
Branchinecta fueguina
|
publication ID |
https://doi.org/ 10.5281/zenodo.184274 |
|
DOI |
https://doi.org/10.5281/zenodo.6228625 |
|
persistent identifier |
https://treatment.plazi.org/id/C73E3B25-411B-1B3E-6A9E-E1C9AD06EC28 |
|
treatment provided by |
Plazi |
|
scientific name |
Branchinecta fueguina |
| status |
|
Branchinecta fueguina View in CoL nov. sp.
Type material. Deposited in the Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”. Holotype, temporary pond in Tierra del Fuego province ( Argentina), 53°47'12'' S, 67°46'55'' W, male number: MACN-In 37498; date: January 23, 2001; collector: M. C. Marinone; Allotype, same collecting data as holotype, female (ovigerous), number: MACN-In 37499.
Type locality. temporary pond by Route C, in front of Laguna de los Cisnes, behind Río Grande Airport (53°47'12'' S, 67°46'55'' W), Tierra del Fuego, Antártida e Islas del Atlántico Sur Province, Argentina.
Etymology. The specific epithet is a toponym of Tierra del Fuego, from which the specimens came.
Description. Male: Mean total length from front to end of the abdomen with uropods (cercopods) without setae: 16.9 mm (16.1–18.1 mm). Mean uropods (cercopods) without setae length: 1.3 mm (1.1–1.4 mm). Average ratio of “head + thorax” to “abdomen + uropods without setae”: 1.02.
Pedunculate eye diameter: 0.6–0.7 mm. Antenna 1 as usual, filiform and slender, 2.3–2.8 mm, long. Antenna 1 length is 4 times the diameter of compound eye.
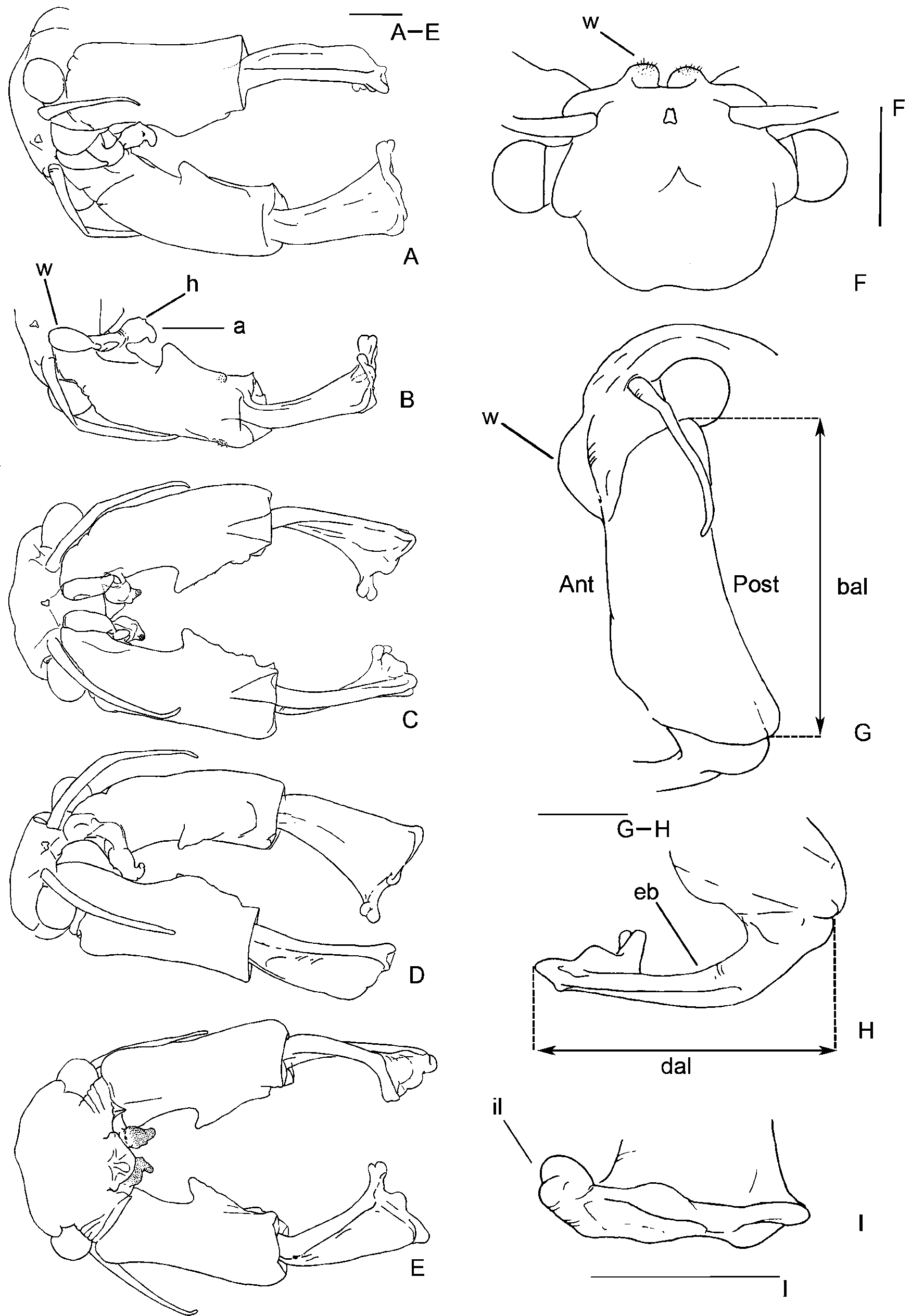
Antenna 2 basal article ( Figs 1 View FIGURE 1 A–E), 2.7–3.1 mm long ( Fig. 1 View FIGURE 1 G), bearing a proximal anteriomedial rough welt provided with sensory setae, projecting beyond basal article anterior surface and visible in dorsal view ( Fig.1 View FIGURE 1 F). Welt extending posteriorly and terminating as a conspicuous apophysis ( Figs 1 View FIGURE 1 A–E). Apophysis hanging medially from the posteriomedial side of