of several cells in the former. The histologically investigated specimen of E. asbecki sp. nov. was a juvenile and nearly no female structures were formed yet. Comparisons with adult members of other Elysia species clearly show that these special cells in E. asbecki sp. nov. are not part of the albumen gland or even prostate gland, both usually ramifying within the parapodia. The function of these special cell structures in our new species is not known or investigated yet, but needs further analyses.
Molecular characters. Sequence divergence of the partial CO1 gene between the two investigated specimens collected at the same locality and same time is extremely low (0.15%) and lies within the normal range of intraspecific variability as was also observed for two sequences of E. pratensis Ortea and Espinosa, 1996 (0.18%) and of E. tomentosa (0.37%). Values of sequence divergence between Elysia species closely related to E. asbecki sp. nov. are lower, as is observed for the Limapontiidae data set discussed above. They range from about 10% ( E. pratensis / E. subornata Verrill, 1901 ) to a maximum of 15,5% ( E. asbecki sp. nov. / E. tomentosa ). Nevertheless, these species are clearly separated, especially when considering the very low intraspecific variability.
The sequences of the three different molecular markers used in the phylogenetic analysis of Elysia species within Plakobranchidae are already published (see Table 5): 16S rDNA as Elysia spec . (accession number EU 140856 View Materials in Händeler & Wägele 2007), CO1 and 28S rDNA as Elysia spec . 1 (accession numbers GQ996690 View Materials and GQ996629 View Materials respectively in Händeler et al. 2009). The phylogenetic analysis including these three genes unambiguously revealed the assignment of the new species to the genus Elysia , but also clearly showed its distinctiveness to all other included 23 Elysia species ( Händeler et al. 2009, Fig. 11 View FIGURE 11 ).
Photosynthetic activity. There are three different categories of photosynthetic activity in Sacoglossa : no functional retention, short-term retention and long-term retention ( Händeler et al. 2009). Elysia asbecki sp. nov. shows retention with a high starting maximum quantum yield (Φ IIe-max) that decreases only slightly within about ten days. Since retention behaviour can vary on situation and specimen (see Händeler et al. 2009 and references therein) and just one specimen has been investigated concerning photosynthetic activity, it can not be ruled out that Elysia asbecki sp. nov. is a long-term retention form with similar photosynthetic performance as is described for E. timida from the Mediterranean Sea and E. crispata from the Caribbean Sea. This would also render E. asbecki sp. nov. the second long-term retention form in the Pacific, along with Plakobranchus ocellatus . Recently Wägele et al. (2010) were able to reject the hypothesis that lateral gene transfer from the algal nuclear genome to the slugs’ nuclear genome is responsible for maintenance of chloroplasts over weeks to months. Their findings are based on genome expression data of starved Plakobranchus ocellatus and Elysia timida (both long term retention forms with photosynthetic activity over several weeks to months). This is in contrast to findings in E. chlorotica Gould, 1870 based on single PCR gene fragment analysis ( Pierce et al. 2007: Rumpho et al. 2008; Schwartz et al. 2010). Elysia asbecki sp. nov. is more closely related to E. timida than to E. chlorotica ( Händeler et al. 2009) . Hence we consider the properties of the chloroplasts as the main factor for photosynthetic activity. Chloroplasts sequestered by E. asbecki sp. nov. originate from at least three different species of ulvophycean algae (Händeler et al. 2010; as Elysia sp. 1). Unfortunately, these cannot be identified yet to species level, due to lack of reference algal sequences.



No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.