article, proximally swollen until it reaches a heel shaped area ( Fig. 1 View FIGURE 1 B), arcing anteriorly, apex rounded. Apophysis and apex posterior surface with cuticular ornamentation and with some scattered sensory setae ( Fig. 1 View FIGURE 1 E). Basal article proximomedial half concave and smooth ( Figs 1 View FIGURE 1 C, 1E). Basal article distal half with a longitudinal crest produced proximally in a strong blunt medioposteriorly directed point with some scattered small sensory setae. Crest lengthened following the major axis of the article but not reaching its distal end ( Figs1 View FIGURE 1 A–E). Crest margin uneven ( Figs 1 View FIGURE 1 C, 1E), with sparse hyaline sensory structures.
Antenna 2 distal article ( Figs 1 View FIGURE 1 A–E, 1H), 2.2–3.1 mm long, is strongly rotated anteriorly and articulates almost perpendicularly to basal article. Therefore, in head anterior view, both the basal article anterior surface and the distal article lateral and anteriorly rotated margin can be seen at the same time ( Fig. 1 View FIGURE 1 C). This margin curves very slightly to the article apex. Anteriomedially rotated article surface is flattened, slightly convex, adopting a strikingly triangular shape ( Figs 1 View FIGURE 1 A–E), widening strongly from its proximal articulation (“vertex” of the triangle) to the apex (“base” of the triangle). Distal portion of article bends anteriorly forming an explanate complicated three-lobed apex ( Figs 1 View FIGURE 1 A-E, 1I). The inner (medial) lobe is prominent, and subdivided distally into two rounded lobes ( Fig. 1 View FIGURE 1 I). Mouth parts typical of the genus. Maxillae 1 with 21-22 long setae plus a short strong spine on limb ventral tip, base as wide as twice long seta width. Maxillae 2 with 5 plumose distal setae plus 1–2 anteriorly directed densely plumose setae, with setules all around.
Thoracic appendages typical of the genus.
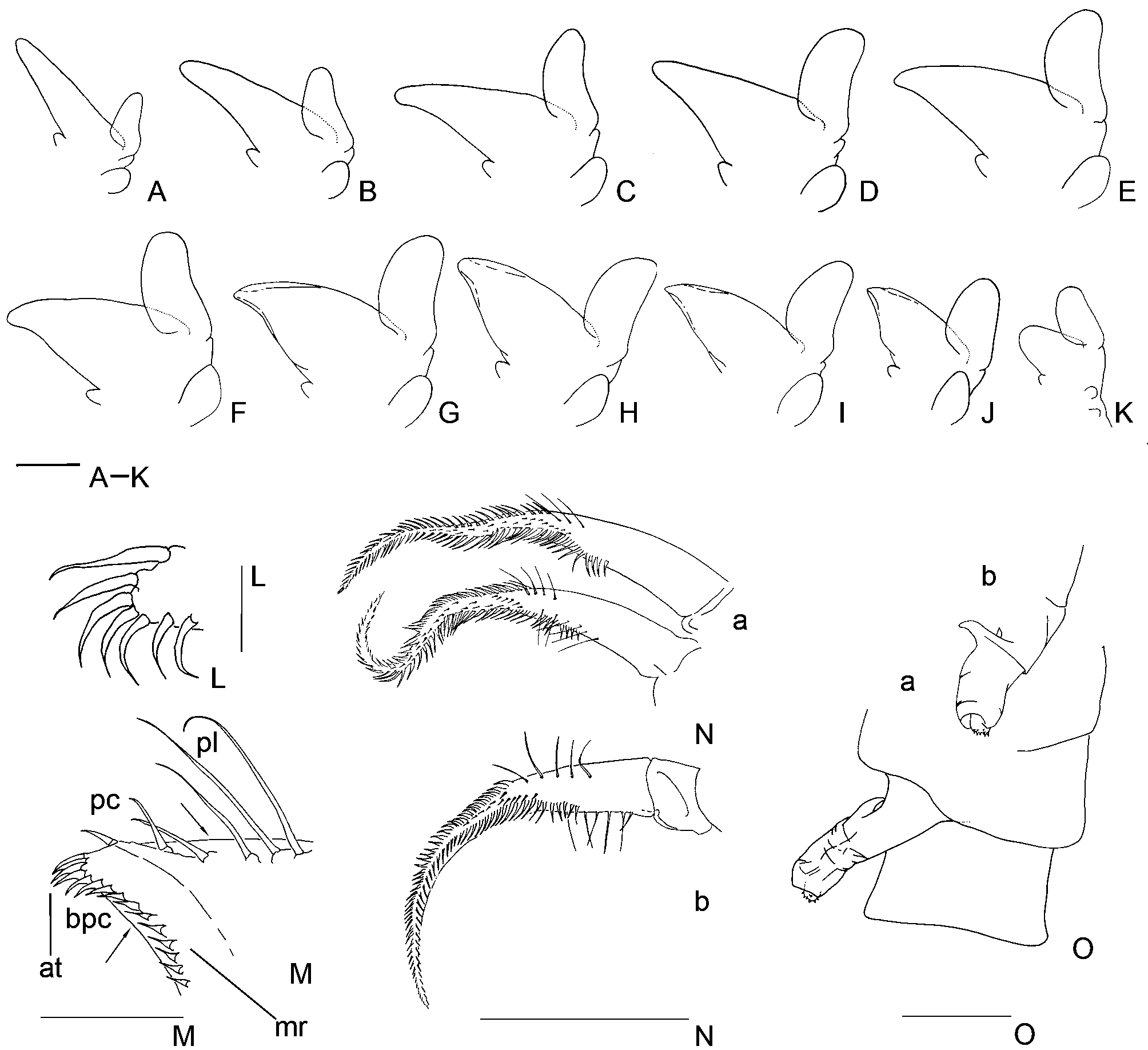
First through 10th endopodites triangular elongate ( Figs 2 View FIGURE 2 A–J), very narrow in first two (endopodite length/ endopodite width ratio 3.7 and 3.2 respectively) gradually widening more than length until 7th limb (endopodite length/endopodite width ratio varies between 3.2 and 1.9); 8th to 10th thoracopod endopodite reduces in size yet maintaining the length and width proportions. First through 10th endopodites with medial cuticular reinforcement, as in other Branchinecta ( Cohen 1995) . 11th endopodite suboval ( Fig. 2 View FIGURE 2 K), without medial reinforcement.
Endopodite and exopodite length ratio (without setae) is approximately 2.5 in the first thoracopod, varies from 2.0 to 1.7 in the 2nd to the 7th thoracopod, and diminishes from 1.6 to 1.4 in the 9th and 10th thoracopod. In the 11th thoracopod the exopodite is slightly longer than the endopodite.
Thoracopod 1 endopodite lower margin proximal half with plumose setae and middle of the distal half with only one half-plumose, half-pectinate seta; more distally, close to the apex of endopodite are two shorter robust pectinate setae. Thoracopods 2 through 10 endopodite lower margin with proximally plumose setae advancing more distally as the appendage series progresses; distally in the margin, with 2–3 widely spaced pectinate setae, decreasing in size and increasing in robustness to apex, and restricted to a progressively shorter margin portion in successive pairs. Thoracopods 1 and 2 endopodite apex with two or three bipectinate uniformly curved or sinuous setae bearing short setules ( Figs 2 View FIGURE 2 L, 2N a, b); these setae are similar shaped though slightly longer than the setae of the endopodite medial margin, which are more spaced than the apical ones. Thoracopods 3 through 10 endopodites with an apical tuft of close set robust bipectinate curved setae similar to those present in thoracopods 1 and 2 apex. Also, the setae along the endopodite medial margin are similar in shape and spacing to the setae in the thoracopods 1 and 2 but smaller and more gracile, almost half the length of the setae of the apical tuft, seeming denticles with short lateral setules ( Fig. 2 View FIGURE 2 M); as consequence, the tuft of scratching setae of the apex seems more conspicuous. In thoracopods 1 through 10, all over the medial margin, from its distal to its proximal part, the curved setae gradually become straight and at the same time sinuous in their tip, whereas in the proximal part of the margin the setae are curved in the opposite direction of the distal setae; the setae are almost uniform in length along the margin. From thoracopod 7 trough 10, a group of setae in the lower and the medial margins are sub-marginally inserted ( Figs 2 View FIGURE 2 G–J; arrows in Fig. 2 View FIGURE 2 M); maybe, this arrangement is related with the folding of the endopodites and could allow the setae to change their attack angle.
In the 11th pair of thoracic appendages, the lower margin of endopodite is only provided with plumose setae. Apex of the lobe widely rounded; the setae bordering it are thin, bipectinate, and slightly curved. Towards the medial margin, in distal to proximal direction, the setae decrease in size; they are bipectinate and gradually become straight and then curved in inverted-direction.
The 1st and 2nd endites in all the thoracic appendages, with the typical anostracan number and arrangement of anterior setae ( Linder 1941). In the 1st pair of thoracic appendages the number of anterior setae on the remaining endites differs from the values stated by Linder (1941) for the genus: 4 to 5 anterior setae were found in the 3rd endite, 5 to 7 in the 4th endite and 5 to 7 in the 5th endite. On the other hand, in the 2nd to 11th pairs of thoracic appendages, the number of anterior setae agrees with the values stated by Linder (1941): 2 for the 3rd endite, 2 for the 4th and 4 to 6 for the 5th. In addition, thoracopods 1st to 10th show elliptical branchial sacs with smooth margins, and branchial laminae with serrated margins without a conspicuous marginal deep notch (although in some appendages small discontinuities were observed). In the 11th pair, both branchial sac and lamina are markedly reduced, rounded and triangular shaped respectively ( Fig. 2 View FIGURE 2 K).
Genital segments not much prominent in lateral view. Gonopods (according to Rogers et al. 2007), typical of the genus ( Fig. 2 View FIGURE 2 O a, b). Non-eversible part with a sclerotized, curved and sharp spur-like process. Eversible part, with the two dentate warts typical of the genus; in one male, in the middle of this part, one gonopod of the pair also showed a couple of small triangular and flattened evaginations ending in a hyaline sinuous tip; these structures have been interpreted like anomalies. Testes extending 2 or 3 segments behind the genital segments.
Joint between the two last abdominal segments, not well defined. Uropods not convergent, uniformly provided with plumose setae all over the margin.
Female: Mean total length from front to end of the abdomen with uropods (cercopods) without setae: 15.8 mm (14.3–17.3mm). Mean uropods (cercopods) without setae length: 0.9 mm (0.8–1mm). Although females are smaller than males, the proportions among tagmata are maintained. Average ratio of “head + thorax” to “abdomen + uropods without setae”: 1.04.
Pedunculate eye 0.4 mm diameter, smaller than in the male. Antenna 1 as usual, filiform and slender, shorter than in the male, 0.9–1.1 mm long. Therefore, unlike the male, antenna 1 length is only 2.4 times the diameter of compound eye.
Antenna 2 ( Fig. 3 View FIGURE 3 A) sub-conical, with the usual verrucose sensory areas with hyaline hairs, ending in a spine which fits 5 to 7 times in the whole appendage length (antenna 2 total length: 1.4–1.7 mm; spine length: 0.2–0.3 mm.
Mouth parts as in the male. Maxillae 1 with 21 long setae plus a short strong spine on limb ventral tip, base as wide as twice long seta width. Maxillae 2 with 6 plumose distal setae plus 1–2 anteriorly directed densely plumose setae, with setules all around.
Thoracic appendages uniform in type and arrangement of setae bordering the lobes. Endopodite not lengthened as in the male, sub-rectangular in thoracopods 1st through 5th, sub-oval in pairs 6th to 11th; without cuticular reinforcement along the medial margin. First through 10th thoracopods endopodite lower margin with plumose setae, and distally 2 to 4 long pectinate ones. Distal medial endopodite margin with curved pectinate setae smaller than those of distal lower margin, decreasing slowly in size and becoming straight bipectinate towards proximal medial margin. Thoracopod 11th lower margin, with plumose setae all along. Rounded apex and medial margin with straight and bipectinate setae, gradually decreasing in size proximally.
The first and 2nd endites in all the thoracic appendages with the typical anostracan number and arrangement of anterior setae ( Linder 1941). In the 1st pair of thoracic appendages the number of anterior setae on the remaining endites differs from the values stated by Linder (1941) for the genus: 4 to 6 anterior setae were found in the 3rd endite, 5 to 7 in the 4th endite and 5 to 6 in the 5th endite. On the other hand, in the 2nd to 11th pairs of thoracic appendages, the number of anterior setae agrees with the values stated by Linder (1941): 2 for the 3rd endite, 2 for the 4th and 4 to 8 for the 5th. Branchial sacs elliptical in pairs 1st to 10th and somewhat pointed from pairs 5 to 10, with smooth margins; branchial laminae as in the male. In the 11th pair, both branchial sac and lamina are markedly reduced, rounded somewhat pointed and triangular shaped respectively.
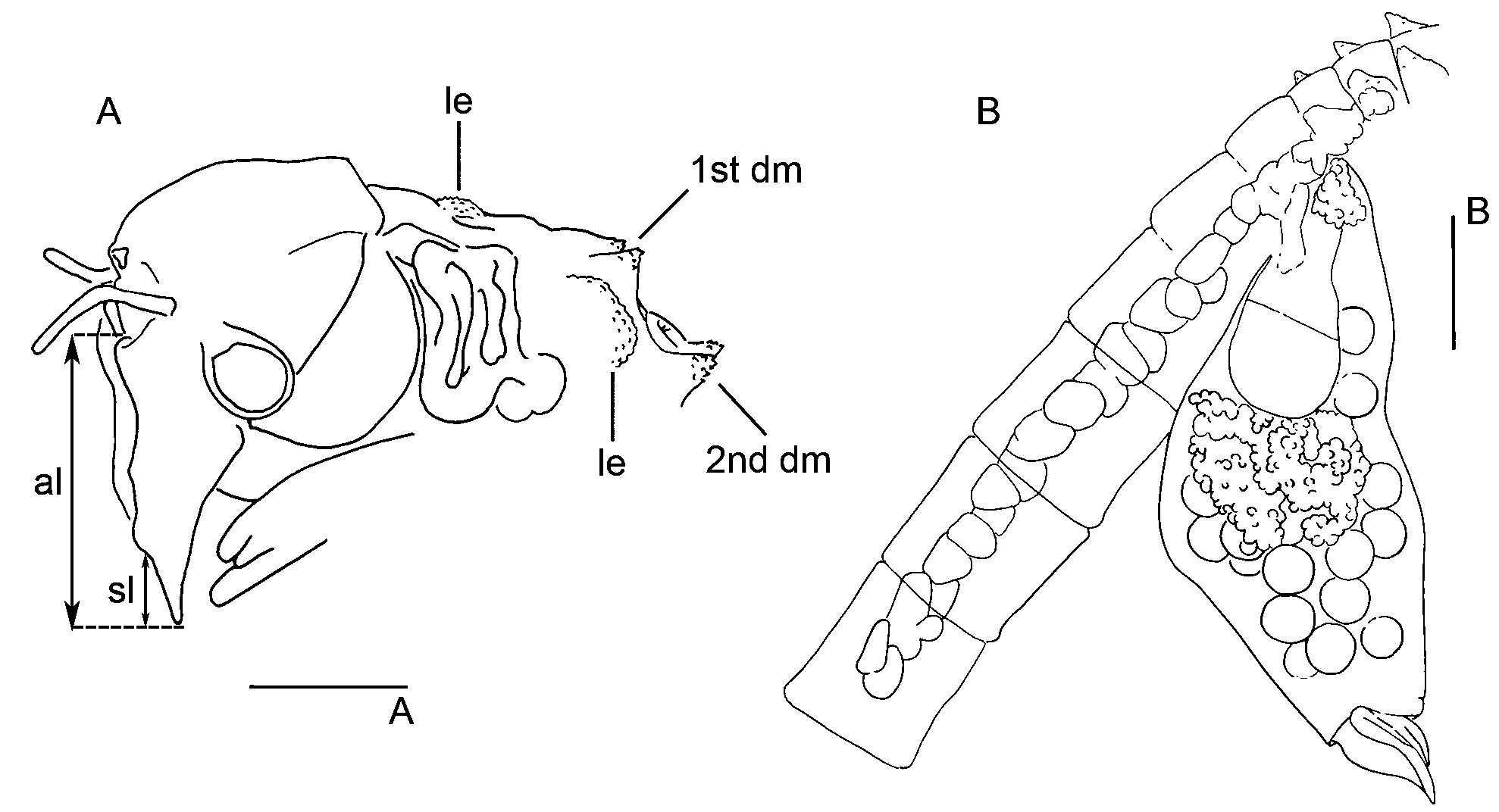
First thoracic segment bearing a pair of lateral roughed auriculate evaginations just posterior to maxillary gland, and a pair of small dorsal and conical spiny mounds with the apex posteriorly directed ( Fig. 3 View FIGURE 3 A), which initiates a row of dorsal similarly shaped mounds, that extends up to 11th thoracic segment. The 2nd pair of mounds is the most prominent of the series; the remaining pairs slightly decreasing in size posteriorly. Both mounds of each pair are very close in the dorsal middline.
Brood pouch fusiform ( Fig. 3 View FIGURE 3 B), dorsally more convex than ventrally, forming an angle of 30° with the longitudinal axis of the body and extending to the 3rd–5th post-genital segments. The lips at the apex are the dorsal almost twice the length of the ventral one. Oviducts wide, fitting up to 3 oocytes in its width. Ovaries extending at most to the 8th thoracic segment forwards and to the 4th post-genital segment backwards. Cyst diameter 299–403 µm.
Joint between the last two abdominal segments not well defined. Uropods as in the male.
Discussion. Our collection of Branchinecta fueguina nov. sp., and B. granulosa , represents the southernmost branchinectid records in the American continent (53°47'12'' S, 67°46'55'' W). However, it does not constitute the unique presence of anostracans in these high American latitudes since Artemia persimilis Piccinelli & Prosdocimi (Artemiidae) is reported from laguna Amarga (50°29’S; 72°45' W) in Torres del Paine (Región XII) ( Gajardo et al. 1999; De los Ríos & Zúñiga 2000) and from laguna de los Cisnes (53º15´S, 70º 22’ W) in the Chilean Tierra del Fuego ( Crespo & Baessolo 2002; Amat et al. 2004), the last site as the most meridional known location of Artemia . Further south, outside the continent, B. gaini Daday, 1910 inhabits Antarctica and the South Atlantic Ocean islands.
The new species is similar to other Branchinecta having robust, short and coarse male antenna 2. It shares this trait with B. granulosa , B. gaini , B. valchetana Cohen, 1981 , B. prima Cohen, 1983 , and B. vuriloche Cohen, 1985 from South America; and with B. packardi Pearse, 1912 , B. belki Maeda-Martínez, Obregón Barboza and Dumont, 1992 , B. constricta Rogers, 2006 , B. hiberna Rogers and Fugate, 2001 , B. cornigera Lynch, 1958 , and B. mexicana Maeda-Martínez, Obregón Barboza and Dumont, 1993 from North America. However, it differs from these species in the shape and armature of the proximal and/or the distal article of the antenna 2.
The male antenna 2 basal article of B. granulosa , B. gaini , B. vuriloche , B. constricta , B. cornigera , B. prima , B. valchetana , and B. hiberna ( Cohen 1981, 1983, 1985, 1992; Daday de Dées 1902, 1910; Lynch 1958; Rogers 2006; Rogers & Fugate 2001) have a convex proximomedial pulvillus, with spinules in the five former species, with sparse sensilla in B. prima ( Cohen 1983) , with a single broad and rough spine in B. valchetana ( Cohen 1981) ; or like a spiny ridge in B. hiberna ( Rogers & Fugate 2001) . In contrast B. fueguina nov. sp., has a convex welt and conspicuous apophysis, similar to that observed in B. belki , B. mexicana and B. packardi ( Lynch 1964; Maeda-Martínez, et al. 1992, 1993, Pearse 1912).
However, B. fueguina nov. sp. differs from these species by the presence of a longitudinal medial crest, produced medially into a strong blunt point, and gradually declining distally; the crest and point are provided with small sparse sensory setae. B. belki has a medial distinctive prominent outgrowth with 2-3 spiny protuberances ( Maeda-Martínez et al. 1992); B. packardi and B. mexicana present a peg like squamate appendix, more extended distally and with transverse clefts in B. mexicana ( Maeda-Martínez et al. 1993).
The morphology of the male antenna 2 distal article in B. fueguina nov. sp. is the main morphological trait that distinguishes the species from other branchinectids, because of its peculiar articulation, its remarkable triangular and strikingly flattened shape and its tip. Distally, the broadened article bends anteriorly forming an extended and complicate three-lobed tip (base of triangle), with the inner (medial) lobe divided into two rounded lobes. On the other hand, in B. belki the apex is bifurcated, with a dorsal acute and a ventral round projections ( Maeda-Martínez et al. 1992); in B packardi the tip of the article tapers a little, its anterior half turning inward in a medially projecting spur and its posterior half, short, not turning medially ( Lynch 1964); in B. mexicana , the apex of the article is not notably inflexed ( Maeda-Martínez et al. 1993).
Like in the majority of species of the genus, B. fueguina nov. sp. shows a marked sexual dimorphism particularly evident in the thoracic appendage setose armature (arrangement and shape of setae) and in the endopodite shape. In the male, the distally extended triangular endopodites of the 10 former thoracopods of the series bear a cuticular reinforcement along the medial margin lacking in the shorter sub-rectangular to suboval endopodites of the female. This structure is also observed in the male endopodites of other Branchinecta , bearing distally extended endopodites ( Cohen 1993, 1995), such as B. iheringii Lilljeborg, 1889 , B. palustris Birabén, 1946 , B. rocaensis Cohen, 1982 , and B. valchetana .
The articulation between the two last abdominal segments, in both sexes, is not well defined in B. fueguina as it occurs in other branchinectids like B. valchetana and B. prima ( Cohen 1981, 1983).
The dorsal row of conical spiny mounds along the female 1st to 11th thoracic segments, may be of relative taxonomical value. It is worth noting that even though several authors have stressed the usefulness of the trait as a specific character in the genus ( Linder 1941; Lynch 1958, 1972; Cohen 1981, 1985, 1987, 1992, 1993; Rogers 2002), they have also pointed out the great variability of the female trunk armature among populations and according to the size of specimens within each population.
No known copyright restrictions apply. See Agosti, D., Egloff, W., 2009. Taxonomic information exchange and copyright: the Plazi approach. BMC Research Notes 2009, 2:53 for further explanation